| FEIS Home Page |
 |
|
Gray alder thicket. Photo by Robert Vidéki, Doronicum Kft., Bugwood.org. |
Alnus incana (L.) Moench subsp. rugosa (Du Roi) R.T. Clausen, specked alder [120,137,192,205,237,255]
Alnus incana (L.) Moench subsp. tenuifolia (Nutt.) Breitung, thinleaf alder [120,167,179,192,355]
European gray alder, Alnus incana (L.) Moench subsp. incana, is native to western Europe [120,127]. It has been introduced in the northeastern United States [231]. Since variation in the subspecies is continuous rather than discrete, gray alder subspecies are sometimes difficult to tell apart ([127], review by [143]). Speckled alder and thinleaf alder intergrade where their ranges overlap, mostly in Saskatchewan and Manitoba [106,120,127,231].
In this review, gray alder refers to information that is general to the species. Speckled alder refers to A. i. subsp. rugosa, and thinleaf alder refers to A. i. subsp. tenuifolia.
Hybrids: Gray alder hybridizes with European alder (A. glutinosa), an introduced species [133]. Speckled alder hybridizes and intergrades with hazel alder (A. serrulata) [120,231,295]. Speckled alder × hazel alder hybrid swarms may occur in Massachusetts and elsewhere [127]. Thinleaf alder may hybridize with red alder (A. rubra) in Idaho [202].
SYNONYMS:For thinleaf alder (Alnus incana subsp. tenuifolia):
Alnus incana (L.) Moench var. occidentalis (Dippel) C.L. Hitchc. [208,304]
Alnus incana (L.) Moench var. virescens S. Watson
Alnus × purpusii Callier
Alnus tenuifolia Nutt. [155,208,231,242]
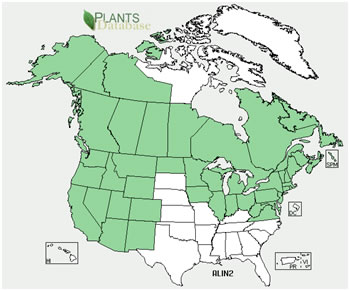
| Distributions of gray alder and its subspecies. Maps courtesy of USDA, NRCS. 2011. The PLANTS Database. National Plant Data Team, Greensboro, NC. (14 June 2011). | |
|
Gray alder occurs in North America and Europe [120,192,296]. It occurs in the northeastern and central regions of Europe, extending locally into the southern mountains [296]. States and provinces (as of 2011 [335]): |
 |
Speckled alder,
the eastern subspecies, occurs at least as far west as Saskatchewan [120,192]. Little [230] places its distribution as far west and north as northwestern Yukon,
central Northwest Territories, and central Nunavut. Its distribution is mostly continuous, with scattered populations in Michigan, Ohio, and West Virginia [230].
States and provinces (as of 2011 [335]): |
 |
Thinleaf alder, the western subspecies, has a mostly continuous distribution. Populations are scattered and sometimes isolated in the southern Sierra
Nevada [179,192,229,229]. It has been reported in Baja California, although it may no longer occur there [358].
States and provinces (as of 2011 [335]): |
The alder, whose fat shadow nourisheth.
Each plant set there to him long flourisheth.
—William Brown, Brittania's Pastorials. 1613.
Speckled alder commonly occurs or codominates with willows (Salix spp.), a mixture that may be controlled by length of flooding season. Willows may tolerate longer flooding seasons than speckled alder. A laboratory study showed speckled alder was less tolerant to continuous flooding than willow species. Speckled alders suffered 33% average mortality when water levels were kept above the root crown for 2 years, while speckled alders experienced no mortality when the water level was maintained below the root crown for the same length of time [196].
Soils: Speckled alder tolerates a wide variety of soil textures and pH but is most common on nutrient-poor soils. It grows in sandy loam [49,160], chalky, rocky till, and mucky soils [160]. In Ontario, speckled alder is an indicator of fine loamy-clayey soils with thick black humus; it is also an indicator of poor to rich fens with moderately decomposed organic soils [188]. In swamps of northern Michigan, speckled alder communities tended to occur on relatively well-drained, sandy loams, while black ash/speckled alder communities occurred mostly on poorly drained, silty clay loams [275]. In the Catskill and Adirondack mountains, speckled alder grows in limestone-, gneiss- and anorthosite-derived soils [205]. In black ash-paper birch communities of the Great Lakes states, speckled alder was noted on minerotrophic peat substrates [31], and it grew in coarse woody peats in northern white-cedar (Thuja occidentalis) swamps of northern Wisconsin [67]. It occurred on glacial till in central New York; soils were neutral to "somewhat alkaline" [176]. It is reported on soils with pH from 3.4 to 7.7 in Ohio (review by [160]). In Wisconsin, speckled alder thickets occurred on peats of pH 5.5 [85]. In jack pine (Pinus banksiana) stands in Minnesota, speckled alder was most common on medium-quality silvicultural sites (18% frequency). It also occurred on good-quality sites (7% frequency) but not on poor-quality sites (Hansen 1946 cited in [39]). In an assessment of Michigan shrub habitats, speckled alder occurred on the moistest, coolest, and most nutrient-poor sites when compared to other shrubs [36]. However, speckled alder also occurs on nutrient-rich sites [204]. On the Saskatchewan River Delta, speckled alder was positively associated with relatively nutrient-rich, shallow peat fens and levees. Soil water was alkaline and high in calcium and magnesium [97].
Elevation and topography: Speckled alder ranges from near sea level on the North Atlantic coast to about 2,710 feet (825 m) in the Appalachians [127]. It grows on low-elevation alluvial soils in Nova Scotia [295] but at "moderately high elevations" elsewhere in Canada [304]. It is restricted to high elevations in the southernmost portion of its range [160]. In a tamarack bog in Itasca State Park, Minnesota, speckled alder was more abundant on hummocks (35% cover and 10% frequency) than in depressions (2% cover and <1% frequency) [142].
Plant communities: Speckled alder occurs in forest, shrub, and herbaceous communities. It is an important component of northern spruce-fir (Picea-Abies spp.) [22,247], red-white-jack pine (Pinus strobus-P. resinosa-P. banksiana) [69], Atlantic white-cedar (Chamaecyparis thyoides) [106], and mixed-hardwood [341] forests, especially at forest and bog ecotones [247]. It also occurs in elm-ash-cottonwood (Ulmus-Fraxinus-Populus spp.) galleries and forests [108]. In Newfoundland, it occurs in rich fens dominated by pale sedge (Carex livida) [289]. Species diversity is generally high in wetland communities where speckled alder is an important to dominant component of the vegetation (for example, [33,114,356]).
Regional descriptions of plant communities with speckled alder and site characteristics of speckled alder habitats follow.
Great Lakes: Speckled alder is common in coniferous, mixed conifer-hardwood, shrubland, and graminoid wetland communities in the Great Lakes region. It is common, for example, in the understories of red pine forests and plantations. American hazelnut (Corylus americana) and beaked hazelnut (C. cornuta var. cornuta) usually dominate or codominate the understories [46]. In Voyageurs National Park, Minnesota, black spruce/speckled alder communities occurred on nutrient-poor, wet sites [207]. On the Lake Agassiz Peatlands Natural Area of Minnesota, speckled alder was abundant (covered 0.25% to 0.5% of an area) in rich swamp forests dominated by northern white-cedar, black ash (Fraxinus nigra), tamarack (Larix laricina), or white spruce (Picea glauca). It was not abundant in poor swamp forests, bogs, fens, or heaths [163]. In northern Wisconsin, speckled alder was important to dominant in northern white-cedar swamps [67]. It was also important to dominant in eastern redcedar (Juniperus virginiana) bogs of Ohio [125]. On the shores of Lake Superior, a speckled alder/red raspberry /fowl mannagrass-reed canarygrass (Rubus idaeus/Phalaris arundinacea-Glyceria striata) thicket community occurred on secondary dunes, lees of high dunes, and sand flats. This community type had the highest species richness of 4 types identified [33].
Speckled alder is occasional to invasive on open fens and bogs of the Great Lakes states. In northeastern Ohio, it is characteristic in sedge (Carex) fens. Bottlebrush sedge (C. hystericina), hairy sedge (C. lacustris), and tussock sedge (C. stricta) are typical dominants [301]. Speckled alder often dominates marginal fen or marginal bog zones, which separate the mires from the upland communities surrounding them [73].
Northeast: Speckled alder grows in coniferous forests, mixed and hardwood forests, shrublands, and open wetlands in the Northeast.
Maine: On Peak's Island, Portland, speckled alder was characteristic in the tree strata of an upland red maple (Acer rubrum) forest, parts of which had burned 3 times in 24 years, and of an unburned balsam fir (Abies balsamea) forest. At the time of the survey, speckled alder was the 2nd most common taxon on 7-year-old burned sites in the red maple forest (97% relative density and 13% frequency on 10 m² plots) and the 4th most common in the balsam fir forest (56% relative density and 16% frequency). Speckled alder clumps were also frequent in a nearby red maple swamp, where speckled alder dominated poorly drained areas of the swamp [79].
New England: In pitch pine (Pinus rigida)-hardwood swamp mosaics of New Jersey, speckled alder was a minor to codominant component of red maple-black tupelo-sweetbay/coastal sweetpepperbush (Nyssa sylvatica-Magnolia virginiana/Clethra alnifolia) swamps. It was most important on continuously flooded sites, declining on sporadically flooded or dry sites [112].
Southeast: Speckled alder occurs in mixed-hardwood, pine (Pinus spp.), and shrubland communities of the Southeast. In a 1930 study, it was a component of a climax American chestnut-yellow-poplar-eastern hemlock (Castanea dentata-Liriodendron tulipifera-Tsuga canadensis) cove forest in the Black Mountains of North Carolina [91]. Yellow-poplar became the primary dominant in many cove forests after chestnut blight decimated American chestnut populations [341]. In Georgia, speckled alder occurs in pond pine (Pinus serotina)-shrubland and bay (Magnolia and Persea spp.)-shrubland bogs. Shrub diversity is generally high [356].
The following vegetation classifications describe plant communities, and some of their distinguishing site characteristics, where speckled alder is dominant. In state and province lists, plant communities are arranged geographically from north to south and west to east.
| Great Lakes | Northeast | Southern Appalachians | Southeast |
| Canada |
Thinleaf alder
Site characteristics:
Thinleaf alder is most common on wet to moist sites (review by [143]). It is a frequent component of streamside vegetation throughout mountainous regions of western North America [120]. It is considered an indicator of riparian or subirrigated sites on the Shoshone National Forest, Wyoming [172]; of moist, well-drained sites—especially streambanks and springs at low elevations—in central Oregon [203]; and of moist to wet soils in subboreal spruce (Abies spp.) and pine (Pinus spp.)-spruce ecosystems of British Columbia [23]. Riparian sites with thinleaf alder may experience frequent flooding and/or scouring (for example, [104,114]).
Soils: Thinleaf alder grows on a variety of soil textures and nutrient levels. It grows in coarse-textured soils but is well-adapted to cold, "heavy" soils (review by [143]). It grows well in sand [199,339], and it is common on sandbars [339]. In the Trout Creek Mountains, thinleaf alder grows in sand- and siltbanks about 2.6 feet (0.8 m) above stream channels [114]. Soils supporting thinleaf alder are often rocky in mountainous areas [318]. In Montana, thinleaf alder communities typically develop on cobble and gravel, accumulating organic debris over time [149]. Nutrient levels of soils supporting thinleaf alder vary from poor to rich, although they are typically poor (review by [184]). Thinleaf alder typically establishes on poor, skeletal soils in primary succession [149], but it usually enriches in soils in which it grows [149]. In the understories of coniferous forests in Blue Mountains, thinleaf alder is an indicator of productive sites [144].
Elevation and topography: In the conterminous United States, thinleaf alder is primarily restricted to mid- to high-elevation mountains, mountain valleys, and mesic canyons [155,208,231,241], although it grows on low-elevation sites in Alaska and Canada [231]. In the United States, thinleaf alder ranges in elevation from near sea level in Alaska to over 10,000 feet (3,000 m) in Colorado and Arizona [127]. Thinleaf alder populations apparently do not have exacting elevational requirements. Thinleaf alder cuttings have been successfully transplanted onto sites that vary greatly in elevational range [256].
| Elevational range of thinleaf alder by state | |
| State | Range (feet) |
| Arizona | 5,000-9,000 [195,228] |
| California | 3,900-7,900 [167] |
| Colorado | 5,000-10,000 [155] |
| New Mexico | 6,000-9,500 [96,228,241]; 1,745-2,438 in Jemez Mountains [6] |
| Nevada | 5,000-8,000 [193] |
| Texas | 5,000-8,000 feet [348] |
| Utah | 4,100-9,005 [355] |
Thinleaf alder presence is likely more associated with moist to mesic conditions than with aspect alone. In the Wasatch Mountains of Utah, thinleaf alder was common in the bottom of Cold Canyon [272], but it was not noted on higher, north- or south-facing slopes, which may have drier soils [272]. In Lassen Volcanic National Park, California, the location of willow-thinleaf alder communities was not significantly associated with aspect, but it was positively associated with relatively moist conditions (P=0.01). Willow-thinleaf alder communities were most common on steep slopes, depressions, and valley bottoms (P≤0.002). The authors attributed their presence on steep slopes to the prevalence of intermittent streams in avalanche chutes and their presence in low areas to the prevalence of concave drainage channels [288].
Plant communities: |
Thinleaf alder prefers mesic to moist plant communities. These include the moist slopes of coniferous forests, riparian areas, wet meadows and grasslands, and fen and bog margins [127]. Site moisture and elevation are critical in determining relative dominance of thinleaf alder; the surrounding vegetation usually tolerates drier conditions. On the shore of Slave Lake in Alberta, thinleaf alder codominates the understory of white spruce-paper birch communities on gravel or sand beaches that are 300 feet (90 m) or less from the lake [227]. In the Trout Creek Mountains of Nevada, thinleaf alder communities are concentrated in low-elevation (4,920-5,300 feet (1,500-1,615 m)), narrow riparian corridors. Mountain big sagebrush (Artemisia tridentata subsp. vaseyana) typically dominates vegetation beyond the corridors, and plant species diversity is lower outside than within the riparian corridors [114]. Willows (Salix), red-osier dogwood, common snowberry (Symphoricarpos albus), and horsetails (Equisetum) codominate with thinleaf alder in many plant communities throughout much of thinleaf alder's distribution.
Regional descriptions of plant communities with thinleaf alder and site characteristics of thinleaf alder habitats follow.
Alaska: Thinleaf alder is reported on glacial shrublands (see Successional Status), riparian and wet-upland shrublands, and spruce-birch (Picea-Betula spp.) swamps. Thinleaf alder-willow and resin birch-thinleaf alder-willow cover types are common near timberline in interior Alaska. These communities form thickets that may be "extremely dense" or open and interspersed with reindeer lichens (Cladonia) and heath (Ericaceae) [357]. On the Tanana River of central Alaska, 5-year-old thinleaf alder communities on sandbars were codominated by feltleaf willow (Salix alaxensis), sandbar willow (S. interior), and barrenground willow (S. brachycarpa subsp. niphoclada). Balsam poplar, paper birch (B. papyrifera), and white spruce grew in the understory; meadow horsetail (E. pratense) and variegated scouringrush (E. variegatum) were dominant herbs [339]. See Successional Status for more information on this study.
Pacific Northwest: Thinleaf alder is a component of and sometimes forms glades within coniferous forests of the Pacific Northwest; it is also important to dominant in riparian corridors. Thinleaf alder is prevalent in ponderosa pine (Pinus ponderosa), Douglas-fir (Pseudotsuga menziesii), lodgepole pine (Pinus contorta), and fir-spruce (Abies-Picea spp.) communities [184]. Thinleaf alder glade openings generally occur on wet to mesic sites where thinleaf alder outcompetes conifers for soil moisture [92].
California: Thinleaf alder grows in coniferous, western hardwood, riparian, and other wetland communities in California. The photo above shows a typical Californian thinleaf alder habitat. Thinleaf alder is frequently found on the edges of ponds, bogs, and fens [242]. In the Klamath Mountains and Sierra Nevada, it grows on the margins of California pitcher plant (Darlingtonia californica) bogs [221]. Prior to extensive water diversions, thinleaf alder was likely important in quaking aspen-black cottonwood (Populus balsamifera subsp. trichocarpa) riparian corridors leading to Mono Lake, which is situated in the arid Mono Basin [320].
Northern and Central Rockies: Thinleaf alder is reported in coniferous forests, riparian galleries and shrublands, and wetland edges. In western Montana, thinleaf alder is frequent in Douglas-fir, western hemlock-western redcedar (Tsuga heterophylla-Thuja plicata), and grand fir (Abies grandis) forests. Surveys in the Blackfoot and Flathead valleys indicate that most thinleaf alders in these forest types grow as shrubs [121]. In southwestern Colorado, thinleaf alder grows in mixed-conifer riparian woodlands dominated by white fir (A. concolor), southwestern white pine (Pinus strobiformis), interior ponderosa pine (P. ponderosa var. scopulorum), and blue spruce (Picea pungens) [14].
In the Rocky Mountains, thinleaf alder-dominated riparian communities tend to occur on an elevational gradient just above narrowleaf cottonwood (Populus angustifolia) communities [281]. In western Colorado, Baker [15] noted that thinleaf alder was nearly always present at varying frequencies, and sometimes dominant, along narrowleaf cottonwood and blue spruce riparian forests. Thinleaf alder, red-osier dogwood, and water birch (Betula occidentalis) formed an often impenetrable shrub layer, with thinleaf alder dominant along stream margins and becoming less frequent with distance from the streams [15]. Thinleaf alder dominates in the upper reaches of the narrowleaf cottonwood-Scouler willow (S. scouleriana) formation in Boulder County; it may also dominate in canyon bottoms [364]. Its distribution extends into the mountain sagebrush-mountain grassland zone, mostly above 5,000 feet (2,000 m), in the Northern Rocky Mountains (reviews by [184,256]). In the Sawtooth National Recreation Area, Idaho, thinleaf alder dominates high-gradient streams (those on steep slopes or with rapid water flows) [173]. In northwestern Montana, thinleaf alder community types occur from 3,760 to 6,700 feet (1,150-2,040 m) elevation on moist stream edges, overflow channels, and slope seeps [32]. Thinleaf alder is common in many riparian shrublands dominated by other species, especially willows, red-osier dogwood, and Wood's rose (Rosa woodsii) (for example, [149,284]).
Southwest: In the Southwest, thinleaf alder occurs in wetlands within desert ecosystems and at high elevations. It may be a component of or locally dominant in riparian scrublands of Arizona; these scrublands are usually dominated by Bebb willow (Salix bebbiana), Scouler willow (S. scouleriana), and/or other willow species [40]. Thinleaf alder is an obligate riparian taxon in New Mexico, where it may dominate riparian shrublands solely or codominate with willows [96,276]. Its relative dominance generally increases with increasing elevation [95]. Thinleaf alder is a common shrub component of narrowleaf cottonwood communities of Arizona, New Mexico [20], and southern Colorado. Within wet areas, it sometimes grows as low as the pinyon-juniper (Pinus-Juniperus spp.) zone in northern Arizona [184]. In Great Sand Dunes National Monument, Colorado, it is an important component of narrowleaf cottonwood/Kentucky bluegrass (Poa pratensis) riparian communities [243]. Its distribution extends into the Engelmann spruce-corkbark fir zone (Picea engelmannii-Abies lasiocarpa var. arizonica), from 7,000 to 9,000 feet (2,000-3,000 m)), in the Rincon Mountains of Arizona (review by [184]). Thinleaf alder may be locally important to dominant in alpine riparian communities of the Southwest [40].
The following vegetation classifications describe plant communities, and some of their distinguishing site characteristics, where thinleaf alder is dominant or is a community type indicator. In state and province lists, plant communities are arranged geographically from north to south and west to east.
| Boreal North America, general | West, general | Alaska | Pacific Northwest |
| California | Southwest | Great Basin | Northern and Central Rockies |
| Northern Great Plains | Canada |
Utah (see entries in Great Basin)
Northern Great Plains |
|
Photo © 2005 Louis-M. Landry. |
Gray alder is a tree or shrub, growing from 15 feet to 82 feet (4.6-25 m) tall. [120]. Bark is smooth and thin [155,348], often with conspicuous lenticels [106] (see photo in Fire Effects and Management). The wood is soft [171]. Leaves are oblong and serrated at the margins [208,355]. The inflorescences are small, naked [131] catkins. Male catkins grow in clusters of 2 to 4. They are 0.8 to 3 inches (2-8 cm) long and pendulous at maturity. Female catkins are woody and resemble cones, growing in clusters of 2 to 6 [106,120,237]. The "cones" are 0.4 to 0.8 inch (1-2 cm) long at maturity [155]. The fruits are described as either irregular samaras [120] or nutlets with small, narrow wings [106,154,171,179,242,345]. They hold 1 to 4 seeds/cone scale [137,155,318]. The seeds lack endosperms, so the cotyledons are relatively small [154]. The root system is shallow and spreading [171]. Roots are typically infected with nitrogen-fixing, actinomycete bacteria [171,205,215,261]. A review reported that thinleaf alder fixes more nitrogen than Sitka alder (Alnus crispa subsp. sinuata) and quantities similar to those of red alder [143].
Gray alder is adapted to periodic flooding in spring or other run-off periods [23,204,267], although it cannot tolerate long periods of inundation. In the laboratory, speckled alder growth and root development were "severely reduced" when water levels were at or above the root crown for 30 days or more (P<0.05) [196,267]. A review ranked gray alder more flood tolerant than cottonwoods, birches, and elms but less tolerant than willows [130].
Morphological characteristics of the gray alder subspecies overlap (review by [143]). Thinleaf alder is typically more tree-like than speckled alder. The subspecies also differ in bark, leaf shape, and leaf margin characteristics [120,127,318].
Speckled alder is a spreading shrub [120] or small tree [131], growing up to 30 feet (9 m) tall [120] and 4.7 inches (12 cm) in diameter. Typically multistemmed with crooked branches, it is "very crooked" in form as a small tree [106], and only assumes tree form on high-quality sites [171]. Its common name refers to the lenticels that give a characteristic "speckled" look to the bark [81]. In a speckled alder community by a small stream in upstate New York, speckled alder stems averaged 14 years old, ranging from 7 to 31 years old. Stem density averaged 7,850 stems/ha [331]. In central New York, age of mature stems ranged from 10 to 25 years. Based on stem sprouting vs. stem mortality rates, the author estimated maximum age of speckled alder clones at about 100 years [176].
Thinleaf alder is an open, spreading shrubby tree or shrub, growing from 15 to 39 feet (4.6-12 m) tall [120,345] and usually less than 4 inches (10 cm) in stem diameter [215]. It often forms thickets along streams [179,231,242,345], although on upland sites it usually grows in discrete, shrubby clumps [208]. Thinleaf alder stems on sandbars of the Tanana River, central Alaska, averaged 14 years old [339]. Morris and others [259] provide a key for identifying thinleaf alder and other western shrubs in winter.
Thinleaf alder is frost-tolerant [23,204].
Raunkiaer [290] life form:
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Pollination and breeding system: Gray alder is monoecious [171,318]. The flowers are wind-pollinated [81,261], so most plants are cross-pollinated. However, thinleaf alder self-pollinates rarely (review by [154]).
Genetic differences among populations are generally small in species with wind-dispersed pollen and seeds (for example, [168]), such as gray alder. Allozyme studies of speckled alder in central Quebec showed high rates of gene flow and weak genetic differentiation among 4 populations [34].
Spatially, the genetic make-up of individuals in speckled alder thickets may be random. Studies of 4 speckled alder thickets in New York showed clones were randomly distributed, and clumps of single genotypes were not aggregated. Thus, the author concluded that although speckled alder regenerates vegetatively, sexual regeneration was driving the genetic structure of these 4 populations [175].
Seed production: Gray alder first produces seed at 25 years or younger. There are usually 1 to 4 years between large seed crops (review by [154]). Thinleaf alder produces "abundant" seed (reviews by [143,256]). Mean annual seed rain of thinleaf alder in white spruce stands of interior Alaska was 745 seeds/m² [366].
Seed dispersal: Wind and water disperse thinleaf alder seeds ([81,93], reviews by [35,312]). In waterways of Alaska, thinleaf alder seeds stayed afloat for "long periods of time" (Densmore 1976 personal observation [93]). On a floodplain of Little Otter Creek in Vermont, speckled alder seed was found in floodwaters but not in seed rain deposited on soil. The surrounding plant community was a red maple-sugar maple (Acer saccharum) forest [178].
Thinleaf alder may establish from seed in crown-stored "cones" after disturbances such as fire ([256], review by [35]), road construction, logging, and mining [256].
Seed banking: Gray alder has a transient seed bank; seed longevity in the field or in water is short [344]. Even in dry storage, seeds do not remain viable for more than 2 years [154]. In a Freeman maple-white ash (Acer × freemanii-Fraxinus americana) swamp in New York, speckled alder seed density was 0.01 seed/120 cm². Speckled alder had 0.9% cover in aboveground vegetation [28]. In dry Douglas-fir forests of south-central British Columbia, thinleaf alder seed was present in the soil seed bank on unlogged and unburned sites. Thinleaf alder seed was not found in the soil seed bank on adjacent sites that were either clearcut 5 or 10 years prior or burned at low or high severity 5 years prior. Thinleaf alder was not present in aboveground vegetation on any of the study sites [317].
Germination: Gray alder seeds are usually nondormant ([27,300], review by [19]) and, under favorable conditions, may germinate immediately after dispersal (review by [143]). However, some seed lots may require a stratification period, from a few days to over winter (review by [154]). Moist soil ([71], review by [143]) and temperatures from approximately 50 to 77 °F (10-25 °C) [154] are required for germination.
Gray alder seed viability is generally low (review by [143]), but preliminary studies suggest that light enhances germination ([93], review by [154]). Some laboratory studies found only 5% viability in thinleaf alder seeds (review by [143]). In greenhouse trials, speckled alder showed low seed viability (4-42%). Germination of filled seed varied from 0 to 50%, with presoaking and light increasing germination rates [27]. For thinleaf alder seeds collected near Fairbanks, germination averaged 90% for seeds cold-stratified in light and 5% for seeds cold-stratified in dark. Unstratified seed showed 100% germination in light and ≤13% germination in dark [93]. Gray alder seed may germinate without light, however. In a laboratory study, speckled alder showed no significant difference in time to germination and germination rate in dark vs. light, averaging 10 days to germination, and 36% germination, in both light and dark [246].
Seedling establishment and plant growth: Gray alder seedling establishment may be rare except in primary succession or on disturbed, open sites. In central New York, 4 populations of speckled alder showed no seedling establishment over 3 years [176]. In a Freeman maple-white ash swamp in New York, speckled alder establishment averaged 10 seedlings/100 m². One-year-old seedlings averaged 5.2 inches (13.1 cm) tall; 2-year-old seedlings averaged 13.5 inches (34.3 cm) tall [29].
Soil disturbance and/or exposed mineral soil favor gray alder establishment ([71], review by [143]). In Michigan, the margins of American beaver ponds were favorable sites for speckled alder germination and establishment [222]. In a Swedish field experiment, European gray alder seedling survival was higher in mineral soil (34%) than in humus (9%) [300].
Gray alder seedling establishment on new sandbars and on banks with receding floodwaters is common, although other substrates also provide favorable establishment sites. After a flood on the Connecticut River, for example, speckled alder seedlings were noted in a sugar maple swamp in an oxbow [170]. On Lassen National Forest, thinleaf alder established most often near wide stream channels on sand- or gravelbars. Thinleaf alder seedling establishment was negatively correlated with canopy cover (P=0.02) and litter depth (P=0.002) and positively correlated with solar radiation (P=0.002) [297]. In a greenhouse experiment in France, flooded European gray alder seedlings grew fastest on moist, sandy loams that were flooded and drained daily [177]. In boreal quaking aspen-paper birch ecosystems near Slave Lake, Alberta, thinleaf alder established on decaying logs and stumps [226]. Gray alder germinants apparently tolerate slightly drier conditions than willow germinants [114].
Gray alder seedlings grow rapidly under favorable conditions ([298], review by [154]). Monsen and others [256] report that thinleaf alder seedlings are "very competitive and vigorous. Once established, few plants can grow as rapidly". In central New York, annual growth increments of 4 speckled alder populations ranged from 0 to 2.8 inches (1.1 cm). Growth rate was generally greatest in midsized stems (0.8-1.5 inches (2.0-3.9 cm) DBH, P<0.01). In one population, gypsy moths removed nearly 100% of foliage in July, but stems leafed out again in late summer. Slowed growth and/or stem mortality was anticipated as a result of the defoliation but was not recorded [176]. Speckled alder seedlings proved "remarkably tough when placed in the field" after growing in a greenhouse. Ninety-seven percent survived their first year [27].
Speckled alder may grow rapidly after canopy removal (review by [143]). MacLean [236] reported that after clearcutting in boreal quaking aspen-paper birch-spruce communities in Ontario, understory speckled alder gained height growth quickly and spread vegetatively. From small clumps with few stems, speckled alder typically formed a tall, closed-shrub canopy within 10 to 12 postharvest years. Some regeneration from seed was also apparent. Best growth occurred on moist to very moist clay loams [236]. Speckled alder also grew rapidly following clearcutting on black spruce peatlands in Ontario. Ten years after harvest, speckled alder averaged 5 to 6 feet (1.5-1.8 m) tall and 16,000 to 30,000 stems/acre. Speckled alder crown closure ranged from 40% to 80% [347].
Colonizing gray alders may facilitate subsequent gray alder establishment and colony expansion. Below receding glaciers above Glacier Bay, Alaska, thinleaf alder established as widely scattered individuals. These individuals were centers of aggregation from which thinleaf alder thickets spread [75].
Ungulate browsing can substantially reduce growth of thinleaf alder on heavily browsed sites. On big game winter rangelands near the Clearwater River in northern Idaho, thinleaf alder's annual biomass accumulation averaged 0.23 g/m² in ungulate exclosures. Study plots outside exclosures, which were subject to moose, elk, and mule deer foraging, did not contain thinleaf alder, Rocky Mountain maple (Acer glabrum), or several other important browse species [4].
One study found midsized speckled alder sprouts suffered the least mortality among size classes. Over 3 growing seasons in New York, short speckled alder stems (<4.6 feet (1.4 m) tall, minimal DBH) died most often, while midsized stems (6.6-13 feet (2.0-3.9 cm) DBH) showed less mortality than larger sprouts. Sampling was conducted in 4 early-successional speckled alder thickets. Variation in stem mortality and growth was greater within populations than across sites (P<0.05 for all variables) [176].
Vegetative regeneration: Cloning is apparently the primary means of spread in established stands of gray alder (review by [143]). Gray alder sprouts from the root crown [35,38,175,208]. It can also sprout from roots, including root offsets [175,176,298], and layers. Root sprouting and layering are apparently less important than root-crown sprouting [176], although root sprouts distant from the parent plant have been noted for both speckled alder and European gray alder (review by [143]). Haeussler and others [143] report that sprouting "can be expected" after mechanical site preparation. In a Magellan's sphagnum bog in Ohio, speckled alder sprouted after cut-stump herbicide applications, growing up to 3 feet (1 m) in one growing season [9]. Gray alder does not spread rapidly, but its clones can be long-lived. In a central New York study, 4 speckled alder populations were monitored for 4 years. There was no seedling establishment on study plots during that time. Clump sprout production and stem mortality were variable within and among populations, although no clumps died out. No lateral extension of clones via root sprouting occurred. Annual sprout production averaged about 3 live stems/clump [176].
In a study comparing root anchorage of riparian species in Italy, European gray alder was less resistant to uprooting by flood than Lombardy poplar (Populus nigra) or Elaeagnus willow (Salix elaeagnos) [191]. This relative ease of uprooting may allow for vegetative spread of gray alder when root fragments are torn off, distributed downstream, and sprout.
SUCCESSIONAL STATUS:Speckled alder
Speckled alder prefers open sites [106,171,205] but tolerates moderate shade (Shirley 1932 cited in [39]). Studies in Michigan found that it occurred on open to closed sites but was most common on lightly shaded, cool sites [16,36]. In southern-boreal, quaking aspen forests of Quebec, speckled alder was associated with relatively high light transmission at 7 to 13 feet (2-4 m) above ground level (P<0.001). Thirty percent of full sun was the highest light level achieved on most sites [18].
Speckled alder is an important shoreline and meadow colonizer, and it is successional in mires and other wet to damp areas throughout its range [120]. In Michigan, it colonizes sand dunes by Lake Huron [13]. In New York, it is a characteristic taxon on unstable beachgrass (Ammophila breviligulata) dunelands [293]. On an estuary on the St Lawrence River, Quebec, speckled alder seedlings and saplings showed 28% cover, the greatest of any colonizing shrub taxa. Drifting ice, wave action, and frequent flooding inhibited establishment of less tolerant shrubs along the shoreline [24]. By upland oak-hickory communities, floodplains with speckled alder generally succeed to sycamore, elms, and red maple (review by [160]). Speckled alder may be invasive in sedge (Carex spp.) wetland meadows of Wisconsin, especially with fire exclusion [294]. Speckled alder is noted in the older zones of bogs in Michigan [350]. In bog succession, speckled alder generally establishes after litter from mosses and low ericaceous shrubs has formed a peat layer. Without disturbance, trees such as tamarack and black spruce establish after speckled alder, willow, and other shrubs [303]. Speckled alder may be the pioneer woody taxon in swamps that succeed to red maple [176].
The successional pathway of acidic mires and other wetland communities may alter after speckled alder establishment [9,38]. Since speckled alder is a nitrogen fixer, acid-loving bog and fen species may be replaced successionally by species with higher nutrient requirements. On a kettle bog in Ohio, speckled alder established on a Magellan's sphagnum mat and was displacing acid-loving herbs such as purple pitcher-plant (Sarracenia purpurea) and round-leaved sundew (Drosera rotundifolia) [9]. Speckled alder is more characteristic of fen and bog margins than of the mires themselves, although it sometimes establishes in mires with peaty soils [89]. On the Lac St-Francois National Wildlife Area in Quebec, speckled alder invaded a water sedge-hairy sedge (C. lacustris) fen. Over several decades, speckled alder became dominant over about 10% of the wetland that had formerly been sedge fen [38]. Woody invasion of herbaceous bogs and fens in eastern North America has been attributed to fire exclusion [9,38], although altered hydrologic regimes may also play a role [38]. A 1938 study of the speckled alder-willow high bog association in Cheboygan County, Michigan, found speckled alder and willows were casting dense shade and becoming "scraggly". They were being replaced successionally by red-osier dogwood, black spruce, tamarack, and northern white-cedar. Charred logs and stumps in the bog indicated that fire had cleared the area of trees in the past [110]. In contrast to speckled alder replacing herbs successionally in wetlands, beaked sedge has established and spread in speckled alder-withe-rod (Viburnum nudum var. cassinoides) communities of east-central West Virginia [90].
In forests, speckled alder may be important after logging, insect attacks, and/or disease. On logged black spruce forests of northeastern Ontario, speckled alder was most common in forests <40 years old, forming dense shrub layers of up to 570 to 600 stems/0.01 ha. In older stands, speckled alder was less important, forming a "diffuse canopy" on most sites. It remained dense in seeps and by open water, however. The authors surmised that speckled alder was probably present in low numbers in the prelogged forest and increased rapidly after logging [43,44]. Speckled alder was prevalent after an eastern spruce budworm attack and subsequent clearcutting in balsam fir-paper birch communities in Ontario. Speckled alder density ranged from 109 stems/ha to 6,667 stems/ha on 8- to 12-year-old clearcuts [158]. In early secondary succession, deforestation due to Dutch elm disease tended to favor speckled alder and other shrubs [108]. In hardwood swamps of central New York, speckled alder was "frequently encountered" in gap succession following death of overstory American elms to Dutch elm disease. It occurred in both small gaps created by the death of single trees and in larger gaps created by deaths of multiple trees [174].
Speckled alder is generally unimportant in late-successional forests. It was a mostly minor component of late-successional eastern hemlock-black spruce-red maple forests of northeastern Pennsylvania, attaining high density and cover only in canopy gaps [100]. However, a 1934 publication reported speckled alder as characteristic of climax balsam fir forests in Itasca County, Minnesota. It was also present in earlier succession [136].
Speckled alder may be of minor importance in old field succession. It formed thickets on old fields in New York [49,175,176]. In southeastern Ontario, speckled alder seedlings were found 19 years after abandonment of ploughed hay fields, although in low numbers (<1% frequency). Speckled alder was also present in adjacent silver maple/roundleaf serviceberry (Acer saccharinum/Amelanchier sanguinea) forest. It presumably established in the old field from wind-blown seed originating from the forest [84]. In an old-field study, speckled alder was an early-seral shrub on the Piedmont Plateau of North Carolina. It preferred wet bottomlands that had been used as hayfields, sometimes forming dense, shoulder-high thickets. Speckled alder was also noted in old fields 34 and 45 years after abandonment on a site succeeding to loblolly pine (Pinus taeda) forest. Its density in the loblolly pine habitat ranged from 1.9 stems/16 m² on a 34-year-old field to 4.1 stems/16 m² on a 45-year-old field. In an adjacent old field on logged streambanks succeeding to paper birch, speckled alder was present 6 and 14 years after abandonment at densities of <2 stems/16 m². In old fields succeeding to mixed-hardwood bottomlands, it was found in 8- and 32-year-old fields (densities not recorded). None of the old-field plant communities surveyed was older than 55 years [270].
Thinleaf alder
Thinleaf alder is moderately shade tolerant. Thinleaf alder is adapted to nearly all types of disturbance [151], including severe disturbance [32], and is most common in early succession. It can grow in forest understories, but it is found more often on open sites. Sprouts may tolerate shade better than seedlings (review by [143]). To date (2011), most successional studies on plant communities with gray alder had been conducted on Alaskan sites in primary succession.
Thinleaf alder is an early-successional shrub in riparian zones in primary succession [93]. The willow/alder stage typically forms on bare floodplains [362]. On the Tanana River in interior Alaska, it typically establishes on bare to nearly bare, recently deposited alluvium [1]. Feltleaf and/or sandbar willow may establish first [1,55]. Thinleaf alder and balsam poplar dominate in midsuccession, after which balsam poplar and finally, white spruce, dominate the overstory [1]. A 5-year-old thinleaf alder sandbar community on the Tanana River was a "nearly impenetrable" thicket of thinleaf alder, feltleaf willow, sandbar willow, and barrenground willow. The shrubs were uniformly about 10 feet (3 m) tall, 0.8 inch (2.0 cm) in stem diameter, and averaged 49,699 stems/ha. Balsam poplar, paper birch, and white spruce grew in the understory; meadow horsetail and variegated scouringrush were dominant herbs. In 15-year-old stands, feltleaf willow was overtopping thinleaf alder, and shrub density declined to 2,827 stems/ha. Groundlayer vegetation was nearly all meadow horsetail [162]. By Glacier Bay, Alaska, thinleaf alder occurs about 15 to 20 years after glacier recession, in the "late pioneer" stage. About 25 to 30 years after recession, thinleaf alder forms closed stands, presenting an "almost impenetrable barrier". At 30 to 35 years, black cottonwoods begin to establish [337].
According to a 1923 study by Cooper [76], thinleaf alder may be abundant even in late succession after glacier recession. Herbs, especially dwarf fireweed (Chamerion latifolium), dominated the northernmost, pioneer community, but thinleaf alder and willows were also establishing. Feltleaf willow-Sitka willow-thinleaf alder communities occurred in isolated patches and on midlatitude landscapes; midlatitude sites had a longer period of recession than the northern sites. Thinleaf alder was "nearly everywhere dominant" and eventually overtopped the willows. Sitka spruce forests occurred in the southernmost portion of the landscape, which had the longest time since glaciation. Thinleaf alder persisted in these late-successional forests, typically at greater abundance than in the pioneer stage [76].
The pattern of willows, and sometimes cottonwoods, establishing before thinleaf alder is typical in riparian succession. On bare gravel bars of Meadow Creek on the Starkey Experimental Forest, Oregon, thinleaf alder established at lower densities (0.96 stem/50 m²) than sandbar willow and black cottonwood [60]. Along the Animas River in southwestern Colorado, narrowleaf cottonwood/thinleaf alder communities occur upland from narrowleaf cottonwood/tickle grass communities and are considered a latter successional community type than the narrowleaf cottonwood/tickle grass community [351].
Thinleaf alder shrub communities are initiated and maintained by disturbances that are often severe. Historically in northwestern Montana, disturbances have included placer mining, ice jams, log transport [32,151], and fire. On Emigrant Creek near Burns, Oregon, thinleaf alder colonized a new alluvial fan 6.5 years after deposition following an intense thunderstorm. Cattle had grazed the area for at least 30 years [145]. Thinleaf alder snowslide communities in the Blue Mountains are maintained by avalanches and soil slippage [144]. Along the San Miguel River in southwestern Colorado, thinleaf alder shrub communities occupied less area than that of later-successional, narrowleaf cottonwood/thinleaf alder communities. Thinleaf alder shrubland also had shorter flood-return intervals (averaging <10 years) than those of narrowleaf cottonwood/thinleaf alder communities (averaging about every 50 years) [126].
Thinleaf alder may facilitate establishment of later-successional riparian species, likely due in part to its ability to fix nitrogen. On floodplains of interior Alaska, for example, balsam poplar and feltleaf willow establish in nitrogen-enriched soil beneath thinleaf alder, although root crowding and shading by thinleaf alder may interfere with growth of later-establishing species on many floodplain sites (reviews by [53,54]). On floodplains near Fairbanks, there was a "marked" increase in exchangeable soil potassium, calcium, magnesium, manganese, and phosphorus within 5 years of thinleaf alder establishment, but there was no consistent increase in soil mineral content from 5 to 20 years after thinleaf alder establishment. Soil pH decreased beneath thinleaf alder stands over 20 years, while soil cation exchange and organic matter increased. At study year 20, most of the total aboveground plant biomass (~43 kg/ha of ~48 kg/ha total) and basal area (7,142 stems/ha of 7,241 stems/ha total) was thinleaf alder [338].
Experiments on the Tanana River floodplain showed changes in soil nutrient dynamics and soil microbe community composition as aboveground succession proceeded from thinleaf alder to balsam poplar. As balsam poplar gained dominance, soil carbon became increasingly more available and soil nitrogen became increasingly less available to soil microbes. Thus, the soil biota changed from carbon-limited microbes under thinleaf alder dominance to nitrogen-limited microbes under balsam poplar dominance [70].
Thinleaf alder occurs in all stages of forest succession, although it is most prevalent in early forest succession. It pioneers in forest communities of British Columbia, sometimes persisting in mature forests [23]. Thinleaf alder is seral in Douglas-fir, spruce, and other coniferous forests in the West (for example, [150]). In forested habitats, the thinleaf alder shrub community is usually an early- to midsuccessional seral stage that arises after severe disturbance. Conifers or taller hardwoods typically replace thinleaf alder successionally [151]. Near Slave Lake in Alberta, thinleaf alder sprouts were more common on sites logged 28 years prior than adjacent sites burned by wildfire 28 years prior (P<0.05). Study sites were dominated by quaking aspen, balsam poplar, and paper birch [225]. Thinleaf alder often dominates the understory of midseral spruce/bluejoint grass forest habitat types in Montana [151]. In northwestern Montana, thinleaf alder was abundant in Engelmann spruce/skunk cabbage riparian communities from early-seral (logged and heavily grazed) to late-seral and climax stages. It dominated early stages of succession in Engelmann spruce/field horsetail and Engelmann spruce/bluejoint reedgrass riparian communities [32]. In central Alaska, thinleaf alder thickets are often replaced successionally by balsam poplar, which in turn is replaced by white spruce, then black spruce [339]. Thinleaf alder occurred late in the succession of white spruce-Engelmann spruce forests on the east slope of the Rocky Mountains in Alberta [78]. By the Peace River in northern Alberta, thinleaf alder was dominant in old-growth white spruce forests. It was most common in canopy gaps, although it persisted in the understory. It also occurred in resin birch clearcuts [332]. In central Alberta, thinleaf alder was more common in mature quaking aspen-balsam poplar-white spruce forest than on edges of 16-year-old clearcuts (P=0.05) [153].
Browsing pressure may alter succession in forest ecosystems with thinleaf alder. Browsing may favor thinleaf alder at the expense of more palatable browse species. In spruce-birch taiga forests of interior Alaska, browsing pressure by moose favored thinleaf alder and quaking aspen over more palatable willow species [52]. In northern Idaho, ungulate browsing helps maintain shrubfields, which contain thinleaf alder and other seral shrubs. Heavy browsing, however, may accelerate succession to conifer species, which are less palatable [4]. |
|
Immediate fire effect on plant:
Gray alder's thin bark [155,348] does not insulate it well from fire damage. Fire generally top-kills gray alder ([38,164,346], reviews by [35,92,312]).
Postfire regeneration strategy [319]:
Tree with adventitious buds, a sprouting root crown, and
root sprouts
Tall shrub, adventitious buds and a sprouting root crown
Geophyte, growing points deep in soil
Crown residual colonizer (on site, initial community)
Initial off-site colonizer (off site, initial community)
Secondary colonizer (on- or off-site seed sources)
Ground residual colonizer (on site, initial community)
Fire adaptations and plant response to fire:
Fire adaptations:
Gray alder has many adaptations that can aid its postfire recovery. It sprouts from the root crown and/or roots after top-kill. Root crown sprouting is most common (see
Vegetative regeneration),
while root sprouting may occur when fire is severe enough to kill the root crown. Gray alder may also establish from seed; this may include seed dispersed from on-site, crown-stored "cones" ([256], review by [35]), wind- or
water-dispersed seed
originating from off-site parents, or seed in the transient soil
seed bank .
Some of gray alder's adaptations to riparian environments, including tolerance of high light and the ability to sprout after repeated top-kill, may also benefit gray alder in early postfire environments [209].
A prescribed April fire top-killed most speckled alders in a study site on the Allegheny River floodplain of Pennsylvania. Open areas with individual trees burned well, but dense stands of speckled alder carried fire mostly on the edges. The fire top-killed 55% of speckled alder stems. By postfire month 9, most burned speckled alders were sprouting at or within 2 inches (5 cm) of the soil surface, from near the bottoms of the stems or from the root crowns. However, sprouting in speckled alder was not related to fire; it occurred on burned and unburned stems at approximately the same rate, 90%. Sampled stems produced an average of 8 to 10 sprouts, with basal diameters ranging from 0.1 to 0.5 inch (3-12 mm) and heights ranging from 9 to 47 inches (22-120 cm). Twelve percent of speckled alder stems were browsed, probably by white-tailed deer and cottontails [49]. See the Research Project Summary of this paper for information on the burning conditions and effects of the fire on hawthorns (Crataegus sp.) in the same area.
A few studies show speckled alder may be more common in early than late postfire succession. In a fire chronosequence study in black spruce-jack pine ecosystems of northern Ontario, speckled alder was most common on wet peatlands burned 1 to 3 years prior (P=0.05). Burn ages ranged from 0 to 57 years [308]. Speckled alder reached greatest postfire abundance in postfire year 10 in red pine-black spruce forests of Saskatchewan [306]. In a chronosequence of 9 burned black spruce/mountain-laurel (Kalmia latifolia) forests in Terra Nova National Park, Newfoundland, speckled alder was present only on a 20-year-old burn, with 2% mean cover. Burn ages ranged from 1 to 38 years [30]. In quaking aspen, jack pine, and black spruce swamp communities of Itasca County, Michigan, speckled alder occurred in 7 of 9 burns surveyed. The burns were from 1 to about 58 years old [135]. A review of fire succession in balsam fir forests of the Northeast reported that speckled alder thickets may replace balsam fir after fire and that those thickets may persist for decades before trees establish [128].
Frequent fire likely favors speckled alder over conifers. Speckled alder thickets may form after stand-replacing fire in northern white-cedar communities [186]. In a northern white-cedar swamp in Michigan, speckled alder was most abundant in an area that burned 3 times in 30 years. The authors reported that dense growth of speckled alder and willows now occur on sites where the northern white-cedars experienced the greatest fire kill [110].
Speckled alder increased over 10 postfire years on a hybrid spruce clearcut that was slashburned at low to moderate severity. The site was on the Mackenzie Forest District in subboreal British Columbia. Speckled alder was the dominant shrub before and after the fire, with abundance at postfire year 10 exceeding its prefire cover and frequency. Speckled alder's prefire cover and frequency were 4.6% and 50%, respectively. At postfire year 10, it had 8.9% cover and 67% frequency. Overall, shrub height was more in postfire year 10 than before fire, primarily due to the rapid postfire growth of speckled alder and Rocky Mountain maple. See the Research Paper of Hamilton's [146] study for details on the fire prescription, fire behavior, and postfire responses of other species in the plant community.
Two studies suggest that speckled alder responds similarly to fire and logging. Pooled across plant communities and stand ages, there was no significant difference in speckled alder frequency between wildfire-burned (37.0%) and logged (31.4%) stands in Minnesota. Surveys were conducted in naturally regenerated quaking aspen, jack pine, and black spruce communities that were 25 to 100 years old [292]. In a black spruce forest in west-central Quebec, there was no significant difference in speckled alder cover on the edges of burns and clearcuts. Burns were 3 to 4 years old, and clearcuts were 2 to 5 years old [152].
Both wildfire and clearcutting apparently favored speckled alder; its average frequency was higher than that of any other deciduous shrub after both disturbances in surveys in northeastern Ontario [57]. Speckled alder was more frequent after clearcutting than after wildfire, but a longer period of postdisturbance succession on burned vs. clearcut stands makes direct comparisons difficult. Time since fire averaged 89.6 years, while time since logging averaged 22.9 years: Given speckled alder's role in early succession in this area, greater abundance in the younger, logged communities seemed likely. On clearcut stands (n=131), speckled alder averaged 6.9% frequency in the canopy and 18.47% cover in the subcanopy. Both wildfire and clearcutting apparently favored speckled alder; its average frequency was higher than that of any other deciduous shrub after both disturbances [57].
Thinleaf alder
Thinleaf alder sprouts from the root crown after top-kill by fire ([38,164,346], reviews by [35,92,312], observations reported in [119]). Stem density may increase after fire due to multiple sprouts arising from single root crowns (review by [312]). A review of fire responses of British Columbian shrubs states that thinleaf alder is "set back by moderate or severe fires" [159]. An anecdotal account from central Oregon reports that in thinleaf alder-dominated riparian communities, thinleaf alder withstands low-severity surface fires but is killed by more severe fires [200]. Root sprouting is possible, however, and further investigations are needed to ascertain thinleaf alder's response to moderate and severe fires.
Thinleaf alder establishes on burns from on- or off-site, wind- or water-dispersed seed (reviews by [35,312]). Wind may disperse seed onto burns from "considerable distances" [312]. Seedlings may also establish from seed in crown-stored "cones" after late summer fires (reviews by [35,312]). Viereck and Schandelmeier [346] reported that fire usually kills crown-stored seed in Alaskan alders such as thinleaf alder, but alder seeds are typically blown onto burns from off-site parents. Alders in lightly burned areas may produce "large numbers" of seeds in early postfire years [346]. Thinleaf alder may establish in early postfire years after stand-replacement fires in lodgepole pine. In the Bitterroot Mountains of northern Idaho, thinleaf alder often establishes from wind-blown seed around postfire year 3 [217]. Establishment from soil-stored seed may also occur (see Seedling establishment).
In a greenhouse study using soil from a wildfire-burned area near Slave Lake in Alberta, thinleaf alder sprouted from the roots but did not emerge from the seed bank. Soil samples were collected a week after the fire (cut in blocks 27.4² inches (177 cm²) across and least 4 inches (10 cm) deep). Thinleaf alder sprouted from soil blocks where fire had been "intense", averaging 23% cover after 2 years in the greenhouse. Thinleaf alder did not sprout from soil blocks collected from lightly burned or unburned sites. The plant community was mixed hardwood-conifer dominated by quaking aspen [224].
After summer wildfires on the Plumas National Forest, California, fewer thinleaf alders sprouted on upland sites—where fire crowned—than on gravelbars, where fire was less severe. Postfire response of vegetation on 2 streams, Third Water and Fourth Water creeks, was evaluated in postfire year 1. On the 1st and 2nd terraces above the creeks' gravelbars, the plant community was an incense-cedar-white fir/Eastwood manzanita-huckleberry oak (Arctostaphylos glandulosa-Quercus vaccinifolia) forest. Thinleaf alder dominated the gravelbars [197].
| For each topographic position, mean relative density of thinleaf alder and percent of burned thinleaf alders that sprouted along Third and Fourth Water creeks in postfire year 1 [197] | ||||||||
| Topographic position | Gravelbar | 1st terrace | 2nd terrace | Riparian-zone slope | ||||
| Site | % of total | % sprouting | % of total | % sprouting | % of total | % sprouting | % of total | % sprouting |
| Third Water Creek n=47 plants |
100 | 100 | 48 | 30 | 38 | 54 | 34 | 67 |
| Fourth Water Creek n=25 plants |
23 | 100 | 40 | 90 | not present | 4 | 0 | |
On hybrid spruce-Engelmann spruce plantations in subboreal British Columbia, thinleaf alder abundance was similar on clearcuts prepared for planting by either prescribed burning or mechanical treatment (blading). Across treatments, mean thinleaf alder cover ranged from 17% to 25% on plantations <7 years old [147].
In a study of disturbed forests near the Peace River in northern Alberta, thinleaf alder was strongly positively associated (P<0.001) with frequently flooded sites (~1-10 years) but not with recent burns (5- to 20-year-old) or older burns (>20 years). Most (48%) of the white spruce and hardwood forests originated after flooding; fewer regenerated after logging (32%) or stand-replacing fire (19%). Thinleaf alder was recorded only on flooded sites. It was typically present in the earliest seral stages after flooding, establishing on gravelbars, levees, and the lowest stream terraces [332].
Thinleaf alder may occur in late postfire succession. On 12 white fir sites in Sierra County, California, that had experienced stand-replacing wildfires, thinleaf alder occurred only on study plots that had burned 80 years or more prior to the study. Time since fire ranged from 5 to 277 years [72].
In sagebrush ecosystems of the West, mesic thinleaf and other alder shrublands may establish after fire, or they may be self-sustaining [322].
FUELS AND FIRE REGIMES:Fuels: Because gray alder adds nitrogen to soils and typically grows in moist soils, gray alder communities are usually highly productive [132]; hence, their fuel loads can be large. Gray alder may produce a "dense" litter layer [353]. However, because gray alder prefers moist sites, in most years gray alder communities may act as firebreaks. Riparian zones with gray alder are often buffer zones where upland fires decrease in severity or stop [285]. Speckled alder and thinleaf alder communities are placed in fuel model 0 [282] and fire group 0 [312], respectively; these plant communities do not burn readily [282,312]. Communities in fuel model 0 are described as vegetation types in which fire will not carry due to saturated ground or standing water, discontinuous fuels, and/or lack of ladder fuels [282]. However, with dry weather and the accumulation of dry fuel, riparian areas can become corridors for fire spread [285].
Speckled alder: A prescribed burning guideline for Ontario reports that speckled alder communities do not burn well under conditions needed for prescribed fires. McRae [248] found that in boreal black spruce-white spruce-quaking aspen forests of Ontario, prescribed fires did not typically spread into drainageways dominated by speckled alder. Because there was no combustible slash, these drainages acted as fire barriers [248]. An extension publication recommends speckled alder as an ornamental due to its low flammability [7].Several publications provide help for estimating fuel loads in speckled alder habitats. See these publications: [314,329] for information on models to estimate speckled alder biomass in the northern United States and Canada. Buech and Rugg [47,48] provide equations to predict aboveground biomass of speckled alder. Their model was developed in northern Minnesota [47,48]. A photoseries for assessing fuels in coniferous forests of northern Ontario provides size class breakdowns and photos for live and woody surface fuels in black spruce, black spruce-tamarack, black spruce-jack pine, black spruce-poplar (Populus spp.), balsam fir-black spruce, paper birch-white spruce, and poplar-white birch stands [321], all of which may include speckled alder as a component of the vegetation (see Site Characteristics and Plant Communities).
In the laboratory, speckled alder's heat of combustion and total heat release were about average for shrubs of the Northeast [94].
Several studies provide what may be representative examples of fuel loads in speckled alder habitats. In a speckled alder/goldenrod (Solidago spp.) community in upstate New York, speckled alder production averaged 730 g/m²/year. Leaves and twigs comprised 42% of total production; 25% was in the boles; 21% in branches; and 12% in fruits. Herbaceous production averaged 241 g/m², yielding a total aboveground production of 971 g/m² for the speckled alder community [331]. A New York study found total speckled alder production averaged 2,225 g/m², 21% of which was roots [349].
In commercial forests in Michigan, speckled alder biomass was least in oak-hickory and greatest in tamarack stands [273]. In a similar study across commercial forests in the Northern Great Lakes (northern Minnesota, northern Wisconsin, and the Upper Peninsula of Michigan), speckled alder biomass was least in jack pine and greatest in balsam fir stands [313]. No information on stand ages was available.
| Speckled alder biomass by forest type | ||
| Forest type | Michigan [273] | Northern Great Lakes [313] |
| Biomass (lbs/acre) | ||
| oak-hickory | 13 | no data |
| maple-birch | 35 | 239 |
| elm-ash-maple | 607 | 2,584 |
| aspen | 134 | 1,989 |
| paper birch | 283 | 552 |
| red pine | 24 | 383 |
| jack pine | 71 | 153 |
| eastern white pine | 118 | 3,368 |
| white spruce | 260 | 1,758 |
| northern white-cedar | 433 | 4,221 |
| balsam fir | 644 | 5,482 |
| black spruce | 1,703 | 4,090 |
| tamarack | 2,607 | 3,037 |
In Nova Scotia, speckled alder biomass averaged 83 kg/ha in dense mixed-conifer, 140 kg/ha in dense mixed-hardwood, and 1,150 kg/ha in open mixed-conifer forests [328].
Thinleaf alder: Thinleaf alder thickets are somewhat fire-resistant because the duff is usually cool and moist, and the undergrowth is sparse [92]. The generally moist conditions in thinleaf alder stands usually inhibit fire spread (review by [312]). Even when fires are severe in upland areas, riparian zones may remain unburned or burn with lower severity. However, fires may burn severely in riparian areas in drought years [315]. A fire management plan for Lassen National Park, California, reported that natural fires seldom ignite in black cottonwood-thinleaf alder woodlands; instead, those woodlands are generally firebreaks. However, old stands with diseased and downed trees may be "quite flammable", especially if white fir or red fir has formed ladder fuels in the understory [181].
In many forested riparian areas, live and dead fuel structure and loads have altered due to fire exclusion. Arno and Harrington [12] report that in riparian ponderosa pine communities of the West, where thinleaf alder is known to occur, most sites are dominated by dense, shade-tolerant trees such as grand fir, and the shrub and ground layers are sparse.
On 3 thinleaf alder wetland communities in west-central Montana, total aboveground plant productivity ranged from 3,320 to 4,820 lbs/acre. Of that total, shrubs averaged 316 lbs/acre, forbs averaged 1,200 lbs/acre, and graminoids averaged 2,713 lbs/acre [286].
Several studies described below provide what may be representative examples of fuel loads in thinleaf alder habitats by the Tanana River in Alaska. Yarie and Mead [363] present models to estimate twig and leaf biomass of thinleaf alder and other species of the Tanana River Basin.
On sandbars of the Tanana River, total production in 5-year-old thinleaf alder thickets averaged 20,000 kg biomass/ha (including root biomass). Production peaked in 15-year-old thickets at an average of 65,000 kg/ha, while 20-year-old thickets averaged 95 kg/ha [339]. On thinleaf alder-dominated floodplains of the Tanana River, thinleaf alder biomass was almost 5 times greater in 20- vs. 5-year-old stands. Codominant willows and horsetails comprised much smaller portions of the total plant community biomass (3.2-8.7%) [339].
| Distribution of biomass (kg/ha) in thinleaf alder-dominated stands of different ages [339] | |||
| Sample | 5 years | 15 years | 20 years |
| Thinleaf alder biomass* | 8,751 | 27,810 | 42,741 |
| % of total plant community biomass | 42.9 | 43.5 | 45.1 |
| Total down woody fuel biomass | not available | 10,464 | 16,389 |
| Total litter biomass | 4,340 | 4,134 | 4,390 |
| Total biomass | 20,388 | 63,876 | 94,689 |
| *Above- & belowground biomass mass was measured for live plants. | |||
After late February logging, a prescribed broadcast burn in July reduced fuels in a white spruce/thinleaf alder/feathermoss (Hylocomium sp.) forest on the Tanana River floodplain. The fire "substantially" decreased organic material on the soil surface and exposed mineral soil. Down and dead woody fuels were reduced an average of 67%, mean depth of the forest floor was reduced 43%, and the soil organic layer was reduced 2.9%. See Zasada and Norum [367] for information on the fire prescription and fire weather conditions.
Fire regimes: Gray alder experiences a wide variety of fire regimes across its broad distribution in North America. For example, ponderosa pine woodlands in the Pacific Northwest and California historically experienced mostly low-severity surface fires with an average return interval of 13 years [210], while mixed-hardwood-spruce forests of the Northeast historically experienced mostly stand-replacement fires at intervals of 400 years or more [212]. As an early-successional species that sprouts after fire, gray alder is likely to flourish in wetland plant communities where fire is frequent or severe enough to maintain or create open conditions. This is likely true across the species' worldwide distribution. Scots pine-Norway spruce (Pinus sylvestris-Picea excelsa) forests of Scandinavia, where European gray alder is an important component of the vegetation, historically experienced frequent, low-severity surface fires and mixed-severity fires under a fire regime similar to that of ponderosa pine and mixed-conifer forests of the western United States. Frequent understory fires helped maintain European gray alder in the understory (review by [10]). Speckled alder: Little information was available on fire frequency and fire behavior in riparian and wetland plant communities where speckled alder is an important to dominant member of the plant community. One study, near Thunder Bay in Ontario, found wildfire did not carry well once it reached a speckled alder community. A stand-replacement wildfire burned a mixed hardwood-coniferous forest, but the fire slowed or stopped in riparian zones dominated by speckled alder. Speckled alder showed scorching and damage to top branches but was otherwise unharmed, and "little damage was evident in the riparian zone" [209]. Research is needed on fire regimes of riparian and wetland plant communities in the eastern United States.Summaries of several fire studies in plant communities where speckled alder was important or dominant follow.
Red-white-jack pine ecosystems of the Great Lakes states historically experienced frequent, low-severity surface fires. Frequent fires prevented development of late-successional tree species such as white spruce and red maple, and also inhibited spread of understory shrubs including speckled alder, mountain alder (A. viridis subsp. crispa), beaked hazelnut, and American hazelnut. Heinselman [165] reported that in these ecosystems, both the late-successional tree and the shrub layers are maturing and expanding under fire exclusion. A fire history study on Pictured Rocks National Lakeshore, Michigan, found red-white-jack pine shoreline forests with speckled alder had a mean fire-return interval of 21.8 years before European settlement, with fire-return intervals lengthening after the early 18th century. At the time of study (1985), the area had not experienced fire for the past 84 years [232].
A study of fire scarring in red and jack pines near Lake of the Clouds in northern Minnesota found fire-return intervals ranging from 16 to 62 years between 1691 and 1818. Speckled alder was a dominant shrub in the area. Over the previous 1,000 years, lake sediment analyses showed a mean fire-return interval of about 70 years; speckled alder pollen was present in most sediment cores sampled across the 1,000 years [323]. A similar lake sediment study in Boundary Waters Canoe Area also found a mean fire-return interval of about 70 years over the past 1,000 years; speckled alder pollen was present in all cores sampled across that time. The extant plant community was a quaking aspen-paper birch/speckled alder forest [324].
Red spruce-balsam fir-white spruce forests of the Northeast historically experienced infrequent, stand-replacement fires at 150- to 300+-year intervals. Surface fires were "extremely uncommon". Speckled alder is an early-successional species in these forests [211].
The black spruce/speckled alder forest cover type of boreal North America is reportedly a stable type that regenerates into forests of similar composition after fire [87]. Information on fire regimes for this type was not found in the literature. In general, boreal black spruce forests experience stand-replacement fires at intervals of 50 to 150 years [165].
A study by Jacobson and others [182] suggests that historically, infrequent fires may have reduced encroachment of speckled alder and other woody species into open mires in the Northeast. Frequent fires, however, may favor sprouting woody taxa such as speckled alder. A study at Crystal Fen in northern Maine suggested that frequent fires combined with high drainage favored encroachment of speckled alder and other woody species. Charcoal analysis of the fen showed fire was uncommon before a railroad was constructed in the area. Fire frequency increased after 1937, when a drainage ditch was excavated next to a railbed, drying out vegetation, and sparks from diesel-powered trains ignited fires frequently. A survey of tree ages showed that "many" speckled alder, northern white-cedar, and other woody species established between 1937 and 1950. Fire frequency dropped after 1950, when diesel engines replaced steam engines, ignition sources became uncommon, and for undescribed reasons, the fen began flooding again. Increased moisture and no fire, however, did not open the fen: Woody species continued establishing on areas of the fens that, based on pollen and charcoal records, historically had few to no woody plants. The authors recommended restoring the historical drainage pattern, cutting woody species, and/or introducing prescribed fire to restore the open fen [182].
As of 2011, almost no literature was available on fire regimes of pine or hardwood ecosystems of the Southeast where speckled alder formed a substantial component of the vegetation. A review reported that pond pine and bay (Magnolia and Persea spp.) shrubland bogs, of which speckled alder is a component, burn at irregular, 5- to 50-year intervals. Nearby bog communities with low shrub diversity tend to have less frequent fire-return intervals than the bog communities with high shrub diversity [356].
See the Fire Regime Table for speckled alder for further information on fire regimes of vegetation communities in which speckled alder may occur. Find further fire regime information for the plant communities in which this species may occur by entering the species name in the FEIS home page under "Find Fire Regimes".
Thinleaf alder: Historical fire regimes for riparian zones of the western United States are not well studied [307,311], although more is known of fire regimes in riparian and wetland areas of the West than those of the East. Historically, thinleaf alder glades and thickets in the Northern Rocky Mountains probably burned less frequently than surrounding coniferous forests, but at "higher intensity" than surrounding vegetation. Davis and others [92] speculated that thinleaf alder thickets may be maintained by infrequent fire. In most years, thinleaf alder thickets in riparian zones do not burn [92]. Similarly, thinleaf alder stands growing on seeps and springs may burn rarely [200].
In general, fires are less frequent in riparian and wetland areas of the West than in drier areas [12]. At low- to midelevations in the southern Cascade Range and Klamath Mountain regions of California, fire-return intervals in riparian areas overall are about twice those of surrounding areas, but fire intensity is generally greater when the riparian areas burn [307]. Fire-return intervals near intermittent and ephemeral streams are likely similar to those of surrounding areas [311]. In most years, perennial stream communities may serve as firebreaks (review by [311]). A fire management plan for Craters of the Moon National Monument, Idaho, states "there is no evidence that fire is necessary to maintain the (riparian) vegetation type in this area". However, the authors speculated that infrequent, periodic fire would probably improve the health and productivity of shrubs and that most shrubs, including thinleaf alder, would survive fires that were "not too frequent" [17]. Arno and Harrington [12] suggest that prior to 1900, riparian ponderosa pine communities in the western United States burned 2 to 5 times per century. Fire-return intervals in riparian and wetland communities may vary across time and space, however.
In riparian zones where thinleaf alder is known or likely to occur, fires tend to have longer return intervals, and burn at higher severity, than in upland sites ([115,268,310]), Arno 2001 personal communication [11]). This trend is not strong or consistent in all areas, however. Fire frequency and severity have been lower in some riparian areas compared to adjacent upland areas; other sites have similar fire histories on riparian and upland zones. Factors affecting fire-return intervals in riparian zones include soil and plant moisture, fire exclusion, livestock grazing, logging, damming and other water-flow regulation, and presence of invasive species that alter fuel characteristics [111]. Coniferous riparian forests may have historically been too moist to burn in some years; consequently, low- and moderate-severity fires in upland areas would often extinguish at the riparian zone. In an Alberta study of upland and riparian lodgepole pine, subalpine fir-Engelmann spruce, white spruce, and balsam poplar communities on the Jumping Creek Watershed, landform was the primary driver of fire frequencies. Using time-since-fire distributions, the authors found that riparian zones and the larger, entire-watershed area had similar mean fire frequencies. At a fine scale, however, fires were less frequent near stream channels with gravel- or sandbars, whereas terraces above straight streams without bars tended to burn at same frequency as the overall watershed. From 1851 to 1890, fire-return intervals averaged 71 years in riparian and 48 years in entire-watershed zones, respectively. After 1891, fire-return intervals lengthened to 208 and 178 years, respectively. The authors concluded that on the Jumping Creek Watershed, except for bars, areas adjacent to streams were just as likely to burn as upland areas [66]. Understory vegetation was not surveyed in the study, but thinleaf alder is known to occur in the area.
Two studies suggest a trend of longer fire-return interval in riparian Douglas-fir than upland Douglas-fir forests. In Douglas-fir forests on the Steamboat Creek Watershed in southwestern Oregon, mean fire-return intervals were longer in riparian than upland sites (37 vs. 31.5 years), but the difference was not significant. Fire-return intervals ranged from 4 to 167 years on riparian and 2 to 110 years on upland sites. The authors speculated that fires were historically common, of mixed severity, and patchy on their study sites, with a high likelihood of fire in both riparian and upland zones. They suggested that historically, many riparian forests burned when upslope fires backed into riparian zones [268]. On eastside riparian Douglas-fir forests in the Cascade Range of Washington, fire-scarred trees were consistently fewer in riparian than on upland sites. The authors concede that fewer fire scars does not necessarily mean fewer fires; fires in the riparian zone could have been mostly low-severity surface fires that caused less scarring than fires in upland areas, or fires could have been more severe, stand-replacement fires that left fewer standing live, fire-scarred trees than in upland zones. Plant association groups in riparian forests had 25% to 42% fewer wildfires than similar plant associations on sideslopes. Fire-return intervals ranged from 11 to 46 years in riparian forests and from 7 to 39 years on sideslopes. On sideslopes, stands on south-, east-, and west-facing slopes in riparian zones were older than those in nonriparian zones (P≤0.1); differences in stand ages between riparian and nonriparian sites on north-facing slopes were not significant [115].
On the Shasta-Trinity National Forest in the Klamath Mountains of California, fire-return intervals in riparian and upland mixed-conifer forests were compared. Intervals between fires were consistently longer in riparian vs. upland sites [310]:
| Fire-return intervals of riparian and upland mixed-conifer sites in the Klamath Mountains [310] | |||
| Site | Mean fire-return interval (range) | Time period | Number of stumps sampled |
| Root Creek, riparian |
33 years (7-56) | 1673-1880 | 4 |
| Root Creek, upland |
7 years (3-44) | 1749-1924 | 14 |
| North Fork Shotgun Creek, riparian |
16 years (5-56) | 1710-1916 | 9 |
| North Fork Shotgun Creek, upland |
8 years (4-64) | 1622-1887 | 16 |
In a fire history study of Jeffrey pine, white fir, and mixed-conifer forests of the Sierra Nevada in which thinleaf alder was "highly abundant", fire-return intervals were usually similar on riparian and upland sites. On 36 paired riparian and upland sites on the Lassen and Tahoe National Forests, only 9 paired sites had significantly different fire-return intervals (P≤0.1). Fire-return intervals ranged from 15.3 to 86.5 years on riparian sites and from 10.0 to 56.3 years on upland sites. They averaged 30.0 and 27.8 years on riparian and upland sites, respectively. Most fire scarring (88% on riparian and 79% on upland sites) happened in late summer and early fall, after tree-ring growth had stopped [340].
Fires in the spruce-birch and aspen ecosystems of interior Alaska are usually stand-replacing and often extensive [234,344], burning "for days with high intensities, covering vast acreages" [357]. Fire frequency is poorly known but probably ranges from 75 to 150 years. In quaking aspen/thinleaf alder stands on Ester Dome, fire kill was the primary cause of tree mortality [206].
Near Rainbow Lake in Wood Buffalo National Park, Alberta, a fire history study found charcoal evidence of at least 12 local fires in 840 years, with a mean return interval of 69 years. In some cases, thinleaf alder pollen counts (a surrogate for thinleaf alder density) peaked about 35 years after a fire; however, thinleaf alder pollen counts declined after 6 of the local fires. The extant plant community is mixed hardwood-conifer. Thinleaf alder is most common at the lake margins, along with scattered black spruce and tamarack [216].
See the Fire Regime Table for thinleaf alder for further information on fire regimes of vegetation communities in which thinleaf alder may occur. Find further fire regime information for the plant communities in which this species may occur by entering the species name in the FEIS home page under "Find Fire Regimes".
FIRE MANAGEMENT CONSIDERATIONS:In the Great Lakes states, frequent prescribed fires (no return interval given) can help control speckled alder, beaked hazelnut, and American hazelnut in the understories of red pine plantations. Summer fires best prepare a mineral soil seedbed necessary for natural red pine regeneration, and the generally drier fuels results in severer fires and better shrub control than spring prescribed fires [46].
On plantations, interference of postfire speckled alder regeneration with conifer regeneration and growth may be minimal on moist lowland sites where fire has prepared a mineral seedbed for conifers. Speckled alder and mountain alder "grew well" on such sites 10 years after clearcutting and 8 years after a late spring (2 June 1970) prescribed fire on a jack pine/black spruce plantation in southeastern Manitoba. Chrosciewicz [69] concluded that even though "plant competition was generally much more severe" on moist lowland sites than on dry or fresh upland sites, the alders and other postfire regeneration "had no detrimental effect on regeneration of commercial tree species, including pine" on moist lowland sites. Postfire conifer establishment was poor on the upland sites, which the author attributed to lower than normal precipitation and consequently, drier than normal soils, during the first 2 postfire growing seasons [69]. Another study by Chrosciewicz [68], near Winnipeg, Manitoba, had similar results. Five years after low- and moderate-severity spring (late May) prescribed fires, shrub cover, including speckled alder, averaged 15% to 20% in regenerating black spruce stands. This "rather luxuriant vegetation" "seemed to provide valuable shelter to the spruce seedlings in their early development" [68]. See the Research Project Summary on Chrosciewicz's [68] research for information on the fire prescriptions, fire behavior, and for details of black spruce's postfire response.
Early spring burning—after snowmelt but before soil thaw—may help control speckled alder and other woody invaders in wetlands. Researchers found that early spring prescribed burning was more cost-effective than early and late summer cutting or late manual torching for reducing woody encroachment in a tussock sedge-bluejoint grass wetland meadow in Wisconsin. Additionally, it caused the least disturbance to groundlayer vegetation and to the soil. The researchers warned that due to sprouting of woody species (which occurred after all fire and cutting treatments), prescribed fire must be viewed as an ongoing treatment strategy. Speckled alder was a component of the woody vegetation, although Sandberg's birch (Betula × sandbergii) and red-osier dogwood had the most cover. Fire effects on speckled alder alone were not measured [294].
Managers of the Lac St-Francois National Wildlife Area in Quebec found that neither a single wildfire nor mechanical treatment (double-cutting shoots) controlled speckled alder invasion on an acidic, water sedge-hairy sedge fen in the short term, while stump applications of herbicide reduced speckled alder but also reduced nontarget plants. The wildfire had burned part of the fen prior to the study, and plots were established on sections of the burn that were "highly invaded by small to medium-sized individual alder shrubs". Speckled alder root crowns were located at or below the water table. All of 40 speckled alder individuals on study plots survived the fire and were sprouting "vigorously" in postfire year 1. For the double-cut treatment, stems were cut to the root crown in mid-June, with a 2nd cutting of new sprouts in early August. The next year, 80% of cut plants were sprouting "vigorously". While conceding that another cut would increase speckled alder mortality, the authors found cutting too ineffective and costly to justify further cuttings. Foliar applications of glyphosate in July gave 100% control of speckled alder but also killed more nontarget plants than considered acceptable (no limits given). For cut-stump glyphosate treatments, either August or March application gave good control (79% and 93%, respectively). The March treatment was preferred, because it was more effective and accessibility was greater on the frozen winter ground than on the wet summer ground [38]. The authors noted that based on other research [46,161], very frequent prescribed fires (1 or 2 fires annually over 5 years) may control speckled alder but would "carry very high operation costs" and possibly be detrimental to the fen. However, they considered prescribed fire a viable option for long-term control of speckled alder [38].
Thinleaf alder: Recommendations for prescribed fire use in thinleaf alder communities are few and sometimes contradictory. Prescribed fire in these communities may increase plant community productivity or conversely, be used as a tool to control thinleaf alder. Olason and Agee [268] note that although fires were probably historically frequent in riparian Douglas-fir forests of the Pacific Northwest, it may be necessary to "totally protect" riparian forests in areas where favorable salmonid habitat is limited. They suggest treating upslope forests with prescribed fire or fire-surrogate treatments, speculating that such treatments would make wildfires less likely to burn into moist portions of riparian zones [268].
Others suggest that excluding fire from riparian zones may be harmful in the long term. On the Payette National Forest, Idaho, a low-severity May prescribed fire in an upland subalpine fir-Engelmann spruce community burned only slightly into an alder (Alnus spp.)-dominated riparian zone. Comparing these results to late summer wildfires in the same communities, the authors found that with wildfires, the area burned in riparian communities was proportionate to that burned in the upland communities. While fire severity was generally lower in the riparian than the upland communities, portions of the riparian zones burned at high severity. The authors concluded that in riparian zones, spring prescribed fire is not an ecological surrogate for fall wildfire and that a "prescribed fire regime" of repeatedly burning upland communities while excluding fire from riparian forests may eliminate important natural disturbances from riparian and stream habitats [8]. Besides wildfire itself, these disturbances may include postfire erosion, altered stream channels, altered stream temperatures, and altered species composition of the aquatic animal communities [8,107].
Effects of prescribed fires in riparian and wetland zones may be difficult to predict. Prescribed fire may help control thinleaf alder and other woody invasives in freshwater wetlands [42]. Jacobson and others' study [182] provides management recommendations for reducing woody encroachment on open fens. However, in forests, thinleaf alder may increase after overstory removal by prescribed fire and/or cutting, especially on wet sites. Researchers in British Columbia report that although moderate to severe fires are needed to "set back the growth" of thinleaf alder, such fires rarely carry in the wet to moist sites where thinleaf alder usually grows [71].
Thinleaf alder-dominated communities may recover more quickly than adjacent upland communities. Six years after the Diamond Peak Wildfire Complex on the Payette National Forest, woody plant cover in thinleaf alder-dominated riparian communities was greatest on sites where fire severity was greatest (P=0.05). Across 6 riparian sites burned at varying severities, total woody cover ranged from a low of 8% in postfire year 2 to 30% in postfire year 6. The authors found that in postfire year 6, "little recovery had taken place in the upland (ponderosa pine) community" [287].
A May wildfire in a quaking aspen-balsam poplar/thinleaf alder-red alder community reduced nesting and hiding cover for ruffed grouse in the short term. Alders (thinleaf and red alder combined) averaged 8.0 feet (2.4 m) tall 1 year before and 3.5 feet (1.1 m) tall 1 year after the fire. Along a 2,000-foot (600 m) transect, alders <3 feet (2 m) tall averaged 35.8 stems/transect before fire and 3.8 stems/transect 1 year after fire. On drumming sites used by male ruffed grouse, alders <3 feet tall averaged 28.7 stems/transect before fire and 2.9 stems/transect after fire. In postfire years 1 and 2, estimated ruffed grouse numbers in burned areas averaged 50% of ruffed grouse numbers 1 year before fire. By postfire year 3, density of males in unburned drumming areas was almost 3 times that of males in burned drumming areas [99]. See the FEIS review of ruffed grouse for details.White-tailed deer and lagomorphs browse speckled alder [176]. It provides winter forage (review by [141]), but browsing wildlife are likely to avoid speckled alder if more palatable species are available. In a winter deer yard in Quebec, white-tailed deer browsed red-osier dogwood and pussy willow more than expected, and speckled alder less than expected, based on availability [41].
In cafeteria feeding trials in Wisconsin, captive ruffed grouse avoided speckled alder catkins, preferring those of quaking aspen and willows. The authors speculated that secondary metabolites in speckled alder catkins rendered the speckled alder catkins unpalatable [140].
Mule deer browse thinleaf alder sparingly [261,348], although thinleaf alder may be more important in winter [261]. Mountain goats also use thinleaf alder [240], and livestock may browse thinleaf alder heavily in riparian areas [256]
Arthropods and mollusks consume gray alder. Millipedes browse fresh speckled alder leaf litter [235]. Landsnails (Cepaea nemoralis) also browse speckled alder leaf litter [360]. Speckled alder is a host of alder aphids (Prociphilus), which feed only on alders (Alnus) [368]. Alder aphids are a larval food of the harvester butterfly (Feniseca tarquinius) [305].
Palatability and/or nutritional value: Gray alder's palatability is rated poor to fair for big game animals. Sprouts and young plants are preferred over older browse ([334], review by [141]). Its palatability for livestock is rated from poor in the Southwest to fair in the Pacific Northwest and California. Gray alder is generally more palatable to cattle than to domestic sheep and goats [334].
Songbirds, grouse, and American woodcocks eat gray alder buds, seeds, and catkins (reviews by [142,160]). See Grodzinski and Sawicka-Kapusta [139] for information on the nutritional value of gray alder seeds.
Speckled alder is not a preferred browse taxon; it is used primarily as winter forage. It was ranked very low in palatability among woody taxa (22nd among 23) that white-tailed deer used as forage [265]. On the Apostle Islands of Wisconsin, speckled alder was 1 of 4 taxa that white-tailed deer chose last as browse; 28 taxa were tested [21]. A habitat suitability model, developed for coniferous and aspen-birch ecosystems of the Great Lakes region, lists speckled alder as low-preference browse for moose during the growing season [5]. A study on Isle Royal, Michigan, found moose used speckled alder less than expected based on speckled alder's availability and nutritional content (P≤0.05) [25]. American beavers, however, used speckled alder as much as expected (P<0.05) [26].
Speckled alder is high in protein. See these sources: [25,235] for nutritional information on speckled alder browse. See this source: [140] for nutritional information on speckled alder catkins.
Thinleaf alder is generally unpalatable to wild ungulates [151,223], cattle, and horses. It is fairly palatable to domestic sheep [32]. Wild ungulates browse new thinleaf alder growth but generally avoid mature thinleaf alder [151] except as emergency winter forage. On summer rangelands in Tehama County, California, mule deer browsed thinleaf alder less than expected based on availability [223].
In Alaska, feltleaf willow and balsam poplar were more palatable to browsing mammals when grown in the shade of thinleaf alder than when grown in the open (Rohleder 1985 cited in [45]).
Cover value: Gray alder provides shade and hiding cover for many vertebrate species.
Speckled alder: In a Quebec study, speckled alder was the dominant streamside shrub in an ecosystem supporting brook trout, American mink, and Appalachian brook crayfish (Cambarus bartoni), the American mink's primary prey species in that stream [50]. The federally endangered bog turtle [336] is positively associated with speckled alder wetlands (review by [50]). In Alberta, white spruce/gray alder/horsetail/moss (Equisetum/Hypnum and Polytichum spp.) riparian communities provide shade and reduce water temperatures in trout streams [77].
Speckled alder is critical for American beavers. They use speckled alder for den and dam construction more than they use it as browse [103,222]. In Algonquin Provincial Park, Ontario, the number of speckled alder stems found in American beaver dams (403 stems) was over twice that of speckled alder stems found in their food caches (176 stems). Speckled alder was selected for dam construction about 4 times as often as spruces, which were the next most preferred [103]. In another Algonquin Provincial Park study, American beavers used speckled alder more than expected. Although speckled alder composed 25.6% of the standing vegetation, 49.2% of stems that American beavers felled were speckled alder. Speckled alder cover and basal area increased significantly with distance from American beaver dens (P<0.009) [101,102].
Speckled alder provides important cover for American woodcocks, ruffed grouse [99,266], and other birds. Speckled alder thickets provide critical feeding, nesting, molting, and premigratory staging habitat for American woodcocks ([252,258,266,291], Sheldon 1967 cited in [49]). In Michigan, ruffed grouse used speckled alder for cover in drumming sites (review by [160]). A study in eastern hemlock-red maple/speckled alder-mountain-laurel riparian corridors of Pennsylvania found the number of obligate wetland bird species, such as kinglets and flycatchers, was significantly higher in undisturbed speckled alder riparian corridors than in speckled alder-dominated riparian corridors disturbed by residential or agricultural development [83]. Niemi and Pfannmuller [266] provide a list of bird species using speckled alder-dominated wetlands as breeding and nesting habitat in Minnesota and Michigan.
Thinleaf alder: Thinleaf alder provides cover for snowshoe hares [353] and big game animals [261]. Mule deer, white-tailed deer, and elk use thinleaf alder communities for thermal and hiding cover [151]. American beavers use thinleaf alder to make their dens (review by [160].
Thinleaf alder provides forage and nesting sites for songbirds, upland game birds, and wetland birds (reviews by [143,149]).
Thinleaf alder provides cover for aquatic organisms. Dense thinleaf alder thickets provide thermal and shade cover for fish. Thinleaf alder and associated shrubs stabilize stream channels, and overhanging banks reduce erosion [32,151]. In Oregon, thinleaf alder on streambanks provide cover, food, and shade for salmonids [200]. In the Trout Creek Mountains, thinleaf alder-Wood's rose overstories shaded over 70% of the stream channels of 4 streams, while the more open thinleaf alder/woolly sedge communities shaded about 50% of the stream channels. Dense woolly sedge that grew over the streambank enhanced trout habitat [114].
Cattle loaf in thinleaf alder stands, although dense thinleaf alder thickets may impair access. Heavy cattle trampling may degrade thinleaf alder habitats by creating dish-shaped stream channels and reducing cover of palatable herbs [32,151].
VALUE FOR REHABILITATION OF DISTURBED SITES:Speckled alder: See these sources: [154,160] for propagation information.
Thinleaf alder: Thinleaf alder is used in revegetation projects [62,256]. It is recommended for riparian revegetation in the Intermountain West [59] and the Northern and Southern Rocky Mountains [58]. It is particularly recommended for high-gradient, high-velocity, low- to midorder streams [59]. See these sources: [104,154,183,256,309] for propagation information.
OTHER USES:American Indians used gray alder medicinally (Moerman 1986 cited in [120]). They used the bark to make red dye [157,190,203,215,348].
OTHER MANAGEMENT CONSIDERATIONS:When using mechanical treatment to control gray alder, the root crown must be destroyed to prevent sprouting. Stems in contact with soil may layer; layering is common after logging [51]. These sources: [38,51] provide information on controlling speckled alder with herbicides.
Although it is not heavily browsed when other forage is available, livestock concentrated in riparian areas sometimes overbrowse and trample thinleaf alder. In northeastern Oregon, thinleaf alder density was significantly greater on ungrazed gravelbar communities than on riparian sites with livestock (P<0.01) [138]. In extreme cases, thinleaf alder becomes scattered and short, with mostly broken branches, and streambanks of thinleaf alder communities degrade to dish shapes. In central Oregon, it took at least 5 years for thinleaf alder to recover from such overbrowsing, and at least 2 to 5 years for streams to reform banked channels, which had become dish-shaped with trampling [200]. Ehrhart and Hansen [113] provide management guidelines for grazing cattle in riparian zones.| Fire regime information on vegetation communities in which speckled alder may occur. This information is taken from the LANDFIRE Rapid Assessment Vegetation Models [214], which were developed by local experts using available literature, local data, and/or expert opinion. This table summarizes fire regime characteristics for each plant community listed. The PDF file linked from each plant community name describes the model and synthesizes the knowledge available on vegetation composition, structure, and dynamics in that community. Cells are blank where information is not available in the Rapid Assessment Vegetation Model. | |||||||||
|
|||||||||
| Great Lakes | |||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||
| Great Lakes Woodland | |||||||||
| Great Lakes pine barrens | Replacement | 8% | 41 | 10 | 80 | ||||
| Mixed | 9% | 36 | 10 | 80 | |||||
| Surface or low | 83% | 4 | 1 | 20 | |||||
| Jack pine-open lands (frequent fire-return interval) | Replacement | 83% | 26 | 10 | 100 | ||||
| Mixed | 17% | 125 | 10 | ||||||
| Northern oak savanna | Replacement | 4% | 110 | 50 | 500 | ||||
| Mixed | 9% | 50 | 15 | 150 | |||||
| Surface or low | 87% | 5 | 1 | 20 | |||||
| Great Lakes Forested | |||||||||
| Northern hardwood maple-beech-eastern hemlock | Replacement | 60% | >1,000 | ||||||
| Mixed | 40% | >1,000 | |||||||
| Conifer lowland (embedded in fire-prone ecosystem) | Replacement | 45% | 120 | 90 | 220 | ||||
| Mixed | 55% | 100 | |||||||
| Conifer lowland (embedded in fire-resistant ecosystem) | Replacement | 36% | 540 | 220 | >1,000 | ||||
| Mixed | 64% | 300 | |||||||
| Great Lakes floodplain forest | Mixed | 7% | 833 | ||||||
| Surface or low | 93% | 61 | |||||||
| Great Lakes spruce-fir | Replacement | 100% | 85 | 50 | 200 | ||||
| Maple-basswood | Replacement | 33% | >1,000 | ||||||
| Surface or low | 67% | 500 | |||||||
| Maple-basswood mesic hardwood forest (Great Lakes) | Replacement | 100% | >1,000 | >1,000 | >1,000 | ||||
| Maple-basswood-oak-aspen | Replacement | 4% | 769 | ||||||
| Mixed | 7% | 476 | |||||||
| Surface or low | 89% | 35 | |||||||
| Northern hardwood-eastern hemlock forest (Great Lakes) | Replacement | 99% | >1,000 | ||||||
| Oak-hickory | Replacement | 13% | 66 | 1 | |||||
| Mixed | 11% | 77 | 5 | ||||||
| Surface or low | 76% | 11 | 2 | 25 | |||||
| Pine-oak | Replacement | 19% | 357 | ||||||
| Surface or low | 81% | 85 | |||||||
| Red pine-eastern white pine (frequent fire) | Replacement | 38% | 56 | ||||||
| Mixed | 36% | 60 | |||||||
| Surface or low | 26% | 84 | |||||||
| Red pine-eastern white pine (less frequent fire) | Replacement | 30% | 166 | ||||||
| Mixed | 47% | 105 | |||||||
| Surface or low | 23% | 220 | |||||||
| Great Lakes pine forest, eastern white pine-eastern hemlock (frequent fire) | Replacement | 52% | 260 | ||||||
| Mixed | 12% | >1,000 | |||||||
| Surface or low | 35% | 385 | |||||||
| Eastern white pine-eastern hemlock | Replacement | 54% | 370 | ||||||
| Mixed | 12% | >1,000 | |||||||
| Surface or low | 34% | 588 | |||||||
| Northeast | |||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||
| Northeast Woodland | |||||||||
| Eastern woodland mosaic | Replacement | 2% | 200 | 100 | 300 | ||||
| Mixed | 9% | 40 | 20 | 60 | |||||
| Surface or low | 89% | 4 | 1 | 7 | |||||
| Rocky outcrop pine (Northeast) | Replacement | 16% | 128 | ||||||
| Mixed | 32% | 65 | |||||||
| Surface or low | 52% | 40 | |||||||
| Pine barrens | Replacement | 10% | 78 | ||||||
| Mixed | 25% | 32 | |||||||
| Surface or low | 65% | 12 | |||||||
| Oak-pine (eastern dry-xeric) | Replacement | 4% | 185 | ||||||
| Mixed | 7% | 110 | |||||||
| Surface or low | 90% | 8 | |||||||
| Northeast Forested | |||||||||
| Northern hardwoods (Northeast) | Replacement | 39% | >1,000 | ||||||
| Mixed | 61% | 650 | |||||||
| Eastern white pine-northern hardwood | Replacement | 72% | 475 | ||||||
| Surface or low | 28% | >1,000 | |||||||
| Northern hardwoods-eastern hemlock | Replacement | 50% | >1,000 | ||||||
| Surface or low | 50% | >1,000 | |||||||
| Northern hardwoods-spruce | Replacement | 100% | >1,000 | 400 | >1,000 | ||||
| Appalachian oak forest (dry-mesic) | Replacement | 2% | 625 | 500 | >1,000 | ||||
| Mixed | 6% | 250 | 200 | 500 | |||||
| Surface or low | 92% | 15 | 7 | 26 | |||||
| Beech-maple | Replacement | 100% | >1,000 | ||||||
| Northeast spruce-fir forest | Replacement | 100% | 265 | 150 | 300 | ||||
| Southeastern red spruce-Fraser fir | Replacement | 100% | 500 | 300 | >1,000 | ||||
| Southern Appalachians | |||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||
| Southern Appalachians Woodland | |||||||||
| Appalachian shortleaf pine | Replacement | 4% | 125 | ||||||
| Mixed | 4% | 155 | |||||||
| Surface or low | 92% | 6 | |||||||
| Table Mountain-pitch pine | Replacement | 5% | 100 | ||||||
| Mixed | 3% | 160 | |||||||
| Surface or low | 92% | 5 | |||||||
| Oak-ash woodland | Replacement | 23% | 119 | ||||||
| Mixed | 28% | 95 | |||||||
| Surface or low | 49% | 55 | |||||||
| Southern Appalachians Forested | |||||||||
| Bottomland hardwood forest | Replacement | 25% | 435 | 200 | >1,000 | ||||
| Mixed | 24% | 455 | 150 | 500 | |||||
| Surface or low | 51% | 210 | 50 | 250 | |||||
| Mixed mesophytic hardwood | Replacement | 11% | 665 | ||||||
| Mixed | 10% | 715 | |||||||
| Surface or low | 79% | 90 | |||||||
| Appalachian oak-hickory-pine | Replacement | 3% | 180 | 30 | 500 | ||||
| Mixed | 8% | 65 | 15 | 150 | |||||
| Surface or low | 89% | 6 | 3 | 10 | |||||
| Eastern hemlock-eastern white pine-hardwood | Replacement | 17% | >1,000 | 500 | >1,000 | ||||
| Surface or low | 83% | 210 | 100 | >1,000 | |||||
| Red pine-eastern white pine (frequent fire) | Replacement | 38% | 56 | ||||||
| Mixed | 36% | 60 | |||||||
| Surface or low | 26% | 84 | |||||||
| Eastern white pine-northern hardwood | Replacement | 72% | 475 | ||||||
| Surface or low | 28% | >1,000 | |||||||
| Appalachian Virginia pine | Replacement | 20% | 110 | 25 | 125 | ||||
| Mixed | 15% | 145 | |||||||
| Surface or low | 64% | 35 | 10 | 40 | |||||
| Appalachian oak forest (dry-mesic) | Replacement | 6% | 220 | ||||||
| Mixed | 15% | 90 | |||||||
| Surface or low | 79% | 17 | |||||||
| Southern Appalachian high-elevation forest | Replacement | 59% | 525 | ||||||
| Mixed | 41% | 770 | |||||||
| Southeast | |||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||
| Southeast Shrubland | |||||||||
| Pocosin | Replacement | 1% | >1,000 | 30 | >1,000 | ||||
| Mixed | 99% | 12 | 3 | 20 | |||||
| Southeast Woodland | |||||||||
| Longleaf pine/bluestem | Replacement | 3% | 130 | ||||||
| Surface or low | 97% | 4 | 1 | 5 | |||||
| Longleaf pine (mesic uplands) | Replacement | 3% | 110 | 40 | 200 | ||||
| Surface or low | 97% | 3 | 1 | 5 | |||||
| Longleaf pine-Sandhills prairie | Replacement | 3% | 130 | 25 | 500 | ||||
| Surface or low | 97% | 4 | 1 | 10 | |||||
| Pine rocklands | Mixed | 1% | 330 | ||||||
| Surface or low | 99% | 3 | 1 | 5 | |||||
| Pond pine | Replacement | 64% | 7 | 5 | 500 | ||||
| Mixed | 25% | 18 | 8 | 150 | |||||
| Surface or low | 10% | 43 | 2 | 50 | |||||
| Atlantic wet pine savanna | Replacement | 4% | 100 | ||||||
| Mixed | 2% | 175 | |||||||
| Surface or low | 94% | 4 | |||||||
| Southeast Forested | |||||||||
| Sand pine scrub | Replacement | 90% | 45 | 10 | 100 | ||||
| Mixed | 10% | 400 | 60 | ||||||
| Coastal Plain pine-oak-hickory | Replacement | 4% | 200 | ||||||
| Mixed | 7% | 100 | |||||||
| Surface or low | 89% | 8 | |||||||
| Atlantic white-cedar forest | Replacement | 34% | 200 | 25 | 350 | ||||
| Mixed | 8% | 900 | 20 | 900 | |||||
| Surface or low | 59% | 115 | 10 | 500 | |||||
| Maritime forest | Replacement | 18% | 40 | 500 | |||||
| Mixed | 2% | 310 | 100 | 500 | |||||
| Surface or low | 80% | 9 | 3 | 50 | |||||
| Loess bluff and plain forest | Replacement | 7% | 476 | ||||||
| Mixed | 9% | 385 | |||||||
| Surface or low | 85% | 39 | |||||||
| Southern floodplain | Replacement | 7% | 900 | ||||||
| Surface or low | 93% | 63 | |||||||
| *Fire Severities— Replacement: Any fire that causes greater than 75% top removal of a vegetation-fuel type, resulting in general replacement of existing vegetation; may or may not cause a lethal effect on the plants. Mixed: Any fire burning more than 5% of an area that does not qualify as a replacement, surface, or low-severity fire; includes mosaic and other fires that are intermediate in effects. Surface or low: Any fire that causes less than 25% upper layer replacement and/or removal in a vegetation-fuel class but burns 5% or more of the area [148,213]. |
|||||||||
| Fire regime information on vegetation communities in which thinleaf alder may occur. This information is taken from the LANDFIRE Rapid Assessment Vegetation Models [214], which were developed by local experts using available literature, local data, and/or expert opinion. This table summarizes fire regime characteristics for each plant community listed. The PDF file linked from each plant community name describes the model and synthesizes the knowledge available on vegetation composition, structure, and dynamics in that community. Cells are blank where information is not available in the Rapid Assessment Vegetation Model. | |||||||||||||
|
|||||||||||||
| Pacific Northwest | |||||||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||||||
| Northwest Grassland | |||||||||||||
| Marsh | Replacement | 74% | 7 | ||||||||||
| Mixed | 26% | 20 | |||||||||||
| Bluebunch wheatgrass | Replacement | 47% | 18 | 5 | 20 | ||||||||
| Mixed | 53% | 16 | 5 | 20 | |||||||||
| Idaho fescue grasslands | Replacement | 76% | 40 | ||||||||||
| Mixed | 24% | 125 | |||||||||||
| Alpine and subalpine meadows and grasslands | Replacement | 68% | 350 | 200 | 500 | ||||||||
| Mixed | 32% | 750 | 500 | >1,000 | |||||||||
| Northwest Shrubland | |||||||||||||
| Wyoming big sagebrush semidesert | Replacement | 86% | 200 | 30 | 200 | ||||||||
| Mixed | 9% | >1,000 | 20 | ||||||||||
| Surface or low | 5% | >1,000 | 20 | ||||||||||
| Wyoming sagebrush steppe | Replacement | 89% | 92 | 30 | 120 | ||||||||
| Mixed | 11% | 714 | 120 | ||||||||||
| Mountain big sagebrush (cool sagebrush) | Replacement | 100% | 20 | 10 | 40 | ||||||||
| Northwest Woodland | |||||||||||||
| Western juniper (pumice) | Replacement | 33% | >1,000 | ||||||||||
| Mixed | 67% | 500 | |||||||||||
| Oregon white oak-ponderosa pine | Replacement | 16% | 125 | 100 | 300 | ||||||||
| Mixed | 2% | 900 | 50 | ||||||||||
| Surface or low | 81% | 25 | 5 | 30 | |||||||||
| Pine savannah (ultramafic) | Replacement | 7% | 200 | 100 | 300 | ||||||||
| Surface or low | 93% | 15 | 10 | 20 | |||||||||
| Ponderosa pine | Replacement | 5% | 200 | ||||||||||
| Mixed | 17% | 60 | |||||||||||
| Surface or low | 78% | 13 | |||||||||||
| Oregon white oak | Replacement | 3% | 275 | ||||||||||
| Mixed | 19% | 50 | |||||||||||
| Surface or low | 78% | 12.5 | |||||||||||
| Subalpine woodland | Replacement | 21% | 300 | 200 | 400 | ||||||||
| Mixed | 79% | 80 | 35 | 120 | |||||||||
| Northwest Forested | |||||||||||||
| Sitka spruce-western hemlock | Replacement | 100% | 700 | 300 | >1,000 | ||||||||
| Douglas-fir (Willamette Valley foothills) | Replacement | 18% | 150 | 100 | 400 | ||||||||
| Mixed | 29% | 90 | 40 | 150 | |||||||||
| Surface or low | 53% | 50 | 20 | 80 | |||||||||
| Oregon coastal tanoak | Replacement | 10% | 250 | ||||||||||
| Mixed | 90% | 28 | 15 | 40 | |||||||||
| Dry ponderosa pine (mesic) | Replacement | 5% | 125 | ||||||||||
| Mixed | 13% | 50 | |||||||||||
| Surface or low | 82% | 8 | |||||||||||
| Douglas-fir-western hemlock (dry mesic) | Replacement | 25% | 300 | 250 | 500 | ||||||||
| Mixed | 75% | 100 | 50 | 150 | |||||||||
| Douglas-fir-western hemlock (wet mesic) | Replacement | 71% | 400 | ||||||||||
| Mixed | 29% | >1,000 | |||||||||||
| Mixed conifer (southwestern Oregon) | Replacement | 4% | 400 | ||||||||||
| Mixed | 29% | 50 | |||||||||||
| Surface or low | 67% | 22 | |||||||||||
| California mixed evergreen (northern California and southern Oregon) | Replacement | 6% | 150 | 100 | 200 | ||||||||
| Mixed | 29% | 33 | 15 | 50 | |||||||||
| Surface or low | 64% | 15 | 5 | 30 | |||||||||
| Mountain hemlock | Replacement | 93% | 750 | 500 | >1,000 | ||||||||
| Mixed | 7% | >1,000 | |||||||||||
| Lodgepole pine (pumice soils) | Replacement | 78% | 125 | 65 | 200 | ||||||||
| Mixed | 22% | 450 | 45 | 85 | |||||||||
| Pacific silver fir (low elevation) | Replacement | 46% | 350 | 100 | 800 | ||||||||
| Mixed | 54% | 300 | 100 | 400 | |||||||||
| Pacific silver fir (high elevation) | Replacement | 69% | 500 | ||||||||||
| Mixed | 31% | >1,000 | |||||||||||
| Subalpine fir | Replacement | 81% | 185 | 150 | 300 | ||||||||
| Mixed | 19% | 800 | 500 | >1,000 | |||||||||
| Mixed conifer (eastside dry) | Replacement | 14% | 115 | 70 | 200 | ||||||||
| Mixed | 21% | 75 | 70 | 175 | |||||||||
| Surface or low | 64% | 25 | 20 | 25 | |||||||||
| Mixed conifer (eastside mesic) | Replacement | 35% | 200 | ||||||||||
| Mixed | 47% | 150 | |||||||||||
| Surface or low | 18% | 400 | |||||||||||
| Red fir | Replacement | 20% | 400 | 150 | 400 | ||||||||
| Mixed | 80% | 100 | 80 | 130 | |||||||||
| Spruce-fir | Replacement | 84% | 135 | 80 | 270 | ||||||||
| Mixed | 16% | 700 | 285 | >1,000 | |||||||||
| California | |||||||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||||||
| California Grassland | |||||||||||||
| California grassland | Replacement | 100% | 2 | 1 | 3 | ||||||||
| Herbaceous wetland | Replacement | 70% | 15 | ||||||||||
| Mixed | 30% | 35 | |||||||||||
| Wet mountain meadow-Lodgepole pine (subalpine) | Replacement | 21% | 100 | ||||||||||
| Mixed | 10% | 200 | |||||||||||
| Surface or low | 69% | 30 | |||||||||||
| Alpine meadows barrens | Replacement | 100% | 200 | 200 | 400 | ||||||||
| California Shrubland | |||||||||||||
| Coastal sage scrub | Replacement | 100% | 50 | 20 | 150 | ||||||||
| Coastal sage scrub-coastal prairie | Replacement | 8% | 40 | 8 | 900 | ||||||||
| Mixed | 31% | 10 | 1 | 900 | |||||||||
| Surface or low | 62% | 5 | 1 | 6 | |||||||||
| Chaparral | Replacement | 100% | 50 | 30 | 125 | ||||||||
| Montane chaparral | Replacement | 34% | 95 | ||||||||||
| Mixed | 66% | 50 | |||||||||||
| California Woodland | |||||||||||||
| California oak woodlands | Replacement | 8% | 120 | ||||||||||
| Mixed | 2% | 500 | |||||||||||
| Surface or low | 91% | 10 | |||||||||||
| Ponderosa pine | Replacement | 5% | 200 | ||||||||||
| Mixed | 17% | 60 | |||||||||||
| Surface or low | 78% | 13 | |||||||||||
| California Forested | |||||||||||||
| California mixed evergreen | Replacement | 10% | 140 | 65 | 700 | ||||||||
| Mixed | 58% | 25 | 10 | 33 | |||||||||
| Surface or low | 32% | 45 | 7 | ||||||||||
| Coast redwood | Replacement | 2% | ≥1,000 | ||||||||||
| Surface or low | 98% | 20 | |||||||||||
| Mixed conifer (north slopes) | Replacement | 5% | 250 | ||||||||||
| Mixed | 7% | 200 | |||||||||||
| Surface or low | 88% | 15 | 10 | 40 | |||||||||
| Mixed conifer (south slopes) | Replacement | 4% | 200 | ||||||||||
| Mixed | 16% | 50 | |||||||||||
| Surface or low | 80% | 10 | |||||||||||
| Aspen with conifer | Replacement | 24% | 155 | 50 | 300 | ||||||||
| Mixed | 15% | 240 | |||||||||||
| Surface or low | 61% | 60 | |||||||||||
| Jeffrey pine | Replacement | 9% | 250 | ||||||||||
| Mixed | 17% | 130 | |||||||||||
| Surface or low | 74% | 30 | |||||||||||
| Mixed evergreen-bigcone Douglas-fir (southern coastal) | Replacement | 29% | 250 | ||||||||||
| Mixed | 71% | 100 | |||||||||||
| Interior white fir (northeastern California) | Replacement | 47% | 145 | ||||||||||
| Mixed | 32% | 210 | |||||||||||
| Surface or low | 21% | 325 | |||||||||||
| Red fir-white fir | Replacement | 13% | 200 | 125 | 500 | ||||||||
| Mixed | 36% | 70 | |||||||||||
| Surface or low | 51% | 50 | 15 | 50 | |||||||||
| Red fir-western white pine | Replacement | 16% | 250 | ||||||||||
| Mixed | 65% | 60 | 25 | 80 | |||||||||
| Surface or low | 19% | 200 | |||||||||||
| Sierra Nevada lodgepole pine (cold wet upper montane) | Replacement | 23% | 150 | 37 | 764 | ||||||||
| Mixed | 70% | 50 | |||||||||||
| Surface or low | 7% | 500 | |||||||||||
| Sierra Nevada lodgepole pine (dry subalpine) | Replacement | 11% | 250 | 31 | 500 | ||||||||
| Mixed | 45% | 60 | 31 | 350 | |||||||||
| Surface or low | 45% | 60 | 9 | 350 | |||||||||
| Southwest | |||||||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||||||
| Southwest Shrubland | |||||||||||||
| Southwestern shrub steppe | Replacement | 72% | 14 | 8 | 15 | ||||||||
| Mixed | 13% | 75 | 70 | 80 | |||||||||
| Surface or low | 15% | 69 | 60 | 100 | |||||||||
| Southwestern shrub steppe with trees | Replacement | 52% | 17 | 10 | 25 | ||||||||
| Mixed | 22% | 40 | 25 | 50 | |||||||||
| Surface or low | 25% | 35 | 25 | 100 | |||||||||
| Mountain sagebrush (cool sage) | Replacement | 75% | 100 | ||||||||||
| Mixed | 25% | 300 | |||||||||||
| Southwest Woodland | |||||||||||||
| Pinyon-juniper (mixed fire regime) | Replacement | 29% | 430 | ||||||||||
| Mixed | 65% | 192 | |||||||||||
| Surface or low | 6% | >1,000 | |||||||||||
| Pinyon-juniper (rare replacement fire regime) | Replacement | 76% | 526 | ||||||||||
| Mixed | 20% | >1,000 | |||||||||||
| Surface or low | 4% | >1,000 | |||||||||||
| Ponderosa pine/grassland (Southwest) | Replacement | 3% | 300 | ||||||||||
| Surface or low | 97% | 10 | |||||||||||
| Bristlecone-limber pine (Southwest) | Replacement | 67% | 500 | ||||||||||
| Surface or low | 33% | >1,000 | |||||||||||
| Southwest Forested | |||||||||||||
| Riparian forest with conifers | Replacement | 100% | 435 | 300 | 550 | ||||||||
| Riparian deciduous woodland | Replacement | 50% | 110 | 15 | 200 | ||||||||
| Mixed | 20% | 275 | 25 | ||||||||||
| Surface or low | 30% | 180 | 10 | ||||||||||
| Ponderosa pine-Gambel oak (southern Rockies and Southwest) | Replacement | 8% | 300 | ||||||||||
| Surface or low | 92% | 25 | 10 | 30 | |||||||||
| Ponderosa pine-Douglas-fir (southern Rockies) | Replacement | 15% | 460 | ||||||||||
| Mixed | 43% | 160 | |||||||||||
| Surface or low | 43% | 160 | |||||||||||
| Southwest mixed conifer (warm, dry with aspen) | Replacement | 7% | 300 | ||||||||||
| Mixed | 13% | 150 | 80 | 200 | |||||||||
| Surface or low | 80% | 25 | 2 | 70 | |||||||||
| Southwest mixed conifer (cool, moist with aspen) | Replacement | 29% | 200 | 80 | 200 | ||||||||
| Mixed | 35% | 165 | 35 | ||||||||||
| Surface or low | 36% | 160 | 10 | ||||||||||
| Aspen with spruce-fir | Replacement | 38% | 75 | 40 | 90 | ||||||||
| Mixed | 38% | 75 | 40 | ||||||||||
| Surface or low | 23% | 125 | 30 | 250 | |||||||||
| Stable aspen without conifers | Replacement | 81% | 150 | 50 | 300 | ||||||||
| Surface or low | 19% | 650 | 600 | >1,000 | |||||||||
| Lodgepole pine (Central Rocky Mountains, infrequent fire) | Replacement | 82% | 300 | 250 | 500 | ||||||||
| Surface or low | 18% | >1,000 | >1,000 | >1,000 | |||||||||
| Spruce-fir | Replacement | 96% | 210 | 150 | |||||||||
| Mixed | 4% | >1,000 | 35 | >1,000 | |||||||||
| Great Basin | |||||||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||||||
| Great Basin Grassland | |||||||||||||
| Mountain meadow (mesic to dry) | Replacement | 66% | 31 | 15 | 45 | ||||||||
| Mixed | 34% | 59 | 30 | 90 | |||||||||
| Great Basin Shrubland | |||||||||||||
| Wyoming big sagebrush semidesert | Replacement | 86% | 200 | 30 | 200 | ||||||||
| Mixed | 9% | >1,000 | 20 | >1,000 | |||||||||
| Surface or low | 5% | >1,000 | 20 | >1,000 | |||||||||
| Wyoming big sagebrush semidesert with trees | Replacement | 84% | 137 | 30 | 200 | ||||||||
| Mixed | 11% | >1,000 | 20 | >1,000 | |||||||||
| Surface or low | 5% | >1,000 | 20 | >1,000 | |||||||||
| Wyoming sagebrush steppe | Replacement | 89% | 92 | 30 | 120 | ||||||||
| Mixed | 11% | 714 | 120 | ||||||||||
| Mountain big sagebrush | Replacement | 100% | 48 | 15 | 100 | ||||||||
| Mountain big sagebrush with conifers | Replacement | 100% | 49 | 15 | 100 | ||||||||
| Mountain sagebrush (cool sage) | Replacement | 75% | 100 | ||||||||||
| Mixed | 25% | 300 | |||||||||||
| Montane chaparral | Replacement | 37% | 93 | ||||||||||
| Mixed | 63% | 54 | |||||||||||
| Gambel oak | Replacement | 75% | 50 | ||||||||||
| Mixed | 25% | 150 | |||||||||||
| Mountain shrubland with trees | Replacement | 22% | 105 | 100 | 200 | ||||||||
| Mixed | 78% | 29 | 25 | 100 | |||||||||
| Black and low sagebrushes | Replacement | 33% | 243 | 100 | |||||||||
| Mixed | 67% | 119 | 75 | 140 | |||||||||
| Black and low sagebrushes with trees | Replacement | 37% | 227 | 150 | 290 | ||||||||
| Mixed | 63% | 136 | 50 | 190 | |||||||||
| Curlleaf mountain-mahogany | Replacement | 31% | 250 | 100 | 500 | ||||||||
| Mixed | 37% | 212 | 50 | ||||||||||
| Surface or low | 31% | 250 | 50 | ||||||||||
| Great Basin Woodland | |||||||||||||
| Juniper and pinyon-juniper steppe woodland | Replacement | 20% | 333 | 100 | >1,000 | ||||||||
| Mixed | 31% | 217 | 100 | >1,000 | |||||||||
| Surface or low | 49% | 135 | 100 | ||||||||||
| Ponderosa pine | Replacement | 5% | 200 | ||||||||||
| Mixed | 17% | 60 | |||||||||||
| Surface or low | 78% | 13 | |||||||||||
| Great Basin Forested | |||||||||||||
| Interior ponderosa pine | Replacement | 5% | 161 | 800 | |||||||||
| Mixed | 10% | 80 | 50 | 80 | |||||||||
| Surface or low | 86% | 9 | 8 | 10 | |||||||||
| Ponderosa pine-Douglas-fir | Replacement | 10% | 250 | >1,000 | |||||||||
| Mixed | 51% | 50 | 50 | 130 | |||||||||
| Surface or low | 39% | 65 | 15 | ||||||||||
| Great Basin Douglas-fir (dry) | Replacement | 12% | 90 | 600 | |||||||||
| Mixed | 14% | 76 | 45 | ||||||||||
| Surface or low | 75% | 14 | 10 | 50 | |||||||||
| Aspen with conifer (low to midelevations) | Replacement | 53% | 61 | 20 | |||||||||
| Mixed | 24% | 137 | 10 | ||||||||||
| Surface or low | 23% | 143 | 10 | ||||||||||
| Douglas-fir (warm mesic interior) | Replacement | 28% | 170 | 80 | 400 | ||||||||
| Mixed | 72% | 65 | 50 | 250 | |||||||||
| Aspen with conifer (high elevations) | Replacement | 47% | 76 | 40 | |||||||||
| Mixed | 18% | 196 | 10 | ||||||||||
| Surface or low | 35% | 100 | 10 | ||||||||||
| Stable aspen-cottonwood, no conifers | Replacement | 31% | 96 | 50 | 300 | ||||||||
| Surface or low | 69% | 44 | 20 | 60 | |||||||||
| Spruce-fir-pine (subalpine) | Replacement | 98% | 217 | 75 | 300 | ||||||||
| Mixed | 2% | >1,000 | |||||||||||
| Aspen with spruce-fir | Replacement | 38% | 75 | 40 | 90 | ||||||||
| Mixed | 38% | 75 | 40 | ||||||||||
| Surface or low | 23% | 125 | 30 | 250 | |||||||||
| Stable aspen without conifers | Replacement | 81% | 150 | 50 | 300 | ||||||||
| Surface or low | 19% | 650 | 600 | >1,000 | |||||||||
| Northern and Central Rockies | |||||||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||||||
| Northern and Central Rockies Grassland | |||||||||||||
| Northern prairie grassland | Replacement | 55% | 22 | 2 | 40 | ||||||||
| Mixed | 45% | 27 | 10 | 50 | |||||||||
| Mountain grassland | Replacement | 60% | 20 | 10 | |||||||||
| Mixed | 40% | 30 | |||||||||||
| Northern and Central Rockies Shrubland | |||||||||||||
| Riparian (Wyoming) | Mixed | 100% | 100 | 25 | 500 | ||||||||
| Wyoming big sagebrush | Replacement | 63% | 145 | 80 | 240 | ||||||||
| Mixed | 37% | 250 | |||||||||||
| Basin big sagebrush | Replacement | 60% | 100 | 10 | 150 | ||||||||
| Mixed | 40% | 150 | |||||||||||
| Low sagebrush shrubland | Replacement | 100% | 125 | 60 | 150 | ||||||||
| Mountain shrub, nonsagebrush | Replacement | 80% | 100 | 20 | 150 | ||||||||
| Mixed | 20% | 400 | |||||||||||
| Mountain big sagebrush steppe and shrubland | Replacement | 100% | 70 | 30 | 200 | ||||||||
| Northern and Central Rockies Woodland | |||||||||||||
| Ancient juniper | Replacement | 100% | 750 | 200 | >1,000 | ||||||||
| Northern and Central Rockies Forested | |||||||||||||
| Ponderosa pine (Northern Great Plains) | Replacement | 5% | 300 | ||||||||||
| Mixed | 20% | 75 | |||||||||||
| Surface or low | 75% | 20 | 10 | 40 | |||||||||
| Ponderosa pine (Northern and Central Rockies) | Replacement | 4% | 300 | 100 | >1,000 | ||||||||
| Mixed | 19% | 60 | 50 | 200 | |||||||||
| Surface or low | 77% | 15 | 3 | 30 | |||||||||
| Ponderosa pine (Black Hills, low elevation) | Replacement | 7% | 300 | 200 | 400 | ||||||||
| Mixed | 21% | 100 | 50 | 400 | |||||||||
| Surface or low | 71% | 30 | 5 | 50 | |||||||||
| Ponderosa pine (Black Hills, high elevation) | Replacement | 12% | 300 | ||||||||||
| Mixed | 18% | 200 | |||||||||||
| Surface or low | 71% | 50 | |||||||||||
| Ponderosa pine-Douglas-fir | Replacement | 10% | 250 | >1,000 | |||||||||
| Mixed | 51% | 50 | 50 | 130 | |||||||||
| Surface or low | 39% | 65 | 15 | ||||||||||
| Western redcedar | Replacement | 87% | 385 | 75 | >1,000 | ||||||||
| Mixed | 13% | >1,000 | 25 | ||||||||||
| Douglas-fir (xeric interior) | Replacement | 12% | 165 | 100 | 300 | ||||||||
| Mixed | 19% | 100 | 30 | 100 | |||||||||
| Surface or low | 69% | 28 | 15 | 40 | |||||||||
| Douglas-fir (warm mesic interior) | Replacement | 28% | 170 | 80 | 400 | ||||||||
| Mixed | 72% | 65 | 50 | 250 | |||||||||
| Douglas-fir (cold) | Replacement | 31% | 145 | 75 | 250 | ||||||||
| Mixed | 69% | 65 | 35 | 150 | |||||||||
| Grand fir-Douglas-fir-western larch mix | Replacement | 29% | 150 | 100 | 200 | ||||||||
| Mixed | 71% | 60 | 3 | 75 | |||||||||
| Mixed conifer-upland western redcedar-western hemlock | Replacement | 67% | 225 | 150 | 300 | ||||||||
| Mixed | 33% | 450 | 35 | 500 | |||||||||
| Western larch-lodgepole pine-Douglas-fir | Replacement | 33% | 200 | 50 | 250 | ||||||||
| Mixed | 67% | 100 | 20 | 140 | |||||||||
| Grand fir-lodgepole pine-larch-Douglas-fir | Replacement | 31% | 220 | 50 | 250 | ||||||||
| Mixed | 69% | 100 | 35 | 150 | |||||||||
| Persistent lodgepole pine | Replacement | 89% | 450 | 300 | 600 | ||||||||
| Mixed | 11% | >1,000 | |||||||||||
| Whitebark pine-lodgepole pine (upper subalpine, Northern and Central Rockies) | Replacement | 38% | 360 | ||||||||||
| Mixed | 62% | 225 | |||||||||||
| Lower subalpine lodgepole pine | Replacement | 73% | 170 | 50 | 200 | ||||||||
| Mixed | 27% | 450 | 40 | 500 | |||||||||
| Lower subalpine (Wyoming and Central Rockies) | Replacement | 100% | 175 | 30 | 300 | ||||||||
| Upper subalpine spruce-fir (Central Rockies) | Replacement | 100% | 300 | 100 | 600 | ||||||||
| Northern Great Plains | |||||||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||||||
| Northern Plains Grassland | |||||||||||||
| Northern mixed-grass prairie | Replacement | 67% | 15 | 8 | 25 | ||||||||
| Mixed | 33% | 30 | 15 | 35 | |||||||||
| Oak savanna | Replacement | 7% | 44 | ||||||||||
| Mixed | 17% | 18 | |||||||||||
| Surface or low | 76% | 4 | |||||||||||
| Northern Plains Woodland | |||||||||||||
| Oak woodland | Replacement | 2% | 450 | ||||||||||
| Surface or low | 98% | 7.5 | |||||||||||
| Northern Great Plains wooded draws and ravines | Replacement | 38% | 45 | 30 | 100 | ||||||||
| Mixed | 18% | 94 | |||||||||||
| Surface or low | 43% | 40 | 10 | ||||||||||
| Great Plains floodplain | Replacement | 100% | 500 | ||||||||||
| *Fire Severities— Replacement: Any fire that causes greater than 75% top removal of a vegetation-fuel type, resulting in general replacement of existing vegetation; may or may not cause a lethal effect on the plants. Mixed: Any fire burning more than 5% of an area that does not qualify as a replacement, surface, or low-severity fire; includes mosaic and other fires that are intermediate in effects. Surface or low: Any fire that causes less than 25% upper layer replacement and/or removal in a vegetation-fuel class but burns 5% or more of the area [148,213]. |
|||||||||||||
1. Adams, C. Phyllis; Viereck, Leslie A. 1992. Multivariate analysis of woody plant succession on the Tanana River in interior Alaska. In: Clary, Warren P.; McArthur, E. Durant; Bedunah, Don; Wambolt, Carl L., compilers. Proceedings--symposium on ecology and management of riparian shrub communities; 1991 May 29-31; Sun Valley, ID. Gen. Tech. Rep. INT-289. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 4-10. [19087]
2. Alexander, Robert R.; Hoffman, George R.; Wirsing, John M. 1986. Forest vegetation of the Medicine Bow National Forest in southeastern Wyoming: a habitat type classification. Res. Pap. RM-271. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 39 p. [307]
3. Allan, John S. 1962. The plant communities of the Big Cottonwood Canyon drainage. Salt Lake City, UT: University of Utah. 108 p. Thesis. [9104]
4. Alldredge, Matthew W.; Peek, James M.; Wall, William A. 2001. Alterations of shrub communities in relation to herbivory in northern Idaho. Northwest Science. 75(2): 137-144. [53493]
5. Allen, Arthur W.; Jordan, Peter A.; Terrell, James W. 1987. Habitat suitability index models: moose--Lake Superior region. Biol. Rep. 82 (10.155). Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service. 47 p. [11710]
6. Allen, Craig Daniel. 1989. Changes in the landscape of the Jemez Mountains, New Mexico. Berkeley, CA: University of California. 346 p. Dissertation. [42116]
7. Appleton, Bonnie Lee; Frenzel, Cindy L.; Hillegass, Julie B.; Lyons, Robert E.; Steward, Larry G. 2009. Virginia firescapes: Firewise landscaping for woodland homes. Virginia Cooperative Extension Publication 430-300. Blacksburg, VA: Virginia Polytechnic Institute and State University, Virginia Cooperative Extension; Virginia Firewise Landscaping Task Force. 9 p. Available online: http://pubs.ext.vt.edu/430/430-300/430-300.pdf [2009, October 6]. [76014]
8. Arkle, Robert S.; Pilliod, David S. 2010. Prescribed fires as ecological surrogates for wildfires: a stream and riparian perspective. Forest Ecology and Management. 259(5): 893-903. [80770]
9. Armstrong, Richard C.; Heston, Katherine. 1982. Control of woody invasion of a kettle bog. Restoration and Management Notes. 1: 18. [22529]
10. Arno, Stephen F. 1998. Fire ecology in Scandinavian forests. Journal of Forestry. 96(12): 20-23. [29306]
11. Arno, Stephen F. 2001. [Personal communication]. December 12. Missoula, MT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory [Retired]. In: FEIS log book. On file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [38714]
12. Arno, Stephen F.; Harrington, Michael G. 1998. The Interior West: managing fire-dependent forests by simulating natural disturbance regimes. In: Proceedings, Forest management into the next century: what will make it work; 1997 November 19-21; Spokane, WA. Madison, WI: Forest Products Society: 53-62. [43185]
13. Bach, Catherine E. 2001. Long-term effects of insect herbivory and sand accretion on plant succession on sand dunes. Ecology. 82(5): 1401-1416. [65387]
14. Baker, William L. 1988. Size-class structure of contiguous riparian woodlands along a Rocky Mountain river. Physical Geography. 9(1): 1-14. [9269]
15. Baker, William L. 1989. Classification of the riparian vegetation of the montane and subalpine zones in western Colorado. The Great Basin Naturalist. 49(2): 214-228. [7985]
16. Bakuzis, E. V.; Hansen, H. L. 1962. Ecographs of shrubs and other undergrowth species of Minnesota forest communities. Minnesota Forestry Notes. 117: 1-2. [10316]
17. Barrington, Mac; Bunting, Steve; Wright, Gerald. 1988. A fire management plan for Craters of the Moon National Monument. Draft. Subagreement No. 5 to Cooperative Agreement CA-9000-8-0005. Moscow, ID: University of Idaho, Range Resources Department. On file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 52 p. [1687]
18. Bartemucci, Paula; Messier, Christian; Canham, Charles D. 2006. Overstory influences on light attenuation patterns and understory plant community diversity and composition in southern boreal forests of Quebec. Canadian Journal of Forest Research. 36: 2065-2079. [65316]
19. Baskin, Carol C.; Baskin, Jerry M. 2001. Seeds: ecology, biogeography, and evolution of dormancy and germination. San Diego, CA: Academic Press. 666 p. [60775]
20. Bassett, Dick; Larson, Milo; Moir, Will. 1987. Forest and woodland habitat types (plant associations) of Arizona south of the Mogollon Rim and southwestern New Mexico. 2nd ed. Albuquerque, NM: U.S. Department of Agriculture, Forest Service, Southwestern Region. Variously paginated. [20308]
21. Beals, E. W.; Cottam, G.; Vogl, R. J. 1960. Influence of deer on vegetation of the Apostle Islands, Wisconsin. Journal of Wildlife Management. 24(1): 68-80. [67570]
22. Beals, E. W.; Cottam, Grant. 1960. The forest vegetation of the Apostle Islands, Wisconsin. Ecology. 41(4): 743-751. [62783]
23. Beaudry, Leisbet; Coupe, Ray; Delong, Craig; Pojar, Jim. 1999. Plant indicator guide for northern British Columbia: boreal, sub-boreal, and subalpine biogeoclimatic zones (BWBS, SBS, SBPS, and northern ESSF). Victoria, BC: British Columbia Ministry of Forests, Forestry Division Sciences Branch. 134 p. [70419]
24. Begin, Yves. 1990. The effects of shoreline transgression on woody plants, upper St. Lawrence estuary, Quebec. Journal of Coastal Research. 6(4): 815-827. [17681]
25. Belovsky, Gary E. 1981. Food plant selection by a generalist herbivore: the moose. Ecology. 62(4): 1020-1030. [64420]
26. Belovsky, Gary E. 1984. Summer diet optimization by beaver. The American Midland Naturalist. 111(2): 209-222. [64423]
27. Berry, Alison M.; Torrey, John G. 1985. Seed germination, seedling inoculation and establishment of Alnus spp. in containers in greenhouse trials. Plant and Soil. 87(1): 161-173. [79124]
28. Blood, Laura E.; Pitoniak, Hilary J.; Titus, Jonathan H. 2010. Seed bank of a bottomland swamp in western New York. Castanea. 75(1): 19-38. [80474]
29. Blood, Laura E.; Titus, Jonathan H. 2010. Microsite effects on forest regeneration in a bottomland swamp in western New York. Journal of the Torrey Botanical Society. 137(1): 88-102. [81830]
30. Bloom, Robin G.; Mallik, Azim U. 2006. Relationships between ericaceous vegetation and soil nutrient status in a post-fire Kalmia angustifolia-black spruce chronosequence. Plant and Soil. 289(1/2): 211-226. [66677]
31. Boelter, Don H.; Verry, Elon S. 1977. Peatland and water in the northern Lake States. Gen. Tech. Rep. NC-31. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 22 p. [8168]
32. Boggs, Keith; Hansen, Paul; Pfister, Robert; Joy, John. 1990. Classification and management of riparian and wetland sites in northwestern Montana. Draft Version 1. Missoula, MT: University of Montana, School of Forestry, Montana Forest and Conservation Experiment Station. 217 p. [8447]
33. Bonanno, Sandra E.; Leopold, Donald J.; St. Hilaire, Lisa R. 1998. Vegetation of a freshwater dune barrier under high and low recreational uses. Journal of the Torrey Botanical Society. 125(1): 40-50. [65377]
34. Bousquet, Jean; Cheliak, William M.; Lalonde, Maurice. 1988. Allozyme variation within and among mature populations of speckled alder (Alnus rugosa) and relationships with green alder (A. crispa). American Journal of Botany. 75(11): 1678-1686. [5148]
35. Bradley, Anne F.; Noste, Nonan V.; Fischer, William C. 1992. Fire ecology of forests and woodlands in Utah. Gen. Tech. Rep. INT-287. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 128 p. [18211]
36. Brand, Gary J. 1985. Environmental indices for common Michigan trees and shrubs. Res. Pap. NC-261. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 5 p. [14465]
37. Braun, E. Lucy. 1989. The woody plants of Ohio. Columbus, OH: Ohio State University Press. 362 p. [12914]
38. Brisson, Jacques; Cogliastro, Alain; Robert, Michel. 2006. Controlling speckled alder (Alnus incana ssp. rugosa) invasion in a wetland reserve of southern Quebec. Natural Areas Journal. 26(1): 78-83. [64513]
39. Brown, Bruce A. 1958. Interrelationship of brush populations and some site factors in jack pine stands of north central Minnesota. St. Paul, MN: University of Minnesota. 120 p. Dissertation. [68263]
40. Brown, David E.; Lowe, Charles H.; Hausler, Janet F. 1977. Southwestern riparian communities: their biotic importance and management in Arizona. In: Johnson, R. Roy; Jones, Dale A., tech. coords. Importance, preservation and management of riparian habitat: a symposium: Proceedings; 1977 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 201-211. [5348]
41. Brown, David T.; Doucet, G. Jean. 1991. Temporal changes in winter diet selection by white-tailed deer in a northern deer yard. Journal of Wildlife Management. 55(3): 361-376. [15406]
42. Brown, James K.; Smith, Jane Kapler, eds. 2000. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech Rep. RMRS-GTR-42-vol. 2. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 257 p. [36581]
43. Brumelis, G.; Carleton, T. J. 1988. The vegetation of postlogged black spruce lowlands in central Canada. I. Trees and tall shrubs. Canadian Journal of Forest Research. 18: 1470-1478. [9267]
44. Brumelis, G.; Carleton, T. J. 1989. The vegetation of post-logged black spruce lowlands in central Canada. II. Understory vegetation. Journal of Applied Ecology. 26: 321-339. [7864]
45. Bryant, John P.; Chapin, F. S., III; Clausen, T. P.; Reichardt, P. R. 1987. Effect of resource availability on woody plant-mammal interaction. In: Provenza, Frederick D.; Flinders, Jerran T.; McArthur, E. Durant, compilers. Proceedings--symposium on plant-herbivore interactions; 1985 August 7-9; Snowbird, UT. Gen. Tech. Rep. INT-222. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 3-8. [7327]
46. Buckman, Robert E. 1965. Silvicultural use of prescribed burning in the Lake States. In: Proceedings--Society of American Foresters meeting; 1964 September 27 - October 1; Denver, CO. Washington, DC: Society of American Foresters: 38-40. [8749]
47. Buech, Richard R.; Rugg, David J. 1989. Biomass relations of shrub components and their generality. Forest Ecology and Management. 26(4): 257-264. [7660]
48. Buech, Richard R.; Rugg, David J. 1995. Biomass relations for components of five Minnesota shrubs. Research Paper NC-325. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 14 p. [63750]
49. Burgason, Barry N. 1976. Prescribed burning for management of hawthorn and alder. New York Fish and Game Journal. 23(2): 160-169. [14317]
50. Burgess, Stephen A.; Bider, J. R. 1980. Effects of stream habitat improvements on invertebrates, trout populations, and mink activity. Journal of Wildlife Management. 44(4): 871-880. [25982]
51. Buse, L. J.; Bell, F. W. 1992. Critical silvics of selected crop and competitor species in northwestern Ontario. Thunder Bay, ON: Ontario Ministry of Natural Resources, Northwestern Ontario Forest Technology Development Unit. 138 p. [30340]
52. Butler, Lem G.; Kielland, Knut. 2008. Acceleration of vegetation turnover and element cycling by mammalian herbivory in riparian ecosystems. Journal of Ecology. 96(1): 136-144. [78493]
53. Callaway, Ragan M. 1995. Positive interactions among plants. The Botanical Review. 61(4): 306-349. [27528]
54. Callaway, Ragan M.; Walker, Lawrence R. 1997. Competition and facilitation: a synthetic approach to interactions in plant communities. Ecology. 78(7): 1958-1965. [27660]
55. Cargill, Susan M.; Chapin, F. Stuart, III. 1987. Application of successional theory to tundra restoration: a review. Arctic and Alpine Research. 19(4): 366-372. [8685]
56. Carleton, T. J.; Maycock, P. F. 1980. Vegetation of the boreal forests south of James Bay: non-centered component analysis of the vascular flora. Ecology. 61(5): 1199-1212. [14734]
57. Carleton, Terence J.; MacLellan, Patricia. 1994. Woody vegetation responses to fire versus clear-cutting logging: a comparative survey in the central Canadian boreal forest. Ecoscience. 1(2): 141-152. [26054]
58. Carlson, Jack R. 1992. Selection, production, and use of riparian plant materials for the western United States. In: Landis, Thomas D., technical coordinator. Proceedings, Intermountain Forest Nursery Association; 1991 August 12-16; Park City, UT. Gen. Tech. Rep. RM-211. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 55-67. [20926]
59. Carlson, Jack R.; Conaway, Gary L.; Gibbs, Jacy L.; Hoag, J. Chris. 1992. Design criteria for revegetation in riparian zones of the Intermountain area. In: Clary, Warren P.; McArthur, E. Durant; Bedunah, Don; Wambolt, Carl L., compilers. Proceedings--symposium on ecology and management of riparian shrub communities; 1991 May 29-31; Sun Valley, ID. Gen. Tech. Rep. INT-289. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 145-150. [19109]
60. Case, Richard L.; Kauffman, J. Boone. 1997. Wild ungulate influences on the recovery of willows, black cottonwood, and thin-leaf alder following cessation of cattle grazing in northeastern Oregon. Northwest Science. 71(2): 115-126. [27671]
61. Chadde, Steve W.; Shelly, J. Stephen; Bursik, Robert J.; Moseley, Robert K.; Evenden, Angela G.; Mantas, Maria; Rabe, Fred; Heidel, Bonnie. 1998. Peatlands on national forests of the Northern Rocky Mountains: ecology and conservation. Gen. Tech. Rep. RMRS-GTR-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 75 p. [29130]
62. Chan, Franklin J.; Wong, Raymond M. 1989. Reestablishment of native riparian species at an altered high elevation site. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 428-435. [13771]
63. Chapin, David M.; Beschta, Robert L.; Shen, Hsieh Wen. 2002. Relationships between flood frequencies and riparian plant communities in the upper Klamath Basin, Oregon. Journal of the American Water Resources Association. 38(3): 603-617. [43834]
64. Chapman, William K.; Bessette, Alan E. 1990. Trees and shrubs of the Adirondacks. Utica, NY: North Country Books. 131 p. [12766]
65. Chappell, Christopher B.; Kagan, Jimmy. 2001. 24. Montane coniferous wetlands. In: Chappell, Christopher B.; Crawford, Rex C.; Barrett, Charley; Kagan, Jimmy; Johnson, David H.; O'Mealy, Mikell; Green, Greg A.; Ferguson, Howard L.; Edge, W. Daniel; Greda, Eva L.; O'Neil, Thomas A. Wildlife habitats: descriptions, status, trends, and system dynamics. In: Johnson, David H.; O'Neil, Thomas A., managing directors. Wildlife-habitat relationships in Oregon and Washington. Corvallis, OR: Oregon State University Press: 97-98. [68138]
66. Charron, I.; Johnson, E. A. 2006. The importance of fires and floods on tree ages along mountainous gravel-bed streams. Ecological Applications. 16(5): 1757-1770. [64733]
67. Christensen, E. M.; Clausen, J. J. (Jones); Curtis, J. T. 1959. Phytosociology of the lowland forests of northern Wisconsin. The American Midland Naturalist. 62(1): 232-247. [49627]
68. Chrosciewicz, Z. 1976. Burning for black spruce regeneration on a lowland cutover site in southeastern Manitoba. Canadian Journal of Forest Research. 6(2): 179-186. [7280]
69. Chrosciewicz, Z. 1983. Jack pine regeneration following postcut burning and seeding in southeastern Manitoba. Information Report NOR-X-252. Edmonton, AB: Environment Canada, Canadian Forestry Service, Northern Forest Research Centre. 10 p. [16917]
70. Clein, Joy S.; Schimel, Joshua P. 1995. Nitrogen turnover and availability during succession from alder to poplar in Alaskan taiga forests. Soil Biology and Biochemistry. 27(6): 743-752. [26503]
71. Coates, D.; Haeussler, S. 1986. A preliminary guide to the response of major species of competing vegetation to silvicultural treatments. Land Management Handbook No. 9. Victoria, BC: Ministry of Forests, Information Services Branch. 88 p. [17453]
72. Conard, S. G.; Radosevich, S. R. 1982. Post-fire succession in white fir (Abies concolor) vegetation of the northern Sierra Nevada. Madrono. 29(1): 42-56. [4931]
73. Conway, Verona M. 1949. The bogs of central Minnesota. Ecological Monographs. 19(2): 173-206. [16686]
74. Cooper, Stephen V.; Neiman, Kenneth E.; Roberts, David W. 1991. Forest habitat types of northern Idaho: a second approximation. [Revised]. Gen. Tech. Rep. INT-236. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 143 p. [14792]
75. Cooper, William S. 1930. The seed-plants and ferns of the Glacier Bay National Monument, Alaska. Bulletin of the Torrey Botanical Club. 57(5): 327-338. [65666]
76. Cooper, William Skinner. 1923. The recent ecological history of Glacier Bay, Alaska: the present vegetation cycle. Ecology. 4(3): 223-246. [65668]
77. Cormack, R. G. H. 1949. A study of trout streamside cover in logged-over and undisturbed virgin spruce woods. Canadian Journal of Research. 27: 78-95. [15689]
78. Cormack, R. G. H. 1953. A survey of coniferous forest succession in the eastern Rockies. Forestry Chronicle. 29: 218-232. [16458]
79. Coveney, Mary Camilla. 1965. Effect of fire and wind-throw on a forest area. Boston, MA: Boston University. 142 p. Dissertation. [35683]
80. Cowardin, Lewis M.; Carter, Virginia; Golet, Francis C.; LaRoe, Edward T. 1979. Classification of wetlands and deepwater habitats of the United States. FWS/OBS-79/31. Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service. 131 p. [41938]
81. Cox, Donald D. 2002. Marshes and swamps. In: A naturalist's guide to wetland plants: An ecology for eastern North America. Syracuse, NY: Syracuse University Press: 64-78. [69694]
82. Crawford, Rex C.; Kagan, Jimmy. 2001. 25. Eastside riparian-wetlands. In: Chappell, Christopher B.; Crawford, Rex C.; Barrett, Charley; Kagan, Jimmy; Johnson, David H.; O'Mealy, Mikell; Green, Greg A.; Ferguson, Howard L.; Edge, W. Daniel; Greda, Eva L.; O'Neil, Thomas A. Wildlife habitats: descriptions, status, trends, and system dynamics. In: Johnson, David H.; O'Neil, Thomas A., managing directors. Wildlife-habitat relationships in Oregon and Washington. Corvallis, OR: Oregon State University Press: 98-100. [67918]
83. Croonquist, Mary Jo; Brooks, Robert P. 1993. Effects of habitat disturbance on bird communities in riparian corridors. Journal of Soil and Water Conservation. 48(1): 65-70. [21107]
84. Crowder, A.; Harmsen, R. 1998. Notes on forest succession in old fields in southeastern Ontario: the woody species. The Canadian Field-Naturalist. 112(3): 410-418. [35971]
85. Curtis, John T. 1959. The vegetation of Wisconsin. Madison, WI: The University of Wisconsin Press. 657 p. [7116]
86. Damman, A. W. H. 1964. Some forest types of central Newfoundland and their relation to environmental factors. Forest Science Monograph 8. Washington, DC: Society of American Foresters. 62 p. [14281]
87. Damman, A. W. H.; Johnston, William F. 1980. Black spruce. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 11-14. [49853]
88. Dansereau, Pierre. 1959. Phytogeographia Laurentiana. II. The principal plant associations of the Saint Lawrence Valley. Contributions of the Botanical Institute No. 75. Montreal, PQ: University of Montreal, Botanical Institute. 147 p. [8925]
89. Dansereau, Pierre; Segadas-Vianna, Fernando. 1952. Ecological study of the peat bogs of eastern North America. Canadian Journal of Botany. 30(5): 490-520. [8869]
90. Darlington, H. Clayton. 1943. Vegetation and substrate of Cranberry Glades, West Virginia. Botanical Gazette. 104(3): 371-393. [64491]
91. Davis, John H., Jr. 1930. Vegetation of the Black Mountains of North Carolina: an ecological study. Journal of the Elisha Mitchell Scientific Society. 45: 291-318. [64613]
92. Davis, Kathleen M.; Clayton, Bruce D.; Fischer, William C. 1980. Fire ecology of Lolo National Forest habitat types. INT-79. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 77 p. [5296]
93. Densmore, Roseann Van Essen. 1979. Aspects of the seed ecology of woody plants of the Alaskan taiga and tundra. Durham, NC: Duke University. 285 p. Dissertation. [70495]
94. Dibble, Alison C.; Patterson, William A., III; White, Robert H. 2003. Relative flammability of native and invasive exotic plants of the northeastern U.S. In: Bennett, Karen P.; Dibble, Alison C.; Patterson, William A., III, compilers. Using fire to control invasive plants: what's new, what works in the Northeast?--2003 workshop proceedings; 2003 January 24; Portsmouth, NH. Durham, NH: University of New Hampshire, Cooperative Extension: 34-37. [81701]
95. Dick-Peddie, William A. 1993. New Mexico vegetation: past, present, and future. Albuquerque, NM: University of New Mexico Press. 244 p. [21097]
96. Dick-Peddie, William A.; Hubbard, John P. 1977. Classification of riparian vegetation. In: Johnson, R. Roy; Jones, Dale A., tech. coords. Importance, preservation and management of riparian habitat: a symposium: Proceedings; 1977 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 85-90. Available from: NTIS, Springfield, VA 22151; PB-274 582. [5338]
97. Dirschl, H. J.; Coupland, R. T. 1972. Vegetation patterns and site relationships in the Saskatchewan River delta. Canadian Journal of Botany. 50: 647-675. [7449]
98. Dirschl, Herman J.; Dabbs, Don L.; Gentle, Garry C. 1974. Landscape classification and plant successional trends in the Peace-Athabasca Delta. Canadian Wildlife Service Report Series 30. Ottawa, ON: Canadian Wildlife Service. 33 p. [6177]
99. Doerr, Phillip D.; Keith, Lloyd B.; Rusch, Donald H. 1971. Effects of fire on a ruffed grouse population. In: Proceedings, annual Tall Timbers fire ecology conference; 1970 August 20-21; Fredericton, NB. No. 10. Tallahassee, FL: Tall Timbers Research Station: 25-46. [14320]
100. Donahue, William H. 1954. Some plant communities in the Anthracite Region of northeastern Pennsylvania. The American Midland Naturalist. 51(1): 203-231. [64481]
101. Donkor, Nobel T.; Fryxell, John M. 1999. Impact of beaver foraging on structure of lowland boreal forests of Algonquin Provincial Park, Ontario. Forest Ecology and Management. 118(1/3): 83-92. [64353]
102. Donkor, Noble T. 2007. Impact of beaver (Castor canadensis Kuhl) foraging on species composition of boreal forests. In: Johnson, Edward A.; Miyanishi, Kiyoko, eds. Plant disturbance ecology: The process and the response. Boston, MA: Academic Press: 579-602. [69473]
103. Doucet, Christine M.; Adams, Ian T.; Fryxell, John M. 1994. Beaver dam and cache composition: are woody species used differently? Ecoscience. 1(3): 268-270. [64355]
104. Dreesen, David R.; Harrington, John T. 1997. Propagation of native plants for restoration projects in the southwestern U.S.--preliminary investigations. In: Landis, Thomas D.; Thompson, Jan R., tech. coords. National proceedings: forest and conservation nursery associations--1997; Regeneration, reforestation, restoration: The seedling is the key; 1997 August 11-14; Bemidji, MN; 1997 August 19-21; Boise, ID. Gen. Tech. Rep. PNW-GTR-419. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 77-88. [29122]
105. Duncan, Wilbur H.; Duncan, Marion B. 1987. The Smithsonian guide to seaside plants of the Gulf and Atlantic coasts from Louisiana to Massachusetts, exclusive of lower peninsular Florida. Washington, DC: Smithsonian Institution Press. 409 p. [12906]
106. Duncan, Wilbur H.; Duncan, Marion B. 1988. Trees of the southeastern United States. Athens, GA: The University of Georgia Press. 322 p. [12764]
107. Dunham, Jason B.; Rosenberger, Amanda E.; Luce, Charlie H.; Rieman, Bruce E. 2007. Influences of wildfire and channel reorganization on spatial and temporal variation in stream temperature and the distribution of fish and amphibians. Ecosystems. 10(2): 335-346. [68647]
108. Dunn, Christopher P. 1986. Shrub layer response to death of Ulmus americana in southeastern Wisconsin lowland forests. Bulletin of the Torrey Botanical Club. 113(2): 142-148. [4793]
109. Durkin, Paula; Muldavin, Esteban; Bradley, Mike; Carr, Stacey E. 1996. A preliminary riparian/wetland vegetation community classification of the upper and middle Rio Grande watersheds in New Mexico. In: Shaw, Douglas W.; Finch, Deborah M., technical coordinators. Desired future conditions for southwestern riparian ecosystems: bringing interests and concerns together: Proceedings; 1995 September 18-22; Albuquerque, NM. Gen. Tech. Rep. RM-GTR-272. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 44-57. [26192]
110. Dutro, Ruth; Cohoe, Edith. 1938. An ecological study of Wolf's Bog, Cheboygan County, Michigan. Transactions of the Kansas Academy of Science. 41(4): 87-95. [80285]
111. Dwire, Kathleen A.; Kauffman, J. Boone. 2003. Fire and riparian ecosystems in landscapes of the western USA. In: Young, Michael K.; Gresswell, Robert E.; Luce, Charles H., eds. Selected papers from an international symposium on effects of wildland fire on aquatic ecosystems in the western USA; 2002 April 22-24; Boise, ID. In: Forest Ecology and Management. Special Issue: The effects of wildland fire on aquatic ecosystems in the western USA. New York: Elsevier Science B. V.; 178(1-2): 61-74. [44923]
112. Ehrenfeld, Joan G.; Culick, Martha. 1981. Structure and dynamics of hardwood swamps in the New Jersey pine barrens: contrasting patterns in trees and shrubs. American Journal of Botany. 68(4): 471-481. [22904]
113. Ehrhart, Robert C.; Hansen, Paul L. 1998. Successful strategies for grazing cattle in riparian zones. Montana BLM Riparian Technical Bulletin No. 4; BLM/MT/ST-98/001+1610. Billings, MT: U.S. Department of the Interior, Bureau of Land Management, Montana State Office. 48 p. [82578]
114. Evenden, Angela G. 1989. Ecology and distribution of riparian vegetation in the Trout Creek Mountains of southeastern Oregon. Corvallis, OR: Oregon State University. 156 p. Dissertation. [10231]
115. Everett, Richard; Schellhaas, Richard; Ohlson, Pete; Spurbeck, Don; Keenum, David. 2003. Continuity in fire disturbance between riparian and adjacent sideslope Douglas-fir forests. Forest Ecology and Management. 175: 31-47. [43635]
116. Eyre, F. H.; Gagnon, Gilles. 1980. Aspen. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 16-17. [49858]
117. Fessenden, R. J. 1979. Use of actinorrhizal plants for land reclamation and amenity planting in the U.S.A. and Canada. In: Gordon, J. C.; Wheeler, C. T.; Perry, D. A., eds. Symbiotic nitrogen fixation in the management of temperate forests: Proceedings of a workshop; 1979 April 2-5; Corvallis, OR. Corvallis, OR: Oregon State University, Forest Research Laboratory: 403-419. [4308]
118. Fites, Jo Ann. 1993. Ecological guide to mixed conifer plant associations--northern Sierra Nevada and southern Cascades: Lassen, Plumas, Tahoe, and El Dorado National Forests. R5-ECOL-TP-001. Vallejo, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Region. 120 p. [82332]
119. Fites-Kaufman, Joann; Bradley, Anne F.; Merrill, Amy G. 2006. Fire and plant interactions. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 94-117. [65534]
120. Flora of North America Editorial Committee, eds. 2011. Flora of North America North of Mexico, [Online]. Flora of North America Association (Producer). Available: http://www.efloras.org/flora_page.aspx?flora_id=1. [36990]
121. Foote, Geoffrey G. 1965. Phytosociology of the bottomland hardwood forests in western Montana. Missoula, MT: University of Montana. 140 p. Thesis. [17369]
122. Fortney, Ronald H. 2000. Plant communities of West Virginia wetlands. West Virginia Academy of Science. 72(3): 41-54. [68447]
123. Fortney, Ronald H.; Rentch, James S. 2003. Post logging era plant successional trends and geospatial vegetation patterns in Canaan Valley, West Virginia, 1945 to 2000. Castanea. 68(4): 317-334. [52648]
124. Frank, Robert M.; Majcen, Zoran; Gagnon, Gilles. 1980. Balsam fir. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 10-11. [49851]
125. Frederick, Clara May. 1974. A natural history study of the vascular flora of Cedar Bog, Champaign County, Ohio. Ohio Journal of Science. 74(2): 65-116. [23770]
126. Friedman, Jonathan M.; Auble, Gregor T.; Andrews, Edmund D.; Kittel, Gwen; Madole, Richard F.; Griffin, Eleanor R.; Allred, Tyler M. 2006. Transverse and longitudinal variation in woody riparian vegetation along a montane river. Western North American Naturalist. 66(1): 78-91. [62273]
127. Furlow, John J. 1979. The systematics of the American species of Alnus (Betulaceae)--II. Rhodora. 81(826): 151-248. [8795]
128. Furyaev, V. V.; Wein, Ross W.; MacLean, David A. 1983. Fire influences in Abies-dominated forests. In: Wein, Ross W.; MacLean, David A., eds. The role of fire in northern circumpolar ecosystems. Scope 18. New York: John Wiley & Sons: 221-234. [14610]
129. Gates, Frank C. 1942. The bogs of northern Lower Michigan. Ecological Monographs. 12(3): 213-254. [10728]
130. Gill, C. J. 1970. The flooding tolerance of woody species--a review. Forestry Abstracts. 31: 671-688. [42161]
131. Gleason, Henry A.; Cronquist, Arthur. 1991. Manual of vascular plants of northeastern United States and adjacent Canada. 2nd ed. New York: New York Botanical Garden. 910 p. [20329]
132. Goldman, Charles R. 1961. The contribution of alder trees (Alnus tenuifolia) to the primary productivity of Castle Lake, California. Ecology. 42(2): 282-288. [68646]
133. Gordon, John C.; Hall, Richard B. 1978. Alder research outside the Northwest: a brief overview. In: Briggs, David G.; DeBell, Dean S.; Atkinson, William A., comps. Utilization and management of alder: Proceedings of a symposium; 1977 April 25-27; Ocean Shores, WA. Gen. Tech. Rep. PNW-70. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: 47-53. [60834]
134. Grandtner, M. M.; Ducruc, Jean-Pierre. 1980. Black spruce-tamarack. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 14. [49855]
135. Grant, Martin L. 1929. The burn succession in Itasca County, Minnesota. Minneapolis, MN: University of Minnesota. 63 p. Thesis. [36527]
136. Grant, Martin L. 1934. The climax forest community in Itasca County, Minnesota, and its bearing upon the successional status of the pine community. Ecology. 15(3): 243-257. [64486]
137. Great Plains Flora Association. 1986. Flora of the Great Plains. Lawrence, KS: University Press of Kansas. 1392 p. [1603]
138. Green, Douglas M.; Kauffman, J. Boone. 1995. Succession and livestock grazing in a northeastern Oregon riparian ecosystem. Journal of Range Management. 48(4): 307-313. [25925]
139. Grodzinski, Wladyslaw; Sawicka-Kapusta, Katarzyna. 1970. Energy values of tree-seeds eaten by small mammals. Oikos. 21: 52-58. [41851]
140. Guglielmo, Christopher G.; Karasov, William H. 1995. Nutritional quality of winter browse for ruffed grouse. The Journal of Wildlife Management. 59(3): 427-436. [81986]
141. Gullion, Gordon W. 1964. Wildlife uses of Nevada plants. Contributions toward a flora of Nevada: No. 49. CR-24-64. Beltsville, MD: U.S. Department of Agriculture, Agricultural Research Service, Crops Research Division; Washington, DC: U.S. National Arboretum, Herbarium. 170 p. [6729]
142. Haeussler, S.; Coates, D. 1986. Autecological characteristics of selected species that compete with conifers in British Columbia: a literature review. Land Management Report No. 33. Victoria, BC: Ministry of Forests, Information Services Branch. 180 p. [1055]
143. Haeussler, S.; Coates, D.; Mather, J. 1990. Autecology of common plants in British Columbia: A literature review. Economic and Regional Development Agreement: FRDA Report 158. Victoria, BC: Forestry Canada, Pacific Forestry Centre; British Columbia Ministry of Forests, Research Branch. 272 p. [18033]
144. Hall, Frederick C. 1973. Plant communities of the Blue Mountains in eastern Oregon and southeastern Washington. R6 Area Guide 3-1. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 82 p. [1059]
145. Hall, Frederick C. 2005. Emigrant Creek cattle allotment: lessons from 30 years of photomonitoring. Gen. Tech. Rep. PNW-GTR-639. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 37 p. [61823]
146. Hamilton, Evelyn H. 2006. Fire effects and post-burn vegetation development in the Sub-Boreal Spruce zone: Mackenzie (Windy Point) site. Technical Report 033. Victoria, BC: Ministry of Forests and Range, Forest Science Program. 19 p. Available online: http://www.for.gov.bc.ca/hfd/pubs/Docs/Tr/Tr033.pdf [2008, October 1]. [64177]
147. Hamilton, Evelyn H.; Yearsley, H. Karen. 1988. Vegetation development after clearcutting and site preparation in the SBS zone. Economic and Regional Development Agreement: FRDA Report 018. Victoria, BC: Canadian Forestry Service, Pacific Forestry Centre; British Columbia Ministry of Forests and Lands. 66 p. [8760]
148. Hann, Wendel; Havlina, Doug; Shlisky, Ayn; [and others]. 2010. Interagency fire regime condition class (FRCC) guidebook, [Online]. Version 3.0. In: FRAMES (Fire Research and Management Exchange System). National Interagency Fuels, Fire & Vegetation Technology Transfer (NIFTT) (Producer). Available: http://www.fire.org/niftt/released/FRCC_Guidebook_2010_final.pdf. [81749]
149. Hansen, Paul L.; Chadde, Steve W.; Pfister, Robert D. 1988. Riparian dominance types of Montana. Misc. Publ. No. 49. Missoula, MT: University of Montana, School of Forestry, Montana Forest and Conservation Experiment Station. 411 p. [5660]
150. Hansen, Paul L.; Hall, James B. 2002. Classification and management of USDI Bureau of Land Management's riparian and wetland sites in eastern and southern Idaho. Corvallis, MT: Bitterroot Restoration. 304 p. [82582]
151. Hansen, Paul L.; Pfister, Robert D.; Boggs, Keith; Cook, Bradley J.; Joy, John; Hinckley, Dan K. 1995. Classification and management of Montana's riparian and wetland sites. Miscellaneous Publication No. 54. Missoula, MT: The University of Montana, School of Forestry, Montana Forest and Conservation Experiment Station. 646 p. [24768]
152. Harper, Karen A.; Lesieur, Daniel; Bergeron, Yves; Drapeau, Pierre. 2004. Forest structure and composition at young fire and cut edges in black spruce boreal forest. Canadian Journal of Forest Research. 34: 289-302. [62295]
153. Harper, Karen A.; Macdonald, S. Ellen. 2002. Structure and composition of edges next to regenerating clear-cuts in mixed-wood boreal forest. Journal of Vegetation Science. 13: 535-546. [45071]
154. Harrington, Constance A.; Brodie, Leslie Chandler; DeBell, Dean S.; Schopmeyer, C. S. 2008. Alnus P. Mill.: alder. In: Bonner, Franklin T., Karrfalt, Robert P., eds. Woody plant seed manual. Agric. Handbook No. 727. Washington, DC: U.S. Department of Agriculture, Forest Service: 232-242. [79017]
155. Harrington, H. D. 1964. Manual of the plants of Colorado. 2nd ed. Chicago, IL: The Swallow Press. 666 p. [6851]
156. Harris, Richard R. 1989. Riparian communities of the Sierra Nevada and their environmental relationships. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 393-398. [13768]
157. Hart, J. 1976. Montana native plants and early peoples. Helena, MT: Montana Historical Society. 75 p. [9979]
158. Harvey, B. D.; Bergeron, Y. 1989. Site patterns of natural regeneration following clear-cutting in northwestern Quebec. Canadian Journal of Forest Research. 19: 1458-1469. [9844]
159. Hawkes, B. C.; Feller, M. C.; Meehan, D. 1990. Site preparation: fire. In: Lavender, D. P.; Parish, R.; Johnson, C. M.; Montgomery, G.; Vyse, A.; Willis, R. A.; Winston, D., eds. Regenerating British Columbia's forests. Vancouver, BC: University of British Columbia Press: 131-149. [10712]
160. Healy, William M.; Gill, John D. 1974. Alders. In: Gill, John D.; Healy, William M., compilers. Shrubs and vines for Northeastern wildlife. Gen. Tech. Rep. NE-9. Broomall, PA: U.S. Department of Agriculture, Forest Service: 6-9. [6208]
161. Heidorn, Randy. 1991. Vegetation management guideline: exotic buckthorns--common buckthorn (Rhamnus cathartica L.), glossy buckthorn (Rhamnus frangula L.), Dahurian buckthorn (Rhamnus davurica Pall.). Natural Areas Journal. 11(4): 216-217. [16332]
162. Heilman, Paul E. 1966. Changes in distribution and availability of nitrogen with forest succession on north slopes in interior Alaska. Ecology. 47(5): 825-831. [81981]
163. Heinselman, M. L. 1970. Landscape evolution, peatland types and the environment in the Lake Agassiz Peatlands Natural Area, Minnesota. Ecological Monographs. 40(2): 235-261. [8378]
164. Heinselman, Miron L. 1981. Fire and succession in the conifer forests of northern North America. In: West, Darrell C.; Shugart, Herman H.; Botkin, Daniel B., eds. Forest succession: concepts and applications. New York: Springer-Verlag: 374-405. [29237]
165. Heinselman, Miron L. 1981. Fire intensity and frequency as factors in the distribution and structure of northern ecosystems. In: Mooney, H. A.; Bonnicksen, T. M.; Christensen, N. L.; Lotan, J. E.; Reiners, W. A., technical coordinators. Fire regimes and ecosystem properties: Proceedings of the conference; 1978 December 11-15; Honolulu, HI. Gen. Tech. Rep. WO-26. Washington, DC: U.S. Department of Agriculture, Forest Service: 7-57. [4390]
166. Hess, Karl; Alexander, Robert R. 1986. Forest vegetation of the Arapaho and Roosevelt National Forests in central Colorado: a habitat type classification. Res. Pap. RM-266. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 48 p. [1141]
167. Hickman, James C., ed. 1993. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 1400 p. [21992]
168. Hiebert, Ronald D.; Hamrick, J. L. 1983. Patterns and levels of genetic variation in Great Basin bristlecone pine, Pinus longaeva. Evolution. 37(2): 302-310. [25931]
169. Hitchcock, C. Leo; Cronquist, Arthur. 1964. Vascular plants of the Pacific Northwest. Part 2: Salicaceae to Saxifragaceae. Seattle, WA: University of Washington Press. 597 p. [1166]
170. Holland, Marjorie M.; Burk, C. John; McLain, David. 2000. Long-term vegetation dynamics of the lower strata of a western Massachusetts oxbow swamp forest. Rhodora. 102(910): 154-174. [40164]
171. Hosie, R. C. 1969. Native trees of Canada. 7th ed. Ottawa, ON: Canadian Forestry Service, Department of Fisheries and Forestry. 380 p. [3375]
172. Houston, Kent E.; Hartung, Walter J.; Hartung, Carol J. 2001. A field guide for forest indicator plants, sensitive plants, and noxious weeds of the Shoshone National Forest, Wyoming. Gen. Tech. Rep. RMRS-GTR-84. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 184 p. [40585]
173. Hudak, Howard G.; Ketcheson, Gary L. 1992. Willow community types as influenced by valley bottom and stream types. In: Clary, Warren P.; McArthur, E. Durant; Bedunah, Don; Wambolt, Carl L., compilers. Proceedings--symposium on ecology and management of riparian shrub communities; 1991 May 29-31; Sun Valley, ID. Gen. Tech. Rep. INT-289. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 16-17. [19089]
174. Huenneke, Laura Foster. 1983. Understory response to gaps caused by the death of Ulmus americanus in central New York. Bulletin of the Torrey Botanical Club. 110(2): 170-175. [4934]
175. Huenneke, Laura Foster. 1985. Spatial distribution of genetic individuals in thickets of Alnus incana ssp. rugosa, a clonal shrub. American Journal of Botany. 72(1): 152-158. [81983]
176. Huenneke, Laura Foster. 1987. Demography of a clonal shrub, Alnus incana ssp. rugosa (Betulaceae). The American Midland Naturalist. 117(1): 43-55. [14327]
177. Hughes, Francine M. R.; Harris, Tim; Richards, Keith; Pautou, Guy; El Hames, Ahmed; Barsoum, Nadia; Girel, Jacqui; Peiry, Jean-Luc; Foussadier, Remi. 1997. Woody riparian species response to different soil moisture conditions: laboratory experiments on Alnus incana (L.) Moench. Global Ecology and Biogeography Letters. 6(3-4): 247-256. [81984]
178. Hughes, Jeffrey W.; Cass, Wendy B. 1997. Pattern and process of a floodplain forest, Vermont, USA: predicted responses of vegetation to perturbation. Journal of Applied Ecology. 34(3): 594-612. [65684]
179. Hulten, Eric. 1968. Flora of Alaska and neighboring territories. Stanford, CA: Stanford University Press. 1008 p. [13403]
180. Hurd, T. M.; Raynal, D. J.; Schwintzer, C. R. 2001. Symbiotic N2 fixation of Alnus incana ssp. rugosa in shrub wetlands of the Adirondack Mountains, New York, USA. Oecologia. 126(1): 94-103. [47080]
181. Husari, Susan. 1980. Fire ecology of the vegetative habitat types in the Lassen Fire Management Planning Area (Caribou Wilderness and Lassen Volcanic National Park). In: Swanson, John R.; Johnson, Robert C.; Merrifield, Dave; Dennestan, Alan. Fire Management Plan: Lassen Fire Management Planning Area. Lassen Volcanic National Park-Caribou Wilderness Unit--Implementation plan. Mineral, CA: U.S. Department of the Interior, National Park Service, Lassen Volcanic National Park; Susanville, CA: U.S. Department of Agriculture, Forest Service, Lassen National Forest: Appendix 3: 1-23. [21408]
182. Jacobson, George L., Jr.; Almquist-Jacobson, Heather; Winne, J. Chris. 1991. Conservation of rare plant habitat: insights from the recent history of vegetation and fire at Crystal Fen, northern Maine, USA. Biological Conservation. 57(3): 287-314. [16533]
183. Java, Bonita J.; Everett, Richard L. 1992. Rooting hardwood cuttings of Sitka and thinleaf alder. In: Clary, Warren P.; McArthur, E. Durant; Bedunah, Don; Wambolt, Carl L., compilers. Proceedings--symposium on ecology and management of riparian shrub communities; 1991 May 29-31; Sun Valley, ID. Gen. Tech. Rep. INT-289. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 138-141. [19107]
184. Johnson, D. 1968. Taxonomy and distribution of northwestern alders. In: Trappe, J. M.; Franklin, J. F.; Tarrant, R. F.; Hansen, G. M., ed. Biology of alder; 1967 April 14-15; Pullman, WA. Portland, OR: U. S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: 9-22. [6187]
185. Johnston, Barry C. 1987. Plant associations of Region 2: Potential plant communities of Wyoming, South Dakota, Nebraska, Colorado, and Kansas. 4th ed. R2-ECOL-87-2. Lakewood, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Region. 429 p. [54304]
186. Johnston, William F. 1980. Northern white-cedar. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 23-24. [49885]
187. Johnston, William F. 1980. Tamarck. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 15-16. [49857]
188. Jones, Keith R. 1984. Site classification in Ontario. In: Bockheim, James G., ed. Forest land classification: experiences, problems, perspectives: Proceedings of a symposium; 1984 March 18-20; Madison, WI. Madison, WI: University of Wisconsin, Department of Soil Science: 82-99. [82274]
189. Jones, R. Keith; Pierpoint, Geoffrey; Wickware, Gregory M.; Jeglum, John K.; Arnup, Robert W.; Bowles, Jane M. 1983. Field guide to forest ecosystem classification for the Clay Belt, site region 3e. Toronto, ON: Ministry of Natural Resources, Ontario Forest Research Institute. 160 p. [16163]
190. Kari, Priscilla Russell. 1987. Tanaina plantlore. Dena'ina K'et'una: An ethnobotany of the Dena'ina Indians of southcentral Alaska. 2nd ed. [Revised]. Anchorage, AK: U.S. Department of the Interior, National Park Service, Alaska Region. 205 p. [67343]
191. Karrenberg, S.; Blaser, S.; Kollman, J.; Speck, T.; Edwards, P. J. 2003. Root anchorage of saplings and woody cuttings of woody pioneer species in a riparian environment. Functional Ecology. 17(2): 170-177. [81988]
192. Kartesz, John T. 1999. A synonymized checklist and atlas with biological attributes for the vascular flora of the United States, Canada, and Greenland. 1st ed. In: Kartesz, John T.; Meacham, Christopher A. Synthesis of the North American flora (Windows Version 1.0), [CD-ROM]. Chapel Hill, NC: North Carolina Botanical Garden (Producer). In cooperation with: The Nature Conservancy; U.S. Department of Agriculture, Natural Resources Conservation Service; U.S. Department of the Interior, Fish and Wildlife Service. [36715]
193. Kartesz, John Thomas. 1988. A flora of Nevada. Reno, NV: University of Nevada. 1729 p. Dissertation. [In 2 volumes]. [42426]
194. Kauffman, J. Boone; Krueger, W. C.; Vavra, M. 1985. Ecology and plant communities of the riparian areas associated with Catherine Creek in northeastern Oregon. Tech. Bull. 147. Corvallis, OR: Oregon State University, Agricultural Experiment Station. 35 p. [6174]
195. Kearney, Thomas H.; Peebles, Robert H.; Howell, John Thomas; McClintock, Elizabeth. 1960. Arizona flora. 2nd ed. Berkeley, CA: University of California Press. 1085 p. [6563]
196. Knighton, M. Dean. 1981. Growth response of speckled alder and willow to depth of flooding. Res. Pap. NC-198. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 6 p. [14328]
197. Kobziar, Leda N.; McBride, Joe R. 2006. Wildfire burn patterns and riparian vegetation response along two northern Sierra Nevada streams. Forest Ecology and Management. 222(1-3): 254-265. [79123]
198. Komarkova, Vera. 1986. Habitat types on selected parts of the Gunnison and Uncompahgre National Forests. Final report: Contract No. 28-K2-234. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 270 p. [1369]
199. Komarkova, Vera; Alexander, Robert R.; Johnston, Barry C. 1988. Forest vegetation of the Gunnison and parts of the Uncompahgre National Forests: a preliminary habitat type classification. Gen. Tech. Rep. RM-163. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 65 p. [5798]
200. Kovalchik, Bernard L. 1987. Riparian zone associations: Deschutes, Ochoco, Fremont, and Winema National Forests. R6 ECOL TP-279-87. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 171 p. [9632]
201. Kovalchik, Bernard L.; Chitwood, Lawrence A. 1990. Use of geomorphology in the classification of riparian plant associations in mountainous landscapes of central Oregon, U.S.A. Forest Ecology and Management. 33/34: 405-418. [6830]
202. Kovalchik, Bernard L.; Clausnitzer, Rodrick R. 2004. Classification and management of aquatic, riparian, and wetland sites on the national forests of eastern Washington: series description. Gen. Tech. Rep. PNW-GTR-593. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 354 p. [53329]
203. Kovalchik, Bernard L.; Hopkins, William E.; Brunsfeld, Steven J. 1988. Major indicator shrubs and herbs in riparian zones on national forests of central Oregon. R6-ECOL-TP-005-88. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 159 p. [8995]
204. Krajina, V. J.; Klinka, K.; Worrall, J. 1982. Distribution and ecological characteristics of trees and shrubs of British Columbia. Vancouver, BC: University of British Columbia. 131 p. [6728]
205. Kudish, Michael. 1992. Adirondack upland flora: an ecological perspective. Saranac, NY: The Chauncy Press. 320 p. [19376]
206. Kurkowski, Thomas A.; Mann, Daniel H.; Rupp, T. Scott; Verbyla, David L. 2008. Relative importance of different secondary successional pathways in an Alaskan boreal forest. Canadian Journal of Forest Research. 38(7): 1911-1923. [77039]
207. Kurmis, Vilis; Webb, Sara L.; Merriam, Lawrence C., Jr. 1986. Plant communities of Voyageurs National Park, Minnesota, U.S.A. Canadian Journal of Botany. 64: 531-540. [16088]
208. Lackschewitz, Klaus. 1991. Vascular plants of west-central Montana--identification guidebook. Gen. Tech. Rep. INT-227. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 648 p. [13798]
209. Lamb, Eric G.; Mallik, Azim U.; Mackereth, Robert W. 2003. The early impact of adjacent clearcutting and forest fire on riparian zone vegetation in northwestern Ontario. Forest Ecology and Management. 177: 529-538. [43861]
210. LANDFIRE Rapid Assessment. 2005. Potential Natural Vegetation Group (PNVG) R1PIPO--ponderosa pine, [Online]. In: Rapid assessment reference condition models. In: LANDFIRE. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory; U.S. Geological Survey; The Nature Conservancy (Producers). Available: https://www.fs.usda.gov /database/feis/pdfs/PNVGs/California/R1PIPO.pdf [2011, May 26]. [82685]
211. LANDFIRE Rapid Assessment. 2005. Potential Natural Vegetation Group (PNVG) R7NESF--northeast spruce-fir forest, [Online]. In: Rapid assessment reference condition models. In: LANDFIRE. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory; U.S. Geological Survey; The Nature Conservancy (Producers). Available: http://www.landfire.gov/zip/NE/R7NESF.pdf [2008, October 24]. [71581]
212. LANDFIRE Rapid Assessment. 2005. Potential Natural Vegetation Group (PNVG) R7NHSP--northern hardwoods-spruce, [Online]. In: Rapid assessment reference condition models. In: LANDFIRE. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory; U.S. Geological Survey; The Nature Conservancy (Producers). Available: http://www.landfire.gov/zip/Ne/R7NHSP.pdf [2011, May 26]. [71582]
213. LANDFIRE Rapid Assessment. 2005. Reference condition modeling manual (Version 2.1), [Online]. In: LANDFIRE. Cooperative Agreement 04-CA-11132543-189. Boulder, CO: The Nature Conservancy; U.S. Department of Agriculture, Forest Service; U.S. Department of the Interior (Producers). 72 p. Available: http://www.landfire.gov/downloadfile.php?file=RA_Modeling_Manual_v2_1.pdf [2007, May 24]. [66741]
214. LANDFIRE Rapid Assessment. 2007. Rapid assessment reference condition models, [Online]. In: LANDFIRE. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Lab; U.S. Geological Survey; The Nature Conservancy (Producers). Available: http://www.landfire.gov/models_EW.php [2008, April 18] [66533]
215. Lanner, Ronald M. 1983. Trees of the Great Basin: A natural history. Reno, NV: University of Nevada Press. 215 p. [1401]
216. Larsen, C. P. S.; MacDonald, G. M. 1998. An 840-year record of fire and vegetation in a boreal white spruce forest. Ecology. 79(1): 106-118. [28521]
217. Larsen, J. A. 1929. Fires and forest succession in the Bitterroot Mountains of northern Idaho. Ecology. 10: 67-76. [6990]
218. Larson, Frederic R. 1994. SRM 901: Alder. In: Shiflet, Thomas N., ed. Rangeland cover types of the United States. Denver, CO: Society for Range Management: 125-126. [67490]
219. Larson, Gary E. 1993. Aquatic and wetland vascular plants of the Northern Great Plains. Gen. Tech. Rep. RM-238. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 681 p. Available online: http://www.npwrc.usgs.gov/resource/plants/vascplnt/index.htm [2011, June 29]. [22534]
220. Larson, Milo; Moir, W. H. 1987. Forest and woodland habitat types (plant associations) of northern New Mexico and northern Arizona. 2nd ed. Albuquerque, NM: U.S. Department of Agriculture, Forest Service, Southwestern Region. 90 p. [8947]
221. Latting, June, ed. 1976. Symposium proceedings--plant communities of southern California. Special Publication No. 2. Berkeley, CA: California Native Plant Society. 164 p. [1414]
222. Lawrence, William H. 1954. Michigan beaver populations as influenced by fire and logging. Ann Arbor, MI: University of Michigan. 219 p. Dissertation. [16480]
223. Leach, Howard R.; Hiehle, Jack L. 1956. Food habits of the Tehama deer herd. California Fish and Game. 43: 161-178. [6874]
224. Lee, Philip. 2004. The impact of burn intensity from wildfires on seed and vegetative banks, and emergent understory in aspen-dominated boreal forests. Canadian Journal of Botany. 82(10): 1468-1480. [51462]
225. Lee, Philip; Sturgess, Kelly. 2001. The effects of logs, stumps, and root throws on understory communities within 28-year-old aspen-dominated boreal forests. Canadian Journal of Botany. 79: 905-916. [38852]
226. Lee, Philip; Sturgess, Kelly. 2002. Assemblages of vascular plants on logs and stumps within 28-year-old aspen-dominated boreal forests. In: Laudenslayer, William F., Jr.; Shea, Patrick J.; Valentine, Bradley E.; Weatherspoon, C. Phillip; Lisle, Thomas E., tech. coords. Proceedings of the symposium on the ecology and management of dead wood in western forests; 1999 November 2-4; Reno, NV. Gen. Tech. Rep. PSW-GTR-181. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 369-380. [44371]
227. Lindsey, Alton A. 1952. Vegetation of the ancient beaches above Great Bear and Great Slave Lakes. Ecology. 33(4): 535-549. [65095]
228. Little, Elbert L., Jr. 1950. Southwestern trees: A guide to the native species of New Mexico and Arizona. Agric. Handb. No. 9. Washington, DC: U.S. Department of Agriculture, Forest Service. 109 p. [20317]
229. Little, Elbert L., Jr. 1976. Atlas of United States trees. Volume 3. Minor western hardwoods. Misc. Publ. 1314. Washington, DC: U.S. Department of Agriculture, Forest Service. 13 p. 290 maps. [10430]
230. Little, Elbert L., Jr. 1977. Atlas of United States trees. Volume 4. Minor eastern hardwoods. Misc. Pub. No. 1342. Washington, DC: U.S. Department of Agriculture, Forest Service. 17 p. [21683]
231. Little, Elbert L., Jr. 1979. Checklist of United States trees (native and naturalized). Agric. Handb. 541. Washington, DC: U.S. Department of Agriculture, Forest Service. 375 p. [2952]
232. Loope, Walter L. 1991. Interrelationships of fire history, land use history, and landscape pattern within Pictured Rocks National Seashore, Michigan. The Canadian Field-Naturalist. 105(1): 18-28. [5950]
233. Lubinski, Sara; Hop, Kevin; Gawler, Susan. 2003. U.S. Geological Survey-National Park Service Vegetation Mapping Program: Acadia National Park, Maine. Final report. [Revised edition]. La Crosse, WI: U.S. Department of the Interior, U.S. Geological Survey, Upper Midwest Environmental Studies Center. 50 p. [+ appendices]. Available online: http://biology.usgs.gov/npsveg/acad/acadrpt.pdf. [79619]
234. Lutz, H. J. 1956. Ecological effects of forest fires in the interior of Alaska. Tech. Bull. No. 1133. Washington, DC: U.S. Department of Agriculture, Forest Service. 121 p. [7653]
235. Lyford, Walter H., Jr. 1943. The palatability of freshly fallen forest tree leaves to millipedes. Ecology. 24(2): 252-261. [64424]
236. MacLean, D. W. 1960. Some aspects of the aspen-birch-spruce-fir type in Ontario. Tech. Note No. 94. Ottawa, Canada: Department of Forestry, Forest Research Division. 24 p. [14608]
237. Magee, Dennis W.; Ahles, Harry E. 2007. Flora of the Northeast: A manual of the vascular flora of New England and adjacent New York. 2nd ed. Amherst, MA: University of Massachusetts Press. 1214 p. [74293]
238. Majcen, Zoran; Gagnon, Gilles; Benzie, John. 1980. Jack pine. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 8-9. [49850]
239. Manning, Mary E.; Padgett, Wayne G. 1989. Preliminary riparian community type classification for Nevada. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Region. Preliminary draft. On file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 135 p. [11531]
240. Martin, Alexander C.; Zim, Herbert S.; Nelson, Arnold L. 1951. American wildlife and plants. New York: McGraw-Hill Book Company. 500 p. [4021]
241. Martin, William C.; Hutchins, Charles R. 1981. A flora of New Mexico. Volume 2. Germany: J. Cramer. 2589 p. [37176]
242. Mason, Herbert L. 1957. A flora of the marshes of California. Berkeley, CA: University of California Press. 878 p. [16905]
243. McArthur, E. Durant; Sanderson, Stewart C. 1991. Great Sand Dunes National Monument vegetation patterns. In: Plumb, Glenn E., ed. University of Wyoming: National Park Service Research Center 15th annual report 1991. Laramie, WY: University of Wyoming: 185-189. [29955]
244. McBride, Joe R. 1994. SRM 203: Riparian woodland. In: Shiflet, Thomas N., ed. Rangeland cover types of the United States. Denver, CO: Society for Range Management: 13-14. [66662]
245. McCain, Cindy; Christy, John A. 2005. Field guide to riparian plant communities in northwestern Oregon. Tech. Pap. R6-NR-ECOL-TP-01-05. [Portland, OR]: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 357 p. [63114]
246. McDermott, R. E. 1953. Light as a factor in the germination of some bottomland hardwood seeds. Journal of Forestry. 51: 203-204. [168]
247. McGinnies, William G. 1972. North America. In: McKell, Cyrus M.; Blaisdell, James P.; Goodin, Joe R., tech. eds. Wildland shrubs--their biology and utilization: An international symposium: Proceedings; 1971 July; Logan, UT. Gen. Tech. Rep. INT-1. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 55-66. [22750]
248. McRae, Douglas J. 1986. Guidelines for prescribed burning in Ontario's Clay Belt boreal mixedwood slash. In: Koonce, Andrea L., ed. Prescribed burning in the Midwest: state-of-the-art: Proceedings of a symposium; 1986 March 3-6; Stevens Point, WI. Stevens Point, WI: University of Wisconsin, College of Natural Resources, Fire Science Center: 28-33. [16265]
249. Mead, Bert R. 1994. SRM 917: Tall shrub swamp. In: Shiflet, Thomas N., ed. Rangeland cover types of the United States. Denver, CO: Society for Range Management: 137-138. [67507]
250. Meades, W. J.; Moores, L. 1989. Forest site classification manual: A field guide to the Damman forest types of Newfoundland. Forest Resources Development Agreement FRDA Report 003. St. Johns, NF: Environment Canada. 295 p. [49220]
251. Meilleur, Alain; Bouchard, Andre; Bergeron, Yves. 1994. The relation between geomorphology and forest community types of the Haut-Saint-Laurent, Quebec. Vegetatio. 111: 173-192. [23539]
252. Mendall, Howard L.; Aldous, Clarence M. 1943. The ecology and management of the American woodcock. Orono, ME: Maine Cooperative Wildlife Research Unit. 201 p. [73807]
253. Merritt, David M.; Wohl, Ellen E. 2006. Plant dispersal along rivers fragmented by dams. River Research and Applications. 22: 1-26. [61821]
254. Mirov, N. T.; Kraebel, C. J. 1937. Collecting and propagating the seeds of California wild plants. Res. Note No. 18. Berkeley, CA: U.S. Department of Agriculture, Forest Service, California Forest and Range Experiment Station. 27 p. [9787]
255. Mohlenbrock, Robert H. 1986. Guide to the vascular flora of Illinois. [Revised edition]. Carbondale, IL: Southern Illinois University Press. 507 p. [17383]
256. Monsen, Stephen B.; Stevens, Richard; Shaw, Nancy L. 2004. Shrubs of other families. In: Monsen, Stephen B.; Stevens, Richard; Shaw, Nancy L., comps. Restoring western ranges and wildlands. Gen. Tech. Rep. RMRS-GTR-136-vol-2. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 598-698. [52846]
257. Morin, Hubert; Payette, Serge. 1988. Buried seed populations in the montane, subalpine, and alpine belts of Mont Jacques-Cartier, Quebec. Canadian Journal of Botany. 66: 101-107. [6376]
258. Morris, L. A.; Mollitor, A. V.; Johnson, K. J.; Leaf, A. L. 1979. Forest management of floodplain sites in the northeastern United States. In: Johnson, R. Roy; McCormick, J. Frank, technical coordinators. Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium; 1978 December 11-13; Callaway Gardens, GA. Gen. Tech. Rep. WO-12. Washington, DC: U.S. Department of Agriculture, Forest Service: 236-242. [4364]
259. Morris, Melvin S.; Schmautz, Jack E.; Stickney, Peter F. 1962. Winter field key to the native shrubs of Montana. Bulletin No. 23. Missoula, MT: Montana State University, Montana Forest and Conservation Experiment Station. 70 p. [17063]
260. Moseley, Robert K. 1998. Riparian and wetland community inventory of 14 reference areas in southwestern Idaho. Technical Bulletin No. 98-5. Boise, Idaho: U.S. Department of the Interior, Bureau of Land Management, Boise State Office. 52 p. [75569]
261. Mozingo, Hugh N. 1987. Shrubs of the Great Basin: A natural history. Reno, NV: University of Nevada Press. 342 p. [1702]
262. Muldavin, Esteban; Durkin, Paula; Bradley, Mike; Stuever, Mary; Mehlhop, Patricia. 2000. Handbook of wetland vegetation communities of New Mexico. Volume 1: classification and community descriptions. Albuquerque, NM: University of New Mexico, Biology Department; New Mexico Natural Heritage Program. 172 p. [+ appendices]. [45517]
263. Munda, P.; Pater, M. 2001. Commercial sources of conservation plant materials, [Online]. Tucson, AZ: U.S. Department of Agriculture, Natural Resources Conservation Service, Tucson Plant Materials Center (Producer). Available: http://plant-materials.nrcs.usda.gov/pubs/azpmsarseedlist0501.pdf [2003, August 25]. [44989]
264. Nevada Department of Conservation and Natural Resources, Nevada Natural Heritage Program. 2003. National vegetation classification for Nevada, [Online]. Carson City, NV: Nevada Department of Conservation and Natural Resources, Nevada Natural Heritage Program (Producer). 15 p. Available: http://heritage.nv.gov/ecology/nv_nvc.htm [2005, November 3]. [55021]
265. Newton, Michael; Cole, Elizabeth C.; Lautenschlager, R. A.; White, Diane E.; McCormack, M. L. 1989. Browse availability after conifer release in Maine's spruce-fir forests. Journal of Wildlife Management. 53(3): 643-649. [8401]
266. Niemi, Gerald J.; Pfannmuller, Lee. 1979. Avian communities: approaches to describing their habitat associations. In: DeGraaf, Richard M.; Evans, Keith E., compilers. Proceedings of the workshop: Management of northcentral and northeastern forests for nongame birds; 1979 January 23-25; Minneapolis, MN. Gen. Tech. Rep. NC-51. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station: 154-178. [18086]
267. Ohmann, Lewis F.; Knighton, M. Dean; McRoberts, Ronald. 1990. Influence of flooding duration on the biomass growth of alder and willow. Res. Pap. NC-292. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest and Range Experiment Station. 5 p. [13179]
268. Olson, Diana L.; Agee, James K. 2005. Historical fires in Douglas-fir dominated riparian forests of the southern Cascades, Oregon. Fire Ecology. 1(1): 50-74. [65420]
269. Olson, R. A.; Gerhart, W. A. 1982. A physical and biological characterization of riparian habitat and its importance to wildlife in Wyoming. Cheyenne, WY: Wyoming Game and Fish Department. 188 p. [6755]
270. Oosting, Henry J. 1942. An ecological analysis of the plant communities of the Piedmont, North Carolina. The American Midland Naturalist. 28: 1-126. [50588]
271. Orloci, Laszlo; Stanek, Walter. 1979. Vegetation survey of the Alaska Highway, Yukon Territory: types and gradients. Vegetatio. 41(1): 1-56. [12748]
272. Ostler, William K.; Buchanan, Hayle. 1973. The effects of slope exposure on plant communities. Proceedings, Utah Academy of Sciences, Arts and Letters. 50(2): 58-63. [42486]
273. Ostrom, Arnold J. 1983. Tree and shrub biomass estimates for Michigan, 1980. Res. Note NC-302. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 7 p. [8162]
274. Padgett, Wayne G.; Youngblood, Andrew P.; Winward, Alma H. 1989. Riparian community type classification of Utah and southeastern Idaho. R4-Ecol-89-01. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Region. 191 p. [11360]
275. Parker, George R.; Schneider, G. 1974. Structure and edaphic factors of an alder swamp in northern Michigan. Canadian Journal of Forestry. 4: 499-508. [15113]
276. Pase, Charles P.; Layser, Earle F. 1977. Classification of riparian habitat in the Southwest. In: Johnson, Roy; Jones, Dale A., technical coordinators. Importance, preservation and management of riparian habitat: A symposium; 1977 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 5-9. Available from: NTIS, Springfield, VA 22151; PB-274 582. [5333]
277. Patterson, Karen D. 2008. Vegetation classification and mapping at Appomattox Court House National Historical Park, Virginia. Technical Report NPS/NER/NRTR--2008/125. Philadelphia, PA: U.S. Department of the Interior, National Park Service, Northeast Region. 261 p. [78026]
278. Patterson, Karen D. 2008. Vegetation classification and mapping at Petersburg National Battlefield, Virginia. Technical Report NPS/NER/NRTR--2008/127. Philadelphia, PA: U.S. Department of the Interior, National Park Service, Northeast Region. 235 p. [79673]
279. Patterson, Karen D. 2008. Vegetation classification and mapping at Richmond National Battlefield Park, Virginia. Technical Report NPS/NER/NRTR--2008/128. Philadelphia, PA: U.S. Department of the Interior, National Park Service, Northeast Region. 244 p. [79674]
280. Peet, Robert K. 1981. Forest vegetation of the Colorado Front Range: composition and dynamics. Vegetatio. 45: 3-75; 1981. [1867]
281. Peet, Robert K. 1988. Forests of the Rocky Mountains. In: Barbour, Michael G.; Billings, William Dwight, eds. North American terrestrial vegetation. New York: Cambridge University Press: 63-101. [6714]
282. Perles, Stephanie J.; Podniesinski, Gregory S.; Eastman, E.; Sneddon, Lesley A.; Gawler, Sue C. 2007. Classification and mapping of vegetation and fire fuel models at Delaware Water Gap National Recreation Area: Volume 1 of 2, [Online]. Technical Report NPS/NER/NRTR2007/076. Philadelphia, PA: U.S. Department of the Interior, National Park Service, Northeast Region, Natural Resource Stewardship and Science. (Producer) 187 p. Available: http://www.nps.gov/nero/science/FINAL/DEWA_veg_map/DEWA_veg_map.htm [2010, March 3]. [78999]
283. Perles, Stephanie J.; Podniesinski, Gregory S.; Eastman, E.; Sneddon, Lesley A.; Gawler, Sue C. 2007. Classification and mapping of vegetation and fire fuel models at Delaware Water Gap National Recreation Area: Volume 2 of 2--Appendix G, [Online]. Technical Report NPS/NER/NRTR--2007/076. Philadelphia, PA: U.S. Department of the Interior, National Park Service, Northeast Region, Natural Resource Stewardship and Science (Producer). 400 p. Available: http://www.nps.gov/nero/science/FINAL/DEWA_veg_map/DEWA_veg_map.htm [2010, March 3]. [79090]
284. Peterson, Eric B. 2008. International vegetation classification alliances and associations occurring in Nevada with proposed additions. Carson City, NV: Nevada Natural Heritage Program. 347 p. Available online: http://heritage.nv.gov/reports/ivclist.pdf [2009, December 21]. [77864]
285. Pettit, Neil E.; Naiman, Robert J. 2007. Fire in the riparian zone: characteristics and ecological consequences. Ecosystems. 10: 673-687. [68289]
286. Pierce, John; Johnson, Janet. 1986. Wetland community type classification for west-central Montana. [Review draft]. Missoula, MT: U.S. Department of Agriculture, Forest Service, Northern Region, Ecosystem Management Program. 158 p. On file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [7436]
287. Pilliod, David S.; Arkle, Robert S.; Yoshimura, Michael A. 2008. Post-fire recovery of stream amphibians, benthic macroinvertebrates, and riparian vegetation in a federally designated wilderness. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. Final Report: Joint venture agreement 06-JV011221644-175. 19 p. In cooperation with: Cal Poly Corporation of the California Polytechnic State University. [64834]
288. Pinder, J. E., III; Kroh, G. C.; White, J. D.; May, A. M. Basham. 1997. The relationships between vegetation type and topography in Lassen Volcanic National Park. Plant Ecology. 131(1): 17-29. [40062]
289. Pollett, Frederick C. 1972. Classification of peatlands in Newfoundland. Proceedings, 4th International Peat Congress. 1: 101-110. [15403]
290. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press. 632 p. [2843]
291. Reardon, Jim Douglas. 1950. Woodcock utilization of improved covers in eastern Maine. Orono, ME: University of Maine. 83 p. Thesis. [73769]
292. Reich, Peter B.; Bakken, Peter; Carlson, Daren; Frelich, Lee E.; Friedman, Steve K.; Grigal, David F. 2001. Influence of logging, fire, and forest type on biodiversity and productivity in southern boreal forests. Ecology. 82(10): 2731-2748. [45136]
293. Reschke, Carol. 1990. Ecological communities of New York State. Latham, NY: New York State Department of Environmental Conservation, Natural Heritage Program. 96 p. [21441]
294. Reuter, D. Dayton. 1986. Effects of prescribed burning, cutting and torching on shrubs in a sedge meadow wetland. In: Koonce, Andrea L., ed. Prescribed burning in the Midwest: state-of-the-art: Proceedings of a symposium; 1986 March 3-6; Stevens Point, WI. Stevens Point, WI: University of Wisconsin, College of Natural Resources, Fire Science Center: 108-115. [16278]
295. Roland, A. E.; Smith, E. C. 1969. The flora of Nova Scotia. Halifax, NS: Nova Scotia Museum. 746 p. [13158]
296. Royal Botanic Garden Edinburgh. 2011. Flora Europaea, [Online]. Edinburgh, UK: Royal Botanic Garden Edinburgh (Producer). Available: http://rbg-web2.rbge.org.uk/FE/fe.html. [41088]
297. Russell, William H.; McBride, Joe R.; Carnell, Ky. 2003. Influence of environmental factors on the regeneration of hardwood species on three streams in the Sierra Nevada. Madrono. 50(1): 21-27. [44804]
298. Rytter, L.; Sennerby-Forsse, L.; Alriksson, A. 2000. Natural regeneration of grey alder (Alnus incana [L.] Moench.) stands after harvest. Journal of Sustainable Forestry. 10(3/4): 287-294. [36403]
299. Sawyer, John O.; Thornburgh, Dale A. 1977. Montane and subalpine vegetation of the Klamath Mountains. In: Barbour, Michael G.; Major, Jack, eds. Terrestrial vegetation of California. New York: John Wiley & Sons: 699-732. [685]
300. Schalin, Ilmari. 1967. Germination analysis of Alnus incana (L.) and Alnus glutinosa (L.) seeds. Oikos. 18(2): 253-260. [81987]
301. Schneider, Gregory J. 1994. The sedge (Carex) flora of Ohio fens. In: Wickett, Robert G.; Lewis, Patricia Dolan; Woodliffe, Allen; Pratt, Paul, eds. Proceedings of the 13th North American prairie conference: spirit of the land, our prairie legacy; 1992 August 6-9; Windsor, ON. Windsor, ON: Department of Parks and Recreation: 147-155. [24686]
302. Schneider, Rick E.; Faber-Langendoen, Don; Crawford, Rex C.; Weakley, Alan S. 1997. The status of biodiversity in the Great Plains: Great Plains vegetation classification--Supplemental document 1, [Online]. [Cooperative Agreement # X 007803-01-3]. In: Ostlie, Wayne R.; Schneider, Rick E.; Aldrich, Janette Marie; Faust, Thomas M.; McKim, Robert L. B.; Chaplin, Stephen J., comps. The status of biodiversity in the Great Plains. Arlington, VA: The Nature Conservancy, Great Plains Program (Producer). 75 p. Available: http://conserveonline.org/docs/2005/02/greatplains_vegclass_97.pdf [2006, May 16]. [62020]
303. Schwintzer, Christa R.; Williams, Gary. 1974. Vegetation changes in a small Michigan bog from 1917 to 1972. The American Midland Naturalist. 92(2): 447-459. [66032]
304. Scoggan, H. J. 1978. The flora of Canada. Part 3: Dicotyledoneae (Saururaceae to Violaceae). National Museum of Natural Sciences: Publications in Botany, No. 7(3). Ottawa: National Museums of Canada. 1115 p. [75493]
305. Scott, James A. 1986. The butterflies of North America: A natural history and field guide. Standord, CA: Stanford University Press. 584 p. [82601]
306. Scotter, George W. 1963. Effects of forest fires on soil properties in northern Saskatchewan. Forestry Chronicle. 39(4): 412-421. [13605]
307. Shaffer, Kevin E.; Laudenslayer, William F., Jr. 2006. Fire and animal interactions. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 118-144. [65536]
308. Shafi, M. I.; Yarranton, G. A. 1973. Vegetational heterogeneity during a secondary (postfire) succession. Canadian Journal of Botany. 51: 73-90. [15191]
309. Shaw, Nancy L. 2004. Production and use of planting stock. In: Monsen, Stephen B.; Stevens, Richard; Shaw, Nancy L., comps. Restoring western ranges and wildlands. Gen. Tech. Rep. RMRS-GTR-136-vol. 3. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 745-768. [42197]
310. Skinner, Carl N. 2002. Fire history in riparian reserves of the Klamath Mountains. In: Sugihara, Neil G.; Morales, Maria; Morales, Tony, eds. Fire in California ecosystems: integrating ecology, prevention and management: Proceedings of the symposium; 1997 November 17-20; San Diego, CA. Misc. Pub. No. 1. [Davis, CA]: Association for Fire Ecology: 164-169. [46203]
311. Skinner, Carl N.; Taylor, Alan H.; Agee, James K. 2006. Klamath Mountains bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 170-194. [65539]
312. Smith, Jane Kapler; Fischer, William C. 1997. Fire ecology of the forest habitat types of northern Idaho. Gen. Tech. Rep. INT-GTR-363. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 142 p. [27992]
313. Smith, W. Brad. 1986. Biomass yields for small trees, shrubs, and herbs in northern Lake States forests. Res. Pap. NC-277. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 11 p. [8159]
314. Smith, W. Brad; Brand, Gary J. 1983. Allometric biomass equations for 98 species of herbs, shrubs, and small trees. Res. Note NC-299. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 8 p. [20785]
315. Snyder, Gordon G.; Haupt, Harold F.; Belt, George H., Jr. 1975. Clearcutting and burning slash alter quality of stream water in northern Idaho. Res. Pap. INT-168. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 34 p. [8615]
316. Stanek, W.; Alexander, K.; Simmons, C. S. 1981. Reconnaissance of vegetation and soils along the Dempster Highway, Yukon Territory: I. Vegetation types. BC-X-217. Victoria, BC: Environment Canada, Canadian Forestry Service, Pacific Forest Research Centre. 32 p. [16526]
317. Stark, Kaeli E.; Arsenault, Andre; Bradfield, Gary E. 2006. Soil seed banks and plant community assembly following disturbance by fire and logging in interior Douglas-fir forests of south-central British Columbia. Canadian Journal of Botany. 84(10): 1548-1560. [65962]
318. Stephens, H. A. 1973. Woody plants of the north Central Plains. Lawrence, KS: The University Press of Kansas. 530 p. [3804]
319. Stickney, Peter F. 1989. Seral origin of species comprising secondary plant succession in Northern Rocky Mountain forests. FEIS workshop: Postfire regeneration. Unpublished draft on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 10 p. [20090]
320. Stine, Scott; Gaines, David; Vorster, Peter. 1984. Destruction of riparian systems due to water development in the Mono Lake watershed. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of the conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 528-533. [5857]
321. Stocks, B. J.; McRae, D. J.; Lynham, T. J.; Hartley, G. R. 1990. A photo-series for assessing fuels in natural forest stands in northern Ontario. COFRDA Report 3304. Sault Ste. Marie, ON: Forestry Canada, Ontario Region. 161 p. [50797]
322. Suring, Lowell H.; Rowland, Mary M.; Wisdom, Michael J.; Schueck, Linda; Meinke, Cara W. 2005. Vegetation communities. In: Wisdom, Michael J.; Rowland, Mary M.; Suring, Lowell H., eds. Habitat threats in the sagebrush ecosystem: methods of regional assessment and applications in the Great Basin. Lawrence, KS: Alliance Communications Group: 94-113. [67402]
323. Swain, Albert M. 1973. A history of fire and vegetation in northeastern Minnesota as recorded in lake sediments. Quaternary Research. 3(3): 383-396. [38931]
324. Swain, Albert M. 1980. Landscape patterns and forest history in the Boundary Waters Canoe Area, Minnesota: a pollen study from Hug Lake. Ecology. 61(4): 747-754. [1895]
325. Swanson, David K.; Schmitt, Craig L.; Shirley, Diane M.; Erickson, Vicky; Schuetz, Kenneth J.; Tatum, Michael L.; Powell, David C. 2010. Aspen biology, community classification, and management in the Blue Mountains. Gen. Tech. Rep. PNW-GTR-806. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 117 p. [81634]
326. Szaro, Robert C. 1989. Riparian forest and scrubland community types of Arizona and New Mexico. Desert Plants. 9(3-4): 70-138. [604]
327. Taverna, Kristin. 2008. Vegetation classification and mapping at Fredericksburg and Spotsylvania National Military Park. Technical Report NPS/NER/NRTR--2008/126. Philadelphia, PA: U.S. Department of the Interior, National Park Service, Northeast Region. 277 p. [79671]
328. Telfer, E. S. 1972. Understory biomass in five forest types in southwestern Nova Scotia. Canadian Journal of Botany. 50: 1263-1267. [13933]
329. Ter-Mikaelian, Michael T.; Korzukhin, Michael D. 1997. Biomass equations for sixty-five North American tree species. Forest Ecology and Management. 97: 1-24. [28034]
330. The Nature Conservancy. 1999. Classification of the vegetation of Isle Royale National Park. USGS-NPS Vegetation Mapping Program. Minneapolis, MN: The Nature Conservancy; Arlington, VA: The Nature Conservancy. 140 p. Available online: http://biology.usgs.gov/npsveg/isro/isrorpt.pdf [2007, October 3]. [68269]
331. Tilton, Donald L.; Bernard, John M. 1975. Primary productivity and biomass distribution in an alder shrub ecosystem. The American Midland Naturalist. 94(1): 251-256. [81985]
332. Timoney, Kevin P.; Peterson, George; Wein, Ross. 1997. Vegetation development of boreal riparian plant communities after fire, flooding, and logging, Peace River, Canada. Forest Ecology and Management. 93: 101-120. [27427]
333. Tuhy, Joel S.; Jensen, Sherman. 1982. Riparian classification for the Upper Salmon/Middle Fork Salmon River drainages, Idaho. Final report. Smithfield, UT: White Horse Associates. Unpublished report on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station Fire Sciences Laboratory, Missoula, MT. 153 p. [8380]
334. U.S. Department of Agriculture, Forest Service. 1937. Range plant handbook. Washington, DC. 532 p. [2387]
335. U.S. Department of Agriculture, Natural Resources Conservation Service. 2011. PLANTS Database, [Online]. Available: https://plants.usda.gov /. [34262]
336. U.S. Department of the Interior, Fish and Wildlife Service, Division of Endangered Species. 2011. Threatened and endangered animals and plants, [Online]. Available: http://www.fws.gov/endangered/wildlife.html. [62042]
337. Ugolini, F. C. 1968. Soil development and alder invasion in a recently deglaciated area of Glacier Bay, Alaska. In: Trappe, J. M.; Franklin, J. F.; Tarrant, R. F.; Hansen, G. M., eds. Biology of alder: Proceedings of a symposium; 1967 April 14-15; Pullman, WA. Portland, OR: U. S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: 115-140. [6211]
338. Van Cleave, Keith; Viereck, Leslie A. 1972. Distribution of selected chemical elements in even-aged alder (Alnus) ecosystems near Fairbanks, Alaska. Arctic and Alpine Research. 4(3): 239-255. [82078]
339. Van Cleave, Keith; Viereck, Leslie A.; Schlentner, Robert L. 1971. Accumulation of nitrogen in alder (Alnus) ecosystems near Fairbanks, Alaska. Arctic and Alpine Research. 3(2): 101-114. [81982]
340. Van de Water, Kip; North, Malcolm. 2010. Fire history of coniferous riparian forests in the Sierra Nevada. Forest Ecology and Management. 260(3): 383-395. [82640]
341. Vandermast, D. B.; Van Lear, D. H. 2002. Riparian vegetation in the southern Appalachian Mountains (USA) following chestnut blight. Forest Ecology and Management. 155: 97-106. [40782]
342. Vestal, Arthur G. 1917. Foothills vegetation in the Colorado Front Range. Botanical Gazette. 64(5): 353-385. [64489]
343. Viereck, L. A.; Dyrness, C. T.; Batten, A. R.; Wenzlick, K. J. 1992. The Alaska vegetation classification. Gen. Tech. Rep. PNW-GTR-286. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 278 p. [2431]
344. Viereck, Leslie A. 1973. Wildfire in the taiga of Alaska. Quaternary Research. 3: 465-495. [7247]
345. Viereck, Leslie A.; Little, Elbert L., Jr. 1972. Alaska trees and shrubs. Agric. Handb. 410. Washington, DC: U.S. Department of Agriculture, Forest Service. 265 p. [6884]
346. Viereck, Leslie A.; Schandelmeier, Linda A. 1980. Effects of fire in Alaska and adjacent Canada--a literature review. BLM-Alaska Tech. Rep. 6; BLM/AK/TR-80/06. Anchorage, AK: U.S. Department of the Interior, Bureau of Land Management, Alaska State Office. 124 p. [28862]
347. Vincent, A. B. 1964. Growth and numbers of speckled alder following logging of black spruce peatlands. Forestry Chronicle. 40: 515-518. [14318]
348. Vines, Robert A. 1960. Trees, shrubs, and woody vines of the Southwest. Austin, TX: University of Texas Press. 1104 p. [7707]
349. Voigt, G. K.; Steucek, G. L. 1969. Nitrogen distribution and accretion in an alder ecosystem. Soil Society of America Proceedings. 33: 946-949. [14329]
350. Voss, Edward G. 1985. Michigan flora. Part II. Dicots (Saururaceae--Cornaceae). Bulletin 59. Bloomfield Hills, MI: Cranbrook Institute of Science; Ann Arbor, MI: University of Michigan Herbarium. 724 p. [11472]
351. Walford, Gillian M. 1993. Spatial variation in riparian vegetation along the Animas River, Colorado. Laramie, WY: University of Wyoming. 99 p. Thesis. [53679]
352. Walford, Gillian; Jones, George; Fertig, Walt; Mellman-Brown, Sabine; Houston, Kent E. 2001. Riparian and wetland plant community types of the Shoshone National Forest. Gen. Tech. Rep. RMRS-GTR-85. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station; Cody, WY: U.S. Department of Agriculture, Natural Resources Conservation Service, Cody Conservation District. 122 p. [40599]
353. Walker, Lawrence R.; Chapin, F. Stuart, III. 1986. Physiological controls over seedling growth in primary succession on an Alaskan floodplain. Ecology. 67(6): 1508-1523. [82085]
354. Wasser, C. H.; Hess, Karl. 1982. The habitat types of Region 2--U.S. Forest Service: a synthesis. Final report: Cooperative Agreement No. 16-845-CA. Lakewood, CO: U.S. Department of Agriculture, Forest Service, Region 2. 140 p. [5594]
355. Welsh, Stanley L.; Atwood, N. Duane; Goodrich, Sherel; Higgins, Larry C., eds. 1987. A Utah flora. The Great Basin Naturalist Memoir No. 9. Provo, UT: Brigham Young University. 894 p. [2944]
356. Wharton, Charles H. 1978. The natural environments of Georgia. Atlanta, GA: Georgia Department of Natural Resources. 227 p. [24582]
357. Whitlock, Clair M.; Kastelic, Joseph P.; Johnson, Theodore Alan; Haertel, Paul; Roberts, Alvin Y.; Settles, Raymond L.; Filkins, Elgin E.; Norum, Rodney A.; Kelleyhouse, David G.; Williams, Thomas D. 1979. The Fortymile Interim Fire Management Plan. Fairbanks, AK: U.S. Department of the Interior, Bureau of Land Management, Fairbanks District Office. 97 p. [Prepared by the Fire Subcommittee of the Alaska Land Managers Cooperative Task Force]. [21231]
358. Wiggins, Ira L. 1980. Flora of Baja California. Stanford, CA: Stanford University Press. 1025 p. [21993]
359. Wofford, B. Eugene. 1989. Guide to the vascular plants of the Blue Ridge. Athens, GA: The University of Georgia Press. 384 p. [12908]
360. Wolda, H.; Zweep, A.; Schuitema, K. A. 1971. The role of food in the dynamics of populations of the landsnail Cepaea nemoralis. Oecologia. 7(4): 361-381. [76272]
361. Wright, A. H.; Wright, A. A. 1932. The habitats and composition of the vegetation of Okefinokee Swamp, Georgia. Ecological Monographs. 2(2): 109-232. [17130]
362. Yarie, J.; Viereck, L.; Van Cleve, K.; Dryness, C. T. 1988. The chronosequence as an aid to understanding the long-term consequences of management activities. In: Dyck, W. J.; Mees, C. A., eds. Research strategies for long-term productivity: Proceedings, IEA/BE A3 workshop; 1988 August; Seattle, WA. IEA/BE A3 Report No. 8. Rotorua, New Zealand: Forest Research Institute: 25-38. [17745]
363. Yarie, John; Mead, Bert R. 1988. Twig and foliar biomass estimation equations for major plant species in the Tanana River basin of interior Alaska. Res. Pap. PNW-RP-401. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 20 p. [13487]
364. Young, Robert T. 1907. The forest formations of Boulder County, Colorado. Botanical Gazette. 44(5): 321-352. [64439]
365. Youngblood, Andrew P.; Padgett, Wayne G.; Winward, Alma H. 1985. Riparian community type classification of eastern Idaho - western Wyoming. R4-Ecol-85-01. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Region. 78 p. [2686]
366. Zasada, J. 1986. Natural regeneration of trees and tall shrubs on forest sites in interior Alaska. In: Van Cleve, K.; Chapin, F. S., III; Flanagan, P. W.; Viereck, L. A.; Dyrness, C. T., eds. Forest ecosystems in the Alaska taiga: A synthesis of structure and function. New York: Springer-Verlag: 44-73. [2291]
367. Zasada, John; Norum, Rodney. 1986. Prescribed burning white spruce slash in interior Alaska. Northern Journal of Applied Forestry. 3(1): 16-18. [7881]
368. Zekveld, Corinthe; Markham, John. 2011. Exposure to aphids increases alder growth and nitrogen fixation. Botany. 89: 255-261. [82602]