| FEIS Home Page |
Table of Contents |
 |
|
| Figure 1—Western juniper/mountain big sagebrush woodland in Harney County, Oregon. Photo courtesy of the PRBO Conservation Science Shrubsteppe Monitoring Program. |
Wildfires in mountain big sagebrush steppe communities are typically high-severity, replacement fires that kill or top-kill most of the aboveground vegetation, although variation in fuels, topography, and weather can produce a patchy mosaic of burned and unburned areas. Mountain big sagebrush plants are easily killed by fire; they do not sprout. The soil seed bank and surviving plants in and adjacent to burns are seed sources for postfire establishment. Most wildfires in sagebrush communities occur in summer and early fall before mountain big sagebrush seeds are ripe. Seeds from surviving plants are typically dispersed within 10 feet (3 m) of parent plants in fall and winter. Seed production is variable, but can be high. Fire characteristics that affect the amount of available seeds (e.g., frequency, severity, season, pattern, and size) can have a strong influence on mountain big sagebrush postfire recovery. Postfire seedling establishment rates vary, but high establishment rates can decrease the amount of time for mountain big sagebrush canopy cover to return to prefire or unburned values (i.e., postfire recovery time).
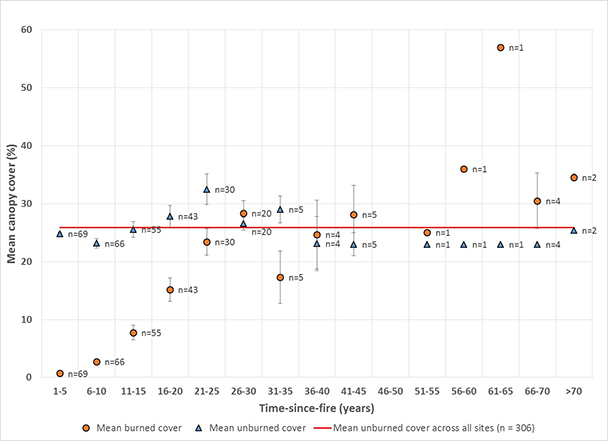
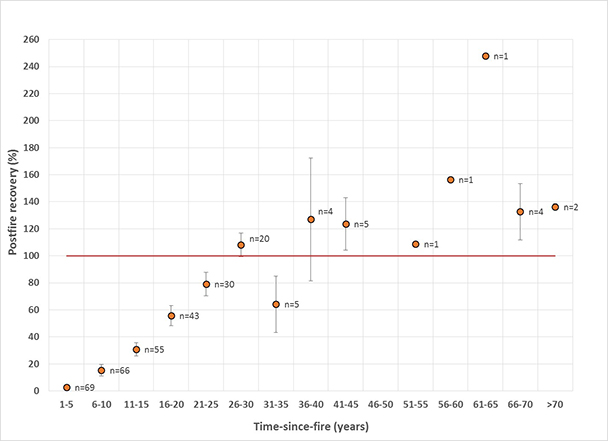
Mountain big sagebrush postfire recovery time is influenced by a number of interacting factors and varies substantially among sites. Our review and analysis of mountain big sagebrush postfire recovery on 306 burned sites examined in 20 studies found that most available data (86%) comes from burns <25 years old, and that few of these sites (9%) had fully recovered. When site-level data was averaged within 5-year bins, sites tended toward full recovery at 28% canopy cover 26 to 30 years after fire; however, not all burns were fully recovered >25 years after fire. Postfire recovery times varied within and among ecoregions. Mountain big sagebrush communities on cold, moist sites are likely to recover faster and be more resilient to fire and more resistant to postfire nonnative annual grass invasion than those on warm, dry sites, although other factors, such as heavy browsing by wild ungulates, can slow postfire recovery regardless of favorable climate and site characteristics.
There are opposing recommendations about the use of fire in mountain big sagebrush communities. In general, prescribed fire is considered an effective management tool only on sites with greater than ~20% cover of native perennial grasses and forbs, abundant mountain big sagebrush, and little to no cheatgrass or other nonnative plants, if the objective is to reduce mountain big sagebrush cover and increase herbaceous plant production. In areas where native perennial plant cover is depleted, seeding after fire helps stabilize soils, speed recovery of sagebrush and other shrubs, and prevent establishment and spread of nonnative species.
FEIS ABBREVIATION:
ARTTRIV
ARTTRI
COMMON NAMES:
mountain big sagebrush
mountain sagebrush
Vasey sage
Vasey's big sagebrush
TAXONOMY:
The scientific name of mountain big sagebrush is Artemisia tridentata subsp. vaseyana (Rydb.) Beetle (Asteraceae) [52,155,204,261,310]. Mountain big sagebrush is one of six subspecies of big sagebrush (Artemisia tridentata Nutt.). Of these, mountain big sagebrush, basin big sagebrush, and Wyoming big sagebrush are the most widely distributed [55,261,311]. In this review, "big sagebrush" refers to the big sagebrush species complex:
Artemisia tridentata Nutt. subsp. parishii (A. Gray) H.M. Hall & Clem., Mojave big sagebrush
Artemisia tridentata Nutt. subsp. spiciformis (Osterh.) Kartesz & Gandhi, snowfield big sagebrush
Artemisia tridentata Nutt. subsp. tridentata, basin big sagebrush
Artemisia tridentata Nutt. subsp. vaseyana (Rydb.) Beetle, mountain big sagebrush
Artemisia tridentata Nutt. subsp. wyomingensis Beetle & Young, Wyoming big sagebrush
Artemisia tridentata Nutt. subsp. xericensis Winward ex R. Rosentreter & R. Kelsey, xeric big sagebrush
Hybridization occurs among mountain big sagebrush and other sagebrush taxa in zones of overlap [209,210,220,231,235,382,385,386,542,574,602,695]. Some of these hybrids have been separated out taxonomically. A hybrid of mountain big sagebrush and Wyoming big sagebrush in Utah and Idaho is designated as Bonneville big sagebrush [220]. Some subspecies may have hybrid origins with mountain big sagebrush [231,382,542,602]. Xeric big sagebrush may have derived by hybridization of mountain big sagebrush and basin big sagebrush [542], and snowfield big sagebrush may have derived by hybridization of mountain big sagebrush and silver sagebrush [231]. Hybridization is likely the source of new genetic combinations that helped big sagebrush adapt to past climate changes, and such hybridization may help big sagebrush adapt to climate changes in the future [381,387].
Subspecies of big sagebrush differ in ploidy levels. Mountain big sagebrush and basin big sagebrush can be either diploid or tetraploid, while Wyoming big sagebrush is tetraploid [381,386,531].
This review refers to plant species and infrataxa by their common names. See table A1 for scientific names of plants mentioned in this review and for links to FEIS Species Reviews.
SYNONYMS: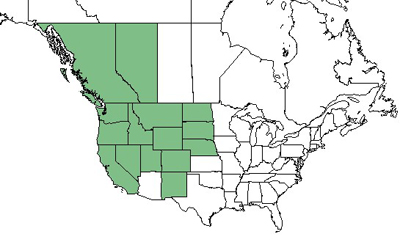 |
| Figure 2—Distribution of mountain big sagebrush. Map courtesy of the PLANTS Database [643], [2016, March 29]. |
Mountain big sagebrush is native to western North America and occurs from southern Alberta and British Columbia south to California, and east to northern Arizona, northern New Mexico, Colorado, Wyoming, and Montana [66,255,261,383,442,480,573,643]. Small populations of mountain big sagebrush occur as far east as North Dakota, South Dakota, and Nebraska [52,236,304,643].
The Columbia Basin, the Great Basin, and the Wyoming Basin support most (~70%) sagebrush in North America. The largest areas of sagebrush in the United States are in the Columbia Basin, where 32% of all sagebrush in the sagebrush ecosystem occurred in 2005, and the Great Basin, where 28% of all sagebrush occurred. In the Great Basin, mountain big sagebrush communities alone comprised 2,712,142 acres (1,097,565 ha) or 3.7% of the land cover. Wyoming big sagebrush communities comprised 17.9%, while black sagebrush and low sagebrush communities comprised 5.1% and 1.1% of the land cover in the Great Basin, respectively. All other sagebrush types comprised <1% [609]. The area occupied by mountain big sagebrush and other sagebrush communities has been reduced by livestock grazing and associated land management, agriculture, urban and industrial land uses, proliferation of nonnative plants, woodland expansion, climate changes, and other factors [79,141,419] (see Other management considerations).
States and provinces [643]:
United States: AZ, CA, CO, ID, MT, ND, NE, NM, NV, OR, SD, UT, WA, WY
Canada: AB, BC
Site characteristics: Topography, climate, and soils affect the distribution of the three major subspecies of big sagebrush [293,380,439]. Mountain big sagebrush occurs most commonly at high elevations, on cool, relatively moist sites, in moderately deep to deep Mollisols [227,232,263,293,294,433,481,612,626,699]. Sites occupied by parental taxa and their hybrids are distinct from one another [211,379,381,409,684].
Topography: Mountain big sagebrush occurs in mountain valleys and on foothills, mountain slopes, rolling uplands, shallow snowdrift depressions and swales, and high-elevation ridges [52,53,66,136,383,433,453,623,626,737]. It occurs as low as 2,560 feet (780 m) in Idaho to as high as 10,800 feet (3,300 m) in Colorado (table 1). Among the three major big sagebrush subspecies, mountain big sagebrush occurs at the highest elevations [417,437]. It occurs on all aspects [398,423], although at high elevations it may be restricted to south- or west-facing slopes [453]. It typically occurs on sites protected from wind [106,407].
| Table 1—Elevational range of mountain big sagebrush by location. | |
| Location | Elevation |
| Arizona | 4,490-7,400 feet (1,370-2,255 m) [266] |
| California | 5,910-9,840 feet (1,800-3,000 m) [261,278,617] |
| Colorado | 7,700-9,500 feet (2,350-2,900 m) [72,623,626]; in the Gunnison Valley it reaches "unusually" high elevations, with isolated populations at 10,830 feet (3,300 m) [336] |
| Idaho | 2,560-8,990 feet (780-2,740 m) [383,734] |
| Montana and British Columbia | 3,770-7,810 feet (1,150-2,380 m) [18,335,396,480] |
| Nevada | 5,450-10,170 feet (1,660-3,100 m) [408,504,617,635,638,763] |
| Oregon | 3,500-9,000 feet (1,070-2,740 m) [300,737] |
| Utah | 4,690-8,990 feet (1,400-2,740 m) [118,408,708] |
| Wyoming | 7,250-8,990 feet (2,210-2,740 m) [106,115,144,499] |
Climate: Mean annual precipitation in areas with mountain big sagebrush ranges from as low as 12 to 14 inches (300-350 mm) [232,417,745] to as high as 20 to 30 inches (500-750 mm) [227,596,745], and precipitation falls mostly during winter and spring [550]. On sites receiving <14 inches (350 mm) of annual precipitation, mountain big sagebrush stands often grade into Wyoming big sagebrush stands. Occasionally, stands of mountain big sagebrush occur on sites receiving >30 inches (750 mm) of annual precipitation. These sites are mostly confined to southerly exposures within the quaking aspen and conifer forest belts [227,232]. Seasonal precipitation patterns in sagebrush ecosystems result in a temporal pattern of moisture availability. Spring soil water recharge is followed by a dry period that begins in late spring and lasts until early winter. Top soil layers dry earlier and more completely than lower layers. Mountain big sagebrush sites dry out later and wet up earlier than sites dominated by other big sagebrush taxa [550,703]. Thus, mountain big sagebrush sites have the narrowest window of opportunity for fire in a given year [703]. Summer precipitation in sagebrush ecosystems varies from almost none in central Nevada to nearly 40% of the annual total in southern Utah, northern Arizona, and northern New Mexico [155,719]. Summer storms can be brief and intense, and most precipitation runs off or evaporates [478]. Wetter than average years promote accumulation of fine fuels that can carry fire the following year. Wildfires typically start during dry lightning storms in dry, hot summers but can also occur in wet years [428].
Among the three major subspecies of big sagebrush, mountain big sagebrush occupies the coolest and moistest sites [417,437,699]. For example, in Utah, mean annual precipitation was higher and mean minimum January temperature was lower in mountain big sagebrush communities than basin big sagebrush or Wyoming big sagebrush communities [437] (table 2).
Growing seasons are short on mountain big sagebrush sites. On the Humboldt National Forest, Nevada, the growing season began between 1 April and 13 May and extended to 3 May to 18 July, depending on elevation and plant community type. It ranged from 28 to 63 days in mountain big sagebrush communities and 46 to 76 days in mountain snowberry-mountain big sagebrush communities [297].
Mountain big sagebrush sites have deep snow pack [623] that may persist continuously for 6 months [406]. Snow pack tends to be deeper and last longer on mountain big sagebrush sites than on Wyoming big sagebrush sites [437]. In south-central Wyoming, mountain big sagebrush was most common where snow depth was >15 inches (38 cm), and Wyoming big sagebrush was most common where snow depth was <16 inches (40 cm) [607].
| Table 2—Climate characteristics of five plant communities in Utah. Values are means with standard errors in parentheses [437]. | |||||
| Characteristics | Mountain big sagebrush | Basin big sagebrush | Wyoming big sagebrush | Mountain brush | Pinyon-juniper |
| Frost-free days | 105.2 (13.0) | 147.2 (7.5) | 115.4 (9.8) | 102.8 (1.1) | 117.5 (6.8) |
| Temperature (°F) | |||||
| maximum January | 36.0 (1.0) | 39.9 (1.0) | 38.6 (0.9) | 36.6 (1.3) | 38.8 (1.6) |
| minimum January | 11.7 (2.6) | 17.2 (1.0) | 12.6 (1.1) | 13.5 (1.7) | 13.8 (0.8) |
| maximum July | 85.0 (1.2) | 91.0 (0.9) | 90.4 (1.0) | 86.9 (1.1) | 88.0 (1.2) |
| minimum July | 49.6 (2.0) | 58.3 (1.2) | 52.7 (1.5) | 50.9 (1.8) | 53.7 (1.4) |
| Mean precipitation | |||||
| Annual (inches) | 16.5 (0.7) | 13.8 (1.1) | 10.6 (0.7) | 19.6 (1.1) | 12.3 (0.6) |
| January-March (%) | 30 | 29 | 25 | 32 | 25 |
| April-June (%) | 24 | 25 | 26 | 23 | 22 |
| July-September (%) | 20 | 20 | 25 | 17 | 28 |
| October-December (%) | 26 | 26 | 24 | 26 | 24 |
| Cumulative snowfall (inches) | 69.8 (5.5) | 46.7 (5.9) | 38.9 (4.7) | 95.5 (11.6) | 55.4 (7.8) |
| Cumulative solar radiation (langlays) | 480.9 (14.3) | 464.1 (11.9) | 510.0 (8.7) | 480.4 (12.5) | 480.5 (16.7) |
| Annual daily minimum relative humidity (%) | 33 (3) | 37 (3) | 26 (2) | 33 (2) | 33 (3) |
| Annual pan evaporation (inches) | 51.6 (2.3) | 66.1 (2.0) | 64.3 (4.8) | 52.2 (2.7) | 56.8 (2.7) |
Soil order: While mountain big sagebrush occurs on Mollisols, Aridisols, and Inceptisols, it occurs most often on Mollisols [227,263,293,294,418,481,612,626,763]. Mollisols develop in areas where grasses have been codominant to dominant for a prolonged period. Aridisols develop where conditions are very dry and potential evapotranspiration typically exceeds precipitation, and they are not associated with specific plant life forms [196]. On the Humboldt National Forest, mountain big sagebrush communities occurred on soils with the thickest mollic epipedons compared to other sagebrush community types [294], indicating that these mountain big sagebrush communities historically had a productive herbaceous layer (see Fuels).
Soil moisture: Soil moisture availability tends to be greater on mountain big sagebrush sites than other sagebrush sites, and mountain big sagebrush is the least drought tolerant among the three major sagebrush subspecies. On the Humboldt National Forest, the transition from black sagebrush to low sagebrush, basin big sagebrush, and mountain big sagebrush stands showed a gradient of increasing available soil moisture [296]. Among the three major big sagebrush subspecies, soil moisture is the least limiting on mountain big sagebrush sites [411,612,736]. In southern Idaho, soil moisture was not deficient until September in two mountain big sagebrush stands and did not become deficient in another. Soil moisture was deficient by mid- to late July in Wyoming big sagebrush stands and by late July or early August in basin big sagebrush stands [736]. When water is limiting, mountain big sagebrush is less drought tolerant than basin big sagebrush or Wyoming big sagebrush. In Oregon, all subspecies were water stressed in August, but mountain big sagebrush was the most water stressed, followed by basin big sagebrush and then Wyoming big sagebrush [411].
All subspecies of big sagebrush grow on well-drained soils [66,146,306,612,684,699]. In general, big sagebrush plants cannot survive long in saturated soils [233,699]. Welch [699] suggested that a high water table may result in the absence of big sagebrush from many western grasslands. In southern Wyoming, mountain big sagebrush on the downwind side of a 12.5-foot (3.8 m) tall snow fence was eliminated by the third growing season after fencing, apparently because the snow fence resulted in deeper snow that left the soil saturated for a longer time. Forty feet (12.2 m) downwind of the fence, average snow depth increased from 4.9 feet (1.5 m) before fencing to 15.1 feet (4.6 m) after fencing. The authors identified a snow depth of 9.2 feet (2.8 m) as a threshold for mountain big sagebrush survival on this site [606].
Soil texture: Mountain big sagebrush grows in a range of soil textures, including clays, silts, loams, and sands, and soils are often gravelly or cobbly [146,217,263,307,351,423,460,481,638,671,678].
Soil texture can help to differentiate sites with basin big sagebrush from sites with Wyoming big sagebrush, whose distributions are closely associated with soil texture [72,146,574]; however, mountain big sagebrush occurs on sites with soil textures common to the other two subspecies. Instead, mountain big sagebrush sites are best separated from basin big sagebrush and Wyoming big sagebrush sites along gradients of elevation and precipitation, with mountain big sagebrush occurring at higher elevations receiving more precipitation than the other two subspecies [72,146] (see Topography and Climate).
Soil fertility: Mountain big sagebrush commonly occurs in soils with high fertility relative to other sagebrush taxa. On 372 relatively undisturbed sites on the Humboldt National Forest, soil organic carbon, nitrogen, and phosphorus levels indicated that mountain big sagebrush and basin big sagebrush commonly dominated sites with the highest soil fertility, and black sagebrush commonly dominated the least fertile sites. Mollic epipedon depth, total depth, and water-holding capacity followed the same pattern, with mountain big sagebrush communities having the highest values and black sagebrush communities the lowest [294]. In a landscape-scale study in Wyoming, soils in topographic depressions occupied by mountain big sagebrush had higher organic matter, nutrients, and microbial biomass than soils in more windblown areas occupied by other vegetation, including basin big sagebrush and Wyoming big sagebrush. This was attributed to greater snow accumulation in the topographic depressions, resulting in greater soil moisture and greater plant and litter biomass accumulation [105,106,107]. For a review of soil chemistry in big sagebrush communities, see Welch [699].
Plant communities: Mountain big sagebrush dominates sagebrush steppe communities from foothills to subalpine zones, codominates some mixed mountain shrublands, is a common component and sometimes an understory dominant in pinyon-juniper woodlands, and is a common component in some montane and subalpine grasslands and other woodland and forest types. Mountain big sagebrush steppe communities occur on mesic sites on stony flats, ridges, nearly flat ridgetops, and mountain slopes on all aspects, although at high elevations they may be restricted to south- or west-facing slopes [453]. Mountain big sagebrush steppe communities often contain multiple vegetation layers: 1) shrubs, 2) tall (8-24 inches (20-61 cm)) forbs and bunchgrasses, 3) low-growing (<8 inches tall) grasses and forbs, and sometimes 4) a biological soil crust [417]. These communities are generally treeless [757], or have scattered trees (i.e., very low density) [426], and vegetation is usually <5 feet (1.5 m) tall [453,719]. However, establishment and spread of junipers and pinyons since the late 1800s has caused widespread conversion of big sagebrush steppe to conifer woodland in areas where these ecosystems historically occurred adjacent to one another [426]. The greatest proportion of conifer expansion has occurred in sagebrush communities that occur in soil temperature and moisture regimes that range from frigid and xeric to mesic and xeric (table 7), including mountain big sagebrush communities at high elevations and low sagebrush and black sagebrush communities on moderately deep to deep soils [303,416,419] (figure 18). A number of interacting factors may contribute to juniper and pinyon infill in woodlands and expansion into big sagebrush communities, including lack of fire and a warming climate [108,109,185,429,539]. See Successional status and Woodland expansion for more information on this topic.
In mountain big sagebrush steppe communities, mountain big sagebrush may form dense monocultures [393] or be codominant with other shrubs, such as antelope bitterbrush. Other codominant or common shrubs in mountain big sagebrush steppe communities include mountain silver sagebrush, snowfield big sagebrush, stiff sagebrush, low sagebrush, rubber rabbitbrush, yellow rabbitbrush, mountain snowberry, wild crab apple, Wood's rose, wax currant, snowbrush ceanothus, and Saskatoon serviceberry [453]. Mountain big sagebrush co-occurs with many sagebrush taxa, including Wyoming big sagebrush, basin big sagebrush, low sagebrush, black sagebrush, and silver sagebrush [136,153,306,398,452,463,504,639,640,757].
Bunchgrasses characterize the understory of mountain big sagebrush steppe communities [757]. Historically, equal proportions of sagebrush and grasses were thought to be typical of big sagebrush steppe communities, although the proportion of each growth form likely varied depending on site characteristics and successional status [719,721]. Idaho fescue, bluebunch wheatgrass, blue grama, and Thurber needlegrass are the principal grasses on relatively dry mountain big sagebrush sites with relatively low productivity and on mountain big sagebrush sites with a well-developed argillic soil horizon. On deeper and often more loamy soils with relatively high productivity, bearded wheatgrass, melicgrass, slender wheatgrass, western needlegrass, and Columbia needlegrass are common [227,417]. Other graminoids that can be abundant include Thurber fescue, bottlebrush squirreltail, tufted hairgrass, Parry's oatgrass, western wheatgrass, California brome, prairie Junegrass, Fendler bluegrass, Sandberg bluegrass, and sedges [453].
Plant diversity is greater in mountain big sagebrush than other sagebrush communities [205,227], largely due to a high diversity of forbs [417,453]. In the North Spring Valley watershed, Nevada, the mountain big sagebrush type had 76 species, the black sagebrush type 70 species, the low sagebrush type 57 species, the Wyoming big sagebrush type 41 species, the pinyon-juniper type 43 species, and the winterfat type 18 species [205]. Across 112 mountain big sagebrush communities in the northern Great Basin, forbs accounted for 247 of the total 335 plant species encountered (Miller et al. 2000 cited in [417]). Common forb genera in mountain big sagebrush steppe communities include agoseris, Indian paintbrush, cinquefoil, fleabane, phlox, milkvetch, avens, lupine, balsamroot, hawksbeard, desertparsley, yarrow, and buckwheat, among others [417,453].
Biological soil crust cover typically increases with increasing aridity and is inversely related to vascular plant cover. Biological soil crusts are usually less abundant at higher elevations in mountain big sagebrush and mixed mountain shrub communities and more abundant at lower elevations in Wyoming big sagebrush, salt desert, and desert shrub communities [416]. In east-central Idaho, mountain big sagebrush communities had about 40% cover of biological soil crusts, while Wyoming big sagebrush communities had about 60% cover [308].
In the Intermountain West, mountain big sagebrush codominates some mixed mountain shrublands with Gambel oak, Saskatoon serviceberry, Utah serviceberry, mountain snowberry, roundleaf snowberry, true mountain-mahogany, curlleaf mountain-mahogany, antelope bitterbrush, and chokecherry [136,437,553,596,640,757].
Mountain big sagebrush is a common component of many montane and subalpine grasslands [153,160,208,548,640], such as subalpine green fescue meadows in Washington and Oregon [208], montane Idaho fescue-bluebunch wheatgrass meadows in Wyoming [640], and montane shrubby cinquefoil-Idaho fescue communities in Oregon, Washington, and Wyoming [153,640]. Mountain big sagebrush communities occur in a landscape mosaic with grasslands and forests, often occurring as small patches, but also as large, contiguous patches [640] (see Fuels).
Mountain big sagebrush is a common component and frequently an understory dominant in juniper and pinyon savannas and woodlands, including those with Colorado pinyon, singleleaf pinyon, Rocky Mountain juniper, Utah juniper, and western juniper (e.g., [132,136,152,185,371,452,463,512,540,625,637,649,722,737]). In the western United States, junipers are more widespread than pinyons [416,539]. For the geographic range of specific pinyons and junipers, see Romme et al. [539] or the Plants Database [643].
Mountain big sagebrush is also a common component in ponderosa pine and Jeffrey pine savannas and woodlands [255,291,298,452,469,504,506,512,540,591,766], and it occurs in openings in quaking aspen [106,118,151,369,445,449,504], water birch [369], Gambel oak [504,766], curlleaf mountain-mahogany [136,152,306,452,463,504,553,766], Rocky Mountain Douglas-fir [255,306,469,506,591,766], limber pine [53,136,255,469,506,509,589,591,639], lodgepole pine [53,118], whitebark pine [53,450,548,636], subalpine fir [208,396,591,639], white fir [636], Engelmann spruce [636], and blue spruce [766] communities. As tree cover increases, mountain big sagebrush cover declines [460] (see Successional status).Mountain big sagebrush has both ephemeral and persistent leaves [628,670,699]. Its small flowers are borne on many-branched, narrow inflorescences that are about 4 to 6 inches (10-15 cm) long [204,573,716]. Researchers have described the one-seeded fruit of mountain big sagebrush as both an achene [403,760] and a cypsela [233,573]. This review uses the term achene. Achenes are about 0.05 to 0.07 inch (1.25-1.78 mm) long and 0.02 to 0.03 inch (0.5-0.84 mm) wide [255,760]. A single mountain big sagebrush achene averages as little as 0.25 to 0.26 mg ([734,736], Meyer et al. 1987, unpublished report cited in [403]) to as much as 0.60 mg [255]. In Gunnison, Colorado, mountain big sagebrush achene weight varied with elevation, ranging from 0.27 mg at 8,000 feet (2440 m) to 0.46 mg at 11,000 feet (3,350 m) [336].
Sagebrush taxa are difficult to distinguish from one another based on morphological characteristics alone. In portions of the Great Basin in California, Nevada, Utah, and Idaho, West et al. [722] found that only 32.6% of the sagebrush specimens examined, which included mountain big sagebrush, basin big sagebrush, Wyoming big sagebrush, black sagebrush, and low sagebrush, were correctly identified based on morphological characteristics alone. However, differences in leaf chemical constituents—particularly concentrations of coumarins [209,379,733]—determined by examining fluorescence of leaves and other plant parts under ultraviolet light [209,379,386,575,587,595,699,733]—can help differentiate sagebrush taxa, especially when used in combination with plant morphological characteristics [52,155,217,261] and site characteristics (e.g., topography, climate, and soils) [217,386].
Mountain big sagebrush develops a dense root network both in the upper soil layers and deep in the soil profile [614,628,699], making it highly competitive for water and nutrients [605,628]. It has a taproot that can extend 3 to 7 feet (1-2 m) vertically, and lateral roots that can extend 5 feet (1.5 m) horizontally [528,614,628,699]. Most mountain big sagebrush roots occur in the upper 1 foot (0.3 m) of soil [614,628,699]. Soil characteristics such as texture, aeration, and moisture influence root distribution [233]. For example, mountain big sagebrush roots were slightly deeper on a dry ridge than on a moist valley bottom and deeper on western than eastern slopes, despite similar soil characteristics [614]. Mountain big sagebrush roots are colonized by fungi that form symbiotic vesicular-arbuscular mycorrhizae [112,635].
Mountain big sagebrush plants may live >100 years, although most plants die when they are <50 years old [90,267,498,499,628,680,699]. On 27 sites throughout Wyoming, the maximum ages were 79 years for mountain big sagebrush, 75 years for Wyoming big sagebrush, and 55 years for basin big sagebrush [498,499]. On 33 sites on the Northern Yellowstone Winter Range of southwestern Montana, mountain big sagebrush plants ranged from 5 to 109 years old. Most plants were <50 years old; 12% were >50 years old, and 5% were >60 years old. The oldest living mountain big sagebrush plant was 88 years old, and the oldest dead plant was 109 years old [267,680]. Large plants (≥9 inches (22 cm)) averaged 32 years old and small plants (<9 inches) averaged 6 years old. Nearly half of the small plants were 5 years old, establishing after the 1988 Yellowstone Fires. Dead plants averaged 41 years old [267]. Dead stemwood is common in old plants. The fraction of stemwood that was dead on mountain big sagebrush and Wyoming big sagebrush plants averaging 36 years old was 48% [90].
Stand structure: Total vegetation cover in mountain big sagebrush communities is often high, and cover of individual species depends on site characteristics and successional stage. Mountain big sagebrush can make up a large portion of the total, especially in late succession. At sites in the Uinta Mountains, Utah, with a history of about 100 years of livestock grazing, total vegetation cover in mountain big sagebrush communities ranged from 87% to 95% and averaged 92%. At one of these sites, cover was 65% just 1.5 growing seasons after fire [229]. According to reviews, shrub canopy cover in undisturbed mountain big sagebrush communities typically varies between 15% and 40% but can reach up to 50% or more in relatively mesic communities on deep loamy soils and on northern aspects [417]. In moderately deep to deep loamy soils in an area of Carbon County, Wyoming, where mean annual precipitation ranges from 10 to 15 inches (254-381 mm), mountain big sagebrush canopy cover on 15 undisturbed, late-seral stands averaged 56% (range: 42%-71%) [686]. In stands undisturbed for 30 years in the Uinta Mountains, Utah, mountain big sagebrush cover ranged between 25% and 40% [227]. In a 70-year postfire chronosequence near Dubois, Idaho, mountain big sagebrush reached steady-state cover (when cover did not change with increasing time-since-last burn) of 31% at 19 years after fire [432]. Mountain big sagebrush can form homogenous-looking, dense stands [393] where shrubs are large enough and dense enough to impede access of wildlife and livestock to understory plants [100]. Stands can have a dense perennial herbaceous layer with >25% cover, and in many cases >50% cover [453,698]. As mountain big sagebrush cover increases, herbaceous cover declines [196,735] (see Successional status). Welch [698] provides a review of herbaceous plant production in big sagebrush communities.
Mountain big sagebrush shrublands often occur in a landscape mosaic with forests, woodlands, shrublands, and grasslands [208,369,452,640]. Boundaries between mountain big sagebrush communities and other communities may be gradual or abrupt [639], and are driven by changes in soils, microtopography, and disturbance regimes [294,295,413,481]. Patches of mountain big sagebrush range from small to large [640].
Raunkiaer [521]
life form:
Phanerophyte
SEASONAL DEVELOPMENT:
Annual growth of big sagebrush plants begins in early spring, and ceases when soil moisture is depleted, usually by late July [52,297,628,699]. Root growth begins before stem growth and continues through the late fall [699]. Big sagebrush produces two types of stems or shoots: short shoots and long shoots [699]. Short shoots produce leaves during early summer, which persist over winter and die the following summer [412,628,699], living a total of 12 to 13 months [699]. The persistence of winter leaves allows big sagebrush to use soil water and nutrients early in the growing season [412]. Current season long shoots produce shorter-lived, ephemeral leaves, which develop in spring and senesce and die in mid- to late summer [628,699], when water stress is high [411,412]. This enables big sagebrush to maximize use of resources (e.g., water and nitrogen) during optimum growing conditions and avoid desiccation during summer drought [412].
Inflorescences are produced on short shoots, which arise from 1-year-old long shoots [699]. Mountain big sagebrush plants usually flower in late summer and fall, but some may flower as early as July [52,66,383,628] or as late as early October [761]. Flowering often occurs after the onset of freezing temperatures [761].
Seeds ripen in fall, from August through December, depending on latitude and elevation [52,66,233,336,383,530,628,760]. Winward [736] stated that mountain big sagebrush in areas with early fall frosts tended to have earlier maturing seeds. Seeds ripen over time, rather than all at once, so there is an extended period of seed dispersal in fall and winter [389,406,628,760]. For example, near Reno, Nevada, dispersal began within 2 weeks of initial seed maturity, peaked within 4 to 6 weeks, then tapered off (table 3) [760]. Although not reported for mountain big sagebrush specifically, researchers reported that the most viable big sagebrush seeds tend to be dispersed earliest, with aborted flowers and half-filled seeds dispersed later [233,255].
| Table 3—Mean number of mountain big sagebrush seeds collected from 0.01-m² traps near Reno, Nevada. Traps were checked after seed maturity at 2-week intervals from 15 December to 1 March, 1983–1986. Means within locations (rows) followed by different lower-case letters and overall means (last column) followed by different upper-case letters are significantly different (P ≤ 0.01) [760]. | ||||||||
| Study site | Trap location | Mean number of seeds |
||||||
2 weeks |
4 weeks |
6 weeks |
8 weeks |
10 weeks |
12 weeks |
Overall mean |
||
| Granite Peak | canopy edge | 60b | 220a | 30b | 20b | 50b | 0b | 380A |
| 1 m from edge | 40b | 70a | 40bc | 20cd | 0d | 0d | 170A | |
| Churchill Canyon | canopy edge | 10b | 180a | 20b | 10b | 0b | 0b | 220B |
| 1 m from edge | 20c | 60a | 30bc | 20cd | 0d | 0d | 120A | |
Germination is usually synchronous [403] and occurs in late winter and early spring, either soon after snowmelt or shortly before [336,389,403,406,407,628,760], although Wagstaff and Welch [661] reported that in north-central Utah a few small seedlings observed in September had apparently emerged during summer, perhaps after a summer thunderstorm.
Timing of phenological events depends on big sagebrush subspecies and site characteristics. In southern Idaho, timing of phenological development differed among big sagebrush taxa. Mountain big sagebrush began growing later and ripened seeds earlier than basin big sagebrush and Wyoming big sagebrush (table 4) [734]. In south-central Idaho, Winward [736] noted that variation in timing of seed maturity among basin big sagebrush and Wyoming big sagebrush plants appeared related to available moisture, with seeds maturing earlier on drier sites. Variation among mountain big sagebrush plants appeared related to temperature more than moisture, because areas with earlier fall frosts had earlier maturing mountain big sagebrush seeds [736]. In eastern Montana, big sagebrush growing above 5,900 feet (1,800 m) initiated growth 2 to 3 weeks later and initiated floral bud enlargement, anther development, anthesis, and dissemination 2 weeks earlier than big sagebrush at lower elevations [255].
| Table 4—Timing of phenological events for big sagebrush subspecies in south-central Idaho [734]. | ||
| Event | Mountain big sagebrush | Basin big sagebrush and Wyoming big sagebrush |
| early shoot development | early July | mid-June |
| medium shoot development | early July | late June to early July |
| full shoot development | mid-July | mid-July |
| flowerheads green | mid- to late July | late July |
| flowerheads yellowing | early August | early September |
| pollination | early September | mid-October |
| seed ripening | mid-October | early November |
REGENERATION PROCESSES:
Mountain big sagebrush regenerates from seed [469], and vegetatively by layering [54,55,255,383]. Seed production [339,713] and seedling emergence [70,333,420] are highly variable. Due to high rates of seedling mortality, most big sagebrush seedlings that emerge are not recruited into the population [70,309,403]. Thus, seedling establishment usually limits recruitment and drives the rate of postfire recovery [469,662,767]. Mountain big sagebrush does not sprout from the root crown or roots after the aerial portion of the plant is killed or removed [65,66,461].
Pollination and breeding system: Big sagebrush is wind-pollinated [695] and self-pollinated [244,379]. Outcrossing by wind leads to a greater production of viable seeds than self-pollination (McArthur 1984 cited in [695]). Because it is self-pollinating, isolated big sagebrush individuals can set seed [403]. For this reason, Meyer [403] stated that seed set is probably not strongly pollen-limited even in years when flowering is sparse. Information on embryo, seed development, and anatomy of big sagebrush can be found in a review by Welch [699].
Seed production: Mountain big sagebrush seed production is highly variable, and depends on site and plant characteristics (e.g., size, age, and genetics) [339]. At 13 sites in central and south-central Utah, the mean number of seeds per mountain big sagebrush plant ranged from 0 to 2,214 for small plants (basal stem diameter <0.4 inches (1 cm)), 841 to 36,265 for medium plants (basal stem diameter = 0.4-1.2 inches (1-3 cm)), and 0 to 117,079 for large plants (basal stem diameter >1.2 inches (3 cm)) [339]. A single, 20-year-old mountain big sagebrush plant in Montana produced an average of 115,400 seeds/year for 3 years [255]. At Salt Creek, Utah, mountain big sagebrush averaged 17 inflorescences/plant with an average of 102 flowering heads/inflorescence. It averaged 452 seeds/branch [235]. In a common garden, seed production in 2- and 3-year-old plants ranged from 0 to 137 grams of live seed/plant [713].
Plant age, time-since-disturbance, and plant genetics affect mountain big sagebrush seed production. Mountain big sagebrush plants as young as 2 years old can produce abundant seeds, as observed in wild populations [43,471], seeded field plots [161], common gardens [713], and greenhouses [763]. However, age of first seed production varies, and observations in two studies suggest that mountain big sagebrush individuals may not reach reproductive maturity until they are 5 to >10 years old [267,309]. In Mono County, California, most mountain big sagebrush plants began flowering when they were 5 to 7 years old, but some did not flower during their first 13 years [309]. No mountain big sagebrush plants growing in burned areas <10 years old on the Northern Yellowstone Winter Range had reproductive stalks, and no mountain big sagebrush seedlings established after the fire. Some of the plants were older than the burns [267].
Seed production may decrease as mountain big sagebrush plants age [339], but even 100-year-old big sagebrush plants can be "prolific seed producers" [763]. At 13 sites in central and south-central Utah, seed production decreased with time-since-fire for small, medium, and large mountain big sagebrush plants (P-value range = 0.019-0.051); however, seed densities at each site increased with increased mountain big sagebrush densities over time (P = 0.004). Seed densities leveled off between 20 and 30 years after fire—before stands reached maximum density and cover [339].
Seed production varies among individual big sagebrush plants in common gardens, where resource limitation is not a factor, suggesting genetic differences in seed production potential [403,713].
Site characteristics and moisture availability can also affect seed production. Over a 3-year period, annual seed production in mountain big sagebrush stands differed between two sites near Reno, Nevada, likely due to differences in moisture availability. Average annual seed production was greater (185 pounds/acre (21 kg/ha)) on sites where average annual precipitation was 16 inches (406 mm) and soils were Mollisols, compared to 115 pounds/acre (13 kg/ha) on sites where average annual precipitation was 12 inches (310 mm) and soils were Aridisols (P < 0.01) [763]. It is likely that mountain big sagebrush growth and reproduction at higher elevations is limited by moisture availability and growing season length. At 13 sites in central and south-central Utah, seed production in large plants (basal stem diameter >1.2 inch (3 cm)) was positively correlated with winter precipitation (P < 0.0001) and negatively correlated with elevation (P = 0.0001), while that of small plants (<0.4 inch (1 cm)) and medium plants (0.4-1.2 inch) was negatively correlated with winter precipitation (P-value range = 0.0106-0.0174) [339].
Summer weather can lead to differences in seed production among years because seed stalks form in late summer (see Seasonal development). Flowering may not occur in big sagebrush if there is too little precipitation during this time [761]. However, too much precipitation before seed maturity can cause seeds to rot on the stalk [159].
Competition for resources may reduce mountain big sagebrush seed production (e.g., [391,763]). In southeastern Oregon, the median number of reproductive branches on mountain big sagebrush was less in the interior of an unburned site (325.5 (SE 52.9)) than within 7 feet (2 m) of the edge of a 1-year-old prescribed burn (458.9 (SE 30.5), P = 0.05), which may have been due to reduced competition for resources as well as a postfire increase in available nitrogen [391].
Overbrowsing by ungulates can drastically reduce mountain big sagebrush seed production [61,538,661,662]. Near Hobble Creek, Utah, protection of a mountain big sagebrush stand from browsing for a single winter resulted in a 20-fold increase in the number of seed stalks on mountain big sagebrush plants the following fall. Successive years of protection resulted in further increases in seed production for 6 years [662,713].
Plant disease, such as stem rust fungi, can reduce seed production in cultivated and probably native stands of big sagebrush (Nelson 1992 personal communication cited in [403]). Insect seed predators and herbivores like thrips that feed on flower parts may also reduce seed production [403].
Seed dispersal: Big sagebrush seeds are dispersed by wind, water, and animals [52,233,255,404,495,628,760]. Among these, wind may be the most important [233], but it is ineffective for long-distance dispersal [495,701]. Pendleton et al. [495] classified the dispersal mechanism of big sagebrush seeds as "microwind", meaning that the seeds are dispersed by wind because they are light and small but lack any special structures that would allow them to float or travel long distances [701]. Most long-distance dispersal of big sagebrush seed may be attributed to large mammals because the mucilaginous seed coat of big sagebrush can attach to their fur [255]. Many large animals, such as mule deer and elk, use burned big sagebrush areas [674]; thus, large animals may disperse big sagebrush seeds to these areas. No big sagebrush seed was reportedly dispersed by rodents near Reno, Nevada [334]. Big sagebrush seeds float in water, aiding in water dispersal [233], and big sagebrush occurs along stream banks [233].
Most big sagebrush seeds, including mountain big sagebrush seeds, are dispersed within 10 feet (3 m) of parent plants, and most fall under the crown [213,255,628,661,701,760] (table 3); however, big sagebrush seedling establishment patterns indicate that some seeds disperse farther (e.g., [213,661]). In eastern Montana, mountain big sagebrush seeds dispersed within 10 feet (3 m) of parent plants, and most fell within 3 feet (1 m) [255]. In general, the maximum distance of big sagebrush seed dispersal is about 100 feet (30 m) from the parent plant [233,416,699]. Most big sagebrush seeds are dispersed on the downwind [233,255,661,701,760] or downhill [320] side of plants.
Colonization of large burned areas by big sagebrush may be a slow process because of short dispersal distances [34]. Assuming a maximum dispersal distance of 100 feet and an age of first reproduction of 2 to 4 years, Welch [699] concluded that big sagebrush could spread from 25 to 50 feet (7-15 m)/year, and noted that it would take about 105 to 211 years to spread 1 mile (1.6 km). Seeds may disperse farther on steep or windy sites [320], so these may be colonized faster.
Seed banking: Mountain big sagebrush has transient [760] or short-term persistent (1-5 years) seed banks [447,729,767,768]. In storage, viability and germinability of mountain big sagebrush seeds can be extended to 5 or more years [404,691,701,702]. In the field, most mountain big sagebrush seeds are lost from the seed bank through germination in late winter and early spring, when primary dormancy is broken [401,760]. Postdispersal loss of big sagebrush seeds to rodents appears minimal [334], possibly because of small seed size [403]. Of the remaining seeds, a small fraction persists in soil seed banks if suitable conditions are not present to cue germination in late winter and early spring and secondary dormancy is induced [403,729].
In late spring and summer, seed abundance in the soil varies from low to none. At six sites in the Great Basin, low densities of viable seeds of both mountain big sagebrush and Wyoming big sagebrush were present in litter and soil in August, 9 months after seeds dispersed, although abundance of seeds in soils varied considerably between years and among sites [729]. Despite similar methods for measuring viable seed abundance in soils, a study at two sites in northwestern Nevada reported no viable mountain big sagebrush seeds in litter and soil sampled in June, 6 months after seeds dispersed (table 5) [760]. Differences between the studies were attributed to the patchy distribution of seeds in soils. Also, seed longevity is enhanced by burial in the soil, and study sites may have differed in soil disturbances that bury seeds [729] (see Planting depth). Meyer [403] stated that the fraction of big sagebrush seeds that enters the persistent seed bank is probably <1%, but on sites where seed production is high this fraction could be ecologically important.
The number of mountain big sagebrush seeds in the soil seed bank likely varies among sites depending on cover and productivity of mature plants and timing of sampling relative to seed dispersal (table 5); however, few studies are available that quantify mountain big sagebrush seed banks. Seed density in soil seed banks is positively correlated with big sagebrush canopy cover [9,256,370]. Davies [161] hypothesized that mountain big sagebrush seed banks may be depleted in mid- to late-successional western juniper woodlands because mountain big sagebrush cover is reduced by canopy shading [423] (see Successional status). In western juniper/mountain big sagebrush/Idaho fescue communities with 10% shrub cover in Oregon, only 2 and 5 mountain big sagebrush seeds germinated from soil samples taken during late October and early November (after most seeds had dispersed) in 2 consecutive years [182]. In a singleleaf pinyon-Utah juniper/mountain big sagebrush community in central Nevada, the germinable seed bank in early May was dominated by mountain big sagebrush, which comprised 44% of the total. Two years later in late October, after most seeds dispersed, it comprised 15% of the germinable seed bank composition [10]. Canopy cover of mountain big sagebrush was not provided. Differences among studies may also be attributed to germination procedures and differing rates of dormancy among sites [182].
| Table 5—Mean number of mountain big sagebrush seedlings germinated from soil samples for each sample date. Means within rows followed by different letters are significantly different (P ≤ 0.01). Samples were collected every 6 weeks from 1983 through 1986 in northwestern Nevada. Seeds ripened and dispersed in December. Shrub canopy cover ranged from 20% to 35% at the two study sites [760], although differences between sites were not provided. | ||||||||
| Study site | Seedling emergence |
|||||||
1 Dec |
15 Jan |
1 Mar |
15 Apr |
1 Jun |
15 Jul |
1 Sep |
15 Oct |
|
| Granite Peak | 0 |
560ab |
300b |
30b |
0 |
0 |
0 |
0 |
| Churchill Canyon | 0 |
680a |
560a |
180a |
0 |
0 |
0 |
0 |
Establishment of mountain big sagebrush 1 or more years after large, severe fires suggests that mountain big sagebrush seeds may persist in soil seed banks at least that long [527,698,768]. The North Fork Fire, one of the many fires that burned 793,880 acres (321,270 ha) in Yellowstone National Park in 1988, killed all sagebrush plants within the burn perimeter, eliminating on-site seed sources except from the edge of the burn. Given the limited dispersal distance of mountain big sagebrush seeds and the apparent even-aged distribution of mountain big sagebrush plants in the burn 10 to 11 years after the fire, Rens [527] concluded that postfire establishment of mountain big sagebrush was from the soil seed bank. Five years after an August 1999 wildfire near Provo, Utah, that killed all mountain big sagebrush plants, 0 to 47 big sagebrush seedlings (a mixture of mountain big sagebrush and Wyoming big sagebrush plants) were observed within 1-acre (0.4-ha) plots along a transect. Because the closest seed source was 0.4 mile (0.6 km) away, the author concluded that these seedlings originated from soil-stored seeds surviving the fire. The author did not report in which of the 5 postfire years the seedlings established [698]. One study in southern Oregon and northwestern Nevada suggested that viable mountain big sagebrush seeds may persist in soils for up to 3 years. The study examined the recruitment year of mountain big sagebrush plants in four large (990-9,880 acres (400-32,000 ha)), 6- to 41-year-old wild and prescribed fires with few to no unburned patches in burn perimeters. In the three youngest burns, recruitment was highest in the first and second postfire growing seasons, then declined but remained high in the third growing season, followed by a period of little to no mountain big sagebrush recruitment [768]. It was unclear whether young plants produced seed and contributed to recruitment after the second postfire year. In the oldest burn, little to no recruitment occurred during the first 17 postfire years [768]. Mountain big sagebrush cover prior to the fires was not known.
Fire kills many seeds in soil seed banks [10] (see Immediate effects on seeds). Thus, repeated burning is likely to deplete mountain big sagebrush seed banks and increase the postfire recovery period [131].
Germination: Seed viability and dormancy: Viability of mountain big sagebrush seeds can be high. Viability of mountain big sagebrush seeds collected from 24 sites in Wyoming, Utah, Idaho, Nevada, California, and Oregon ranged from 84% to 98% [407]. Germination rates of viable mountain big sagebrush seeds vary, but may exceed 90% [255,405,761], depending on environmental conditions. Temperature, light, moisture, and planting depth affect germination rates [255,389,404,408]. Yearly differences in germination rates of mountain big sagebrush seed from a single population appear to be minimal [193,250,255].Some mountain big sagebrush seeds are dormant at the time of dispersal [255,408,760]; the fraction of dormant seeds varies among sites. Mountain big sagebrush seeds collected from sites with long, severe winters germinated slowly and had a larger proportion of dormant seeds than those from sites with shorter, milder winters [405,406,407,408]. Of the mountain big sagebrush seeds collected from 24 sites in Wyoming, Utah, Idaho, Nevada, California, and Oregon, 0% to 58% were dormant per site. Seed dormancy decreased as mean January temperature of the collection site increased (R² = 0.578, P < 0.001). Sites with cold winters (<23 °F (-5 °C) mean January temperatures) had the highest percentage of dormant seeds, and seeds from warmer sites were essentially nondormant [407]. Climate-correlated variation in dormancy appears to be an adaptive feature in big sagebrush that prevents precocious germination and favors germination when frost damage potential is low and chances for establishment are greatest [404,596]. This illustrates the importance of planting seeds that are adapted to the specific climate of a site in rehabilitation projects [404,405,596] (see Value for rehabilitation of disturbed sites). Dormancy in mountain big sagebrush seeds can be broken under natural conditions by cold stratification, light, and removal of the pericarp by weathering and soil microorganisms [390,408].
Temperature: Mountain big sagebrush seeds germinate within a wide range of temperatures [193,255,389,408], but germination appears to be best between 54 and 68 °F (12-20 °C) [255,761,764]. Cold stratification for about 2 to 8 weeks increased mountain big sagebrush germination rates, with seeds collected from warm-winter sites germinating more rapidly than seeds collected from cold-winter sites [389,406].
Light: Mountain big sagebrush seeds germinate in light and dark, but germination rates are higher in light. Germination in light is much faster than germination in dark, regardless of temperature [408]. Mountain big sagebrush germination rates increase as snow cover melts in early spring and sunlight reaches the seed bed [401,405,408].
Moisture: Germination of mountain big sagebrush seeds occurs in late winter and early spring, either shortly before or soon after snowmelt when moisture availability is high [336,389,403,406,407,628,701,760] (see Seasonal development). Open, exposed sites are not favorable seed beds because they tend to dry too rapidly [596]. Litter may provide favorable seed beds by creating moist, protected sites [596], but deep litter inhibits germination [403] (see Planting depth). Mountain big sagebrush seeds can germinate under snow [401], and snow cover provides a moist environment and may protect germinants from spring frosts [596]. Big sagebrush seeds require saturation of the surface soil for germination [233,715]. The highest field germination of big sagebrush in Asotin County, Washington, occurred when "the surface of the soil was so saturated that free water appeared when pressure was applied" [233]; no information was provided on the length of time that soils were saturated.
Mature shrubs and downed juniper and pinyon trees trap snow, creating favorable seed beds for big sagebrush [403]. At a semiarid mined site in a year with average winter precipitation, big sagebrush seedling density was 6 times greater in areas where snow fences increased snow pack depth than where snow fences were absent (P < 0.05), suggesting that in years of average or perhaps below-average winter precipitation, big sagebrush seedling emergence is greater on sites with greater snow depth [438], as long as soils are not saturated for too long [606] (see Soil moisture).
Planting depth: Because of small size and limited energy reserves, mountain big sagebrush seedlings emerge best from the soil surface or when buried at very shallow depths (≤0.6 inch (15 mm)) [233,290,760].
Seeds may get buried by freeze-thaw and wet-dry cycles and winnowing [320,403,701]. Seed burial by winnowing may be more likely on sandy sites [320]. Deeper burial can induce secondary dormancy in mountain big sagebrush seeds and may protect seeds from lethal temperatures during fire [729] (see Immediate effects on seeds). Nelson [460] found a positive relationship between postfire recovery of mountain big sagebrush cover and percentage of sand in the soil, and speculated that sandier soils result in deeper seed burial, which enhanced viability, prolonged dormancy, and insulated seeds during fire, resulting in higher postfire seed retention. A study using seed bags placed at varying soil depths at six locations in the Great Basin found that no mountain big sagebrush or Wyoming big sagebrush seeds on the soil surface were viable after 2 years. In contrast, 30% to 40% of mountain big sagebrush seeds and 29% to 36% of Wyoming big sagebrush seeds remained viable when buried 1.2 inches (3 cm) deep [729].
Overview: Mountain big sagebrush plants establish best on sites with low density and low cover of other vegetation [267,398,486] (see Interference and competition), such as on recent burns. Postfire seedling establishment of big sagebrush can be high (600 seedlings/ft²) when moisture availability is adequate [628]. However, big sagebrush seedling emergence is highly variable [70,333,420] and can be low even after disturbances [70]. Due to high rates of seedling mortality, most big sagebrush seedlings that emerge are not recruited into the population [70,309,403,628]. Of the mountain big sagebrush seedlings that germinated in spring along Convict Creek (south of Mammoth Lakes, California), only 0.3% (5 of 1,890) in 2010 and 0.2% (8 of 3,720) in 2011 survived to late September [309]. Studies of age-class structure at nine undisturbed sites at high elevations in Wyoming suggested that big sagebrush seedling emergence and survival is episodic. Pulses of recruitment occurred at intervals ranging from 1.2 to 2.9 years for mountain big sagebrush stands, 1.3 to 2.7 years for basin big sagebrush stands, and 1.9 to 2.7 years for Wyoming big sagebrush stands [497,498].
Moisture availability: Mountain big sagebrush seedlings are adapted to and tolerant of seasonal drought [73,403] (see Climate); however, seedling mortality increases during unseasonable drought [522] and can be very high in the first few weeks of emergence in spring if moisture availability is low [70,336]. Mountain big sagebrush seedlings may be more susceptible to unseasonable drought than older plants because of their less developed root systems [441].
Mountain big sagebrush seedling survival may depend on snow cover at the time of emergence [403] because snow provides needed moisture [403,434,436] and protects seedlings from damage by late spring frosts [233,403,436,596], as well as browsing animals [527]. Deep (>3 feet (1 m)) snow pack is common in late spring at many mountain big sagebrush sites [106]. Among the mountain big sagebrush plants that established on the Northern Yellowstone Winter Range between 1978 and 1992, 47% germinated in 1988, a year of numerous fires. The following winter had considerably more snow than other winters during the 15-year period and a 35% to 40% reduction in elk numbers. The authors suggested that mountain big sagebrush seedling establishment was high that year not only because elk numbers were reduced but also because snow insulated seedlings from freezing temperatures and protected them from browsing by the remaining elk. Mountain big sagebrush seedlings established during the next several years, when elk numbers remained low [267,527,674,677].
Although moisture availability is critical for mountain big sagebrush seedling establishment, especially when seedlings emerge in spring, studies found little relationship between mountain big sagebrush recruitment and seasonal precipitation patterns [349,365,460]. In Beaverhead and Madison counties, Montana, recruitment of mountain big sagebrush, Wyoming big sagebrush, and basin big sagebrush over 35 years was not strongly associated with seasonal precipitation (previous summer, previous fall, winter, spring, or summer precipitation) in either burned or unburned plots [349]. In Wyoming, mountain big sagebrush recruitment was very weakly but positively correlated with years of below-average February (r² = 0.03), April (r² = 0.03), and May (r² = 0.03) precipitation after the first growing season (P < 0.05). The authors suggested that below-average spring precipitation coincides with warmer temperatures at high elevations, which may result in conditions favorable to mountain big sagebrush recruitment. However, temperature variables were poor predictors of recruitment, and the relationship between precipitation and temperature was not examined [365] (see Climate). On 36 sites burned between 1971 and 2007, mountain big sagebrush stands in relatively dry, low-elevation sites in the Great Basin and Colorado Plateau regions of Utah and eastern Nevada, time-since-fire explained 36% of the variance in mountain big sagebrush cover, and postfire October to March precipitation accounted for an additional 6% of the variance (R = 0.41, P < 0.0001), indicating that mountain big sagebrush establishment and growth was somewhat influenced by available moisture in the fall and winter after fire [460].
Interference and competition: Mountain big sagebrush seedlings establish on undisturbed sites under some conditions [333,355,497,498], but they seem to establish best on sites with low density and cover of vegetation, such as on recent burns (see Postfire seedling establishment). Seedling establishment is limited in areas with a high density of grasses and forbs [267,398,486] or dense big sagebrush [355]. Initial mountain big sagebrush seedling establishment may be high where seed densities are high, such as on artificially seeded sites, but mortality rates are high for closely spaced seedlings [436]. Because of high seed densities and synchronous germination, a review stated that competition among big sagebrush seedlings for water probably accounts for much of the initial mortality as soils dry out in late spring [403].
Dense grasses and forbs, both native and nonnative, can interfere with big sagebrush seedling establishment [177,267,329,398,486,681]; however, on some sites, neighboring forbs may facilitate mountain big sagebrush seedling establishment [336]. Postfire establishment of mountain big sagebrush occurred only in the first 1 to 3 years after fall prescribed burning at four sites near Dubois, Idaho. Increasing herb cover interfered with mountain big sagebrush establishment in postfire years 4 to 8 [177]. On the Northern Yellowstone Winter Range, dense grasses appeared to have interfered with big sagebrush establishment on a 19-year-old burned site [398], and high grass "vigor" after fire may have interfered with mountain big sagebrush seedling establishment on seven, 9- to 13-year-old burns. Three of the seven burned sites had no small mountain big sagebrush plants up to 13 years after fire [267,681]. On several sites in southeastern Idaho that burned in the 1930s, big sagebrush established within the first and second postfire years. After that, dense cover of grasses appeared to hinder big sagebrush establishment [486]. In parts of California [329], Nevada [329,760], and Utah [235], high densities of nonnative grasses may have interfered with mountain sagebrush seedling establishment. Removal of dense grasses and forbs by grazing livestock may result in increased big sagebrush seedling establishment in grasslands, even where nonnative species are present [216]. Historical overgrazing by livestock increased mountain big sagebrush density in many areas (see Livestock grazing).
In contrast, forbs were important for mountain big sagebrush seedling survival and growth in subalpine meadows near Gunnison, Colorado. Seedlings that had neighboring forbs removed were 3.7 times more likely to die during the course of the experiment than those with neighbors retained. Seedlings with neighbors were taller than those without neighbors at the end of the growing season (P = 0.021). Mountain big sagebrush seedling mortality was mostly due to desiccation of seedlings, suggesting that neighboring forbs moderate temperature and water stress for mountain big sagebrush seedlings on these sites [336].
Establishment of big sagebrush from artificial seeding is more successful when established plants are first removed by burning or other methods [436,596,701], and grasses and forbs are not seeded along with big sagebrush [329,529]. On a mined site in Idaho, no mountain big sagebrush established when herbs and mixed shrubs were seeded at the same time, but >2,000 mountain big sagebrush plants established when only mixed shrubs were seeded [529]. Grass biomass was greater than sagebrush biomass on plots seeded with grass prior to or as late as 1 year after sagebrush on a threetip sagebrush-big sagebrush site in Idaho. However, sagebrush biomass was greater than grass biomass on plots seeded with grass 2 or 3 years after seeding with sagebrush [64]. Once established, big sagebrush may become dense and reduce grass biomass [21] (see Successional status). See Value for rehabilitation of disturbed sites for more information on seeding mountain big sagebrush.
Reducing nonnative plants may increase mountain big sagebrush establishment. Mountain big sagebrush stands with abundant cheatgrass and bulbous bluegrass in north-central Utah were tilled in fall after cheatgrass green-up. Some mountain big sagebrush plants were left in tilled areas. The following spring, mountain big sagebrush seedlings were observed on tilled plots, while no mountain big sagebrush seedlings were observed in untilled areas. Sites that were tilled in fall prior to cheatgrass green-up also had no mountain big sagebrush seedlings the following spring, likely because cheatgrass could establish before mountain big sagebrush [661].
Establishment of mountain big sagebrush seedlings is infrequent in dense stands of mountain big sagebrush [355,403,628], and occurs mainly in canopy openings as mature plants break down or die [355]. On the Beaverhead National Forest, Montana, no seedlings were present in a dense mountain big sagebrush stand when it was 31 years old and had 167 mature plants/30 ft² or when the stand was 47 years old and had 114 mature plants/30 ft². However, 5 mountain big sagebrush seedlings were present when the stand was 51 years old and beginning to have openings, with 93 mature plants/30 ft² [355] (see Successional status). In Utah, survival of mountain big sagebrush seedlings on a mesic site and Wyoming big sagebrush seedlings on a dry site were both more negatively impacted by competition from adult big sagebrush plants than by competition from either bluebunch wheatgrass or nonnative crested wheatgrass [522].
Self-thinning of big sagebrush seedlings occurs where seedlings are closely spaced. After artificial seeding on disturbed sites, only about 10% to 15% of seedlings survive due to self-thinning (reviewed in [436]). On a seeded mine site in Nevada, survival of 1-year-old big sagebrush seedlings was higher when seeds were well dispersed than when they were close together, suggesting that much of the first-year seedling mortality was due to self-thinning of closely spaced seedlings [434]. Self-thinning of big sagebrush seedlings and young plants occurs over a 5- to 10-year period after seeding. Approximately 42,000 big sagebrush seedlings and young plants/ha established and persisted for 2 to 3 years after seeding on disturbed sites in southern Idaho and Wyoming. After 10 years, big sagebrush density was about 2,000 plants/ha [436].
Postfire seedling establishment: Mountain big sagebrush establishes from seed from soil seed banks and unburned plants (on-site or off-site) after fire [417,447,628,767,768]. Because seed in the soil is relatively short-lived and susceptible to mortality from fire, and seeds disperse relatively short distances, distribution of seed-producing plants is an important driver of mountain big sagebrush postfire establishment. During the first few postfire growing seasons, seeds in the soil and from unburned plants in the burn interior and perimeter are most important for mountain big sagebrush seedling establishment. As succession proceeds, mountain big sagebrush individuals that established soon after fire mature and contribute seed for subsequent establishment [139]. After the first few postfire years, mountain big sagebrush seedling establishment slows because of depleted soil seed banks [767] and increased competition for resources with grasses and forbs [65,177]. In southern Oregon and northwestern Nevada, most mountain big sagebrush recruitment occurred from the soil seed bank in the first and second growing seasons after fire on four large burns. This initial peak in establishment was followed by several years of low or no recruitment because soil seed banks were depleted. A second peak of establishment occurred around 7 to 10 years after fire, presumably when established plants reached maturity and provided an on-site seed source [767,768]. Mountain big sagebrush densities increased following cutting and burning treatments to remove western junipers from curlleaf-mahogany/mountain big sagebrush/Idaho fescue communities. Establishment occurred in two pulses: the first pulse occurred the first or second year after treatment and was likely from the seed bank, and the second establishment pulse occurred the fourth year after treatment and was likely from seed produced by individuals established during the first pulse and from residual shrubs in treatments without broadcast burning [471].
Several studies show most mountain big sagebrush seedling establishment occurs within the first 4 years after fire [65,177,460,664,767]; however, the timing of peak postfire establishment varies among sites, and postfire seedling establishment may be absent or limited on some sites for many years (e.g., [228,230,249,267,281,420,681,698,767]), especially when grasses and forbs establish at high densities (e.g., [267,398,486]) (see Interference and competition). At four sites near Dubois, Idaho, mountain big sagebrush seedlings established 1 to 3 years after fall prescribed burning, and none established 4 to 8 years after burning [177]. Near Reno, Nevada, mountain big sagebrush started to establish 3 years after a 1957 wildfire, but peak establishment did not occur until 13 to 18 years after the fire, as the previously established plants matured and produced seed. Establishment declined thereafter [763]. Mountain big sagebrush cover was 11% before a prescribed fire but still absent 2 and 4 years after the fire in a Colorado pinyon-Utah juniper community in northeastern Utah [281].
Establishment of mountain big sagebrush can be high immediately following disturbances that remove vegetation [111,177,267,391,420,518,767], but establishment can also be low [267,681]. For example, 1 year after a patchy September prescribed fire in a high-elevation (7,200 feet (2,200 m)) shrub-grass community in western Wyoming, mean density of mountain big sagebrush seedlings on an eastern slope was higher in burned (1,090/acre (2,691/ha)) than unburned (97/acre (240/ha)) plots [518]. In contrast, on seven burns ranging from 9 to 13 years old on the Northern Yellowstone Winter Range, mean density of small mountain big sagebrush plants (6 years old, on average) was 0.08 plant/m² compared to 0.91 plant/m² on unburned areas (P = 0.04), suggesting better recruitment on unburned areas, perhaps due to interference from grasses on burned sites. Grass cover on burned sites (53%) was greater than that on unburned sites (36%, P ≤ 0.03) [267,681] (see Interference and competition).
Plant growth and mortality: Big sagebrush appears to grow best in full sun when moisture is plentiful. It tolerates shading, but grows less in shade than sun [233] (see Successional status).
Mountain big sagebrush seedlings in greenhouses and common gardens grow rapidly [73,207,761]. Six-month-old seedlings in greenhouse studies averaged 15 inches (37 cm) tall [73], and roots averaged 27 inches (68 cm) long [704]. Stems of mountain big sagebrush plants grown in a common garden for 4 months from seeds collected from Provo, Utah, grew 9.2 inches/year (23.3 cm/year) [207].
Although some researchers were unable to differentiate the rate of growth of seedlings among the three major big sagebrush subspecies [251], in general, early shoot growth rates of mountain big sagebrush appear to be faster than those of Wyoming big sagebrush but slower than those of basin big sagebrush [73,207,388,400,403,695,707]. These differences in growth rates parallel differences in absolute size at maturity, with basin big sagebrush the tallest, mountain big sagebrush intermediate, and Wyoming big sagebrush the shortest [403,695]. Slower growth of mountain big sagebrush than basin big sagebrush based upon age-radius relationships of stems in wild populations in south-central Idaho was attributed to shorter growing seasons and higher shrub densities and possibly genetic characteristics [736].
Freezing temperatures can result in injury or death of nondormant mountain big sagebrush plants [235,247,669,700]. Mountain big sagebrush plants were killed and injured over large areas of Idaho, Nevada, Utah, western Wyoming, and western Colorado during the winter of 1976–1977, and virtually all large individuals were killed on high plateaus. Precipitation was about 31% of average from November through February, resulting in shallow snow, and temperatures between November and March fluctuated between above-average and below-average, breaking winter dormancy and resulting in freeze injury and mortality [247,456,457]. Areas with the most severe winter kill were those normally covered by more than 3.3 feet (1 m) of snow but lacked snow that winter [247]. Hardened big sagebrush short shoots and stems tolerated temperatures as low as -58 °F (-50 °C) in a laboratory; however, these same plants were killed at -4 °F (-20 °C) after 6 days at 68 °F (20 °C) [669].
Deep snow may favor mountain big sagebrush growth [511], although a late-melting snow pack shortens the growing season [496] and, in some areas, deep snow pack can be associated with parasitic snow molds [606,607,700]. Growth, as measured from ring widths in 5 mountain big sagebrush plants collected from the Gunnison Basin, Colorado, was positively correlated with maximum snow depth in winter (r = 0.74, P < 0.001) and spring (r = 0.71, P < 0.001), and was negatively correlated with temperature, particularly summer temperature (r = -0.62, P < 0.01) [511]. At a high-elevation (9,580 feet (2,920 m)) meadow in Gunnison County, mountain big sagebrush growth rate was negatively correlated with later snowmelt dates (adjusted r² = -0.45, P = 0.02) [496]. If snow is deep enough, parasitic snow molds can injure or kill mountain big sagebrush [606,607,700]. In Wyoming, when snow was <16 inches (41 cm) deep, an unidentified snow mold infected 2% of mountain big sagebrush plants, but when snow was >47 inches (119 cm) deep, it infected 93% of plants, reducing mountain big sagebrush canopy cover and killing some plants [607]. For a review of snow mold disease and other parasitic diseases of big sagebrush, see Welch [700].
Mountain big sagebrush growth and survival can be adversely impacted by precipitation extremes. Early or extended drought can be an important cause of big sagebrush mortality [12,403,703]. Extensive big sagebrush mortality from drought occurred during the 1930s in Idaho [484], Montana [11,187], and Wyoming [11]. On the other hand, mountain big sagebrush decline (i.e., slow growth, low seed production, shoot dieback, and occasional death) was recorded in Utah and attributed to a period of record-high precipitation from 1977 to 1986 [456].
Hot summer days may increase water stress and limit mountain big sagebrush growth. In common gardens in Utah, mountain big sagebrush and basin big sagebrush transplants grew best at relatively cool temperatures (≤59 °F (15 °C)), while both subspecies were stressed and grew more slowly at warm temperatures (≥86 °F (30 °C)). Basin big sagebrush seemed better able to withstand heat stress than mountain big sagebrush [583]. Increased summer temperatures predicted by global climate change models and changes in the amount and timing of precipitation will likely affect growth of mountain big sagebrush plants [511] (see Climate change).
Mountain big sagebrush plants seem to grow better when competition for resources with other vegetation is reduced. For example, mountain big sagebrush canopy cover declines with increasing overstory canopy cover [240,423,460] (see Successional status). In Idaho, dry mass gain and early season photosynthesis in mountain big sagebrush plants that established after fire were negatively correlated with proximity to neighboring herbs [176].
Herbivory by wildlife and livestock also influence growth and mortality of mountain big sagebrush. For more information, see Herbivory.
Vegetative regeneration: Mountain big sagebrush can reproduce by layering [54,55,255,383], but does not sprout from roots or root crowns after top-kill [65,66,461,485,628,758]. McArthur et al. [383] stated that mountain big sagebrush branches "frequently layer", but field observations in southern Montana suggest that layering was only "occasional" or "rare" on those sites [254]. While Johnson [299] stated that mountain big sagebrush "sprouts from the bases of plants when root crowns are not killed", most sources indicate that it does not sprout from the root crown or roots following top-kill by fire or other means [65,66,461,485,628,758].
Silver sagebrush sprouts, and its hybrids may also sprout. Frisina and Wambolt [217,678] stated that snowfield big sagebrush—a big sagebrush subspecies that may be a hybrid of mountain big sagebrush and silver sagebrush [231]—is the only big sagebrush subspecies known to root sprout. A natural plains silver sagebrush × mountain big sagebrush hybrid from Gallatin County, Montana, produced root sprouts in the field and layered when moved to a greenhouse [255].
SUCCESSIONAL STATUS: Shade tolerance: Mountain big sagebrush grows best in full sun [233,589,722]. Its cover declines as tree cover increases (e.g., [240,413,423,460,540,545,553,589]) and increases when tree cover is removed [540,580,619,745]. In the Great Basin and Colorado Plateau regions of eastern Nevada and Utah, the presence of trees (tree cover ranged from 1%-35%) was negatively associated with mountain big sagebrush cover and explained 48% of the variance among 18 sites where trees had established within mountain big sagebrush communities (P < 0.001) [460]. Mountain big sagebrush cover was negatively correlated with Douglas-fir canopy cover (r = –0.86, P < 0.001) at 25 sites in three mountain big sagebrush communities in southwestern Montana. Mountain big sagebrush canopy cover declined to <15% when Douglas-fir canopy cover increased beyond 20%. Mountain big sagebrush canopy cover declined to <5% when Douglas-fir canopy cover increased beyond 35% [240]. In southeastern Oregon and northeastern California, mountain big sagebrush cover in mountain big sagebrush/Thurber needlegrass, mountain big sagebrush/Idaho fescue, and mountain big sagebrush-mountain snowberry/Columbia needlegrass associations declined as western juniper cover increased (r² = 0.92-0.96, P < 0.05). Herbaceous cover and species diversity also declined, and bare ground cover increased, with increasing western juniper cover [423]. In central Oregon, big sagebrush (mountain big sagebrush, Wyoming big sagebrush, or both) biomass increased 1 year after ponderosa pine was thinned and western juniper was removed [540].Shrubland succession: Mountain big sagebrush is a late-seral dominant or "climax" species in steppe communities (e.g., [54,131,263,342,439,442]) that can persist as a habitat dominant in late-seral stages in the absence of fire or other large-scale disturbances [196,267,355,680]. Important large-scale disturbances in mountain big sagebrush communities include fire, herbivory, freeze-kill, snow mold, and drought [196]. In the absence of disturbances, mountain big sagebrush communities may become dense [196,197,355], and stands adjacent to woodlands may succeed to woodland (e.g., [54,131,239,242,267,355,398,442,446,640,680]).
Postfire successional patterns in sagebrush communities are considered "predictable", although the composition of postfire communities and rates of postfire succession vary considerably [197,239] depending on numerous interacting variables, including prefire plant community and seed bank composition; site characteristics and management history; fire size, severity, and patchiness; and postfire weather [415,416,768] (see Plant response to fire). Typically, annual herbaceous plant cover increases immediately after fire. Perennial grasses, forbs, and sprouting shrub species, if present, then increase and dominate for up to 20 years. Mountain big sagebrush may establish early in postfire succession from seed in the soil seed bank and from seed dispersed from surviving plants in unburned patches and the burn perimeter [417,447,628,767,768]. Most mountain big sagebrush seedling establishment occurs within the first 4 years after fire [65,177,460,664,767], although it is influenced by several factors. After the first few postfire years, mountain big sagebrush seedling establishment typically slows because of depleted soil seed banks [767] and increased competition for resources with grasses and forbs [65,177].
Because mountain big sagebrush must establish from seed and has slower growth rates, it dominates the postfire plant community much later in succession than grasses, forbs, and sprouting shrubs [54,239]. Following the initial postfire establishment period, secondary peaks in establishment occur when mountain big sagebrush individuals that established soon after fire mature and contribute seed to subsequent establishment [139]. Thereafter, establishment may be episodic [497,498] (see Seedling establishment). Canopy cover of mountain big sagebrush in mountain big sagebrush communities typically increases after fire or other disturbances. As mountain big sagebrush cover increases during succession, cover of grasses, forbs, and other shrub species declines [7,54,239,418,448,461,720]. Shrub canopy cover in undisturbed mountain big sagebrush communities typically varies between 15% and 40% but can reach up to 50% or more [417] (see Stand structure). On moderately deep to deep loamy soils in Carbon County, Wyoming, Warren [686] categorized mountain big sagebrush communities with <5% mountain big sagebrush canopy cover as early seral, 5% to 40% canopy cover as mid-seral, and >40% canopy cover as late seral, with mid-seral communities supporting the highest numbers and diversity of wildlife (see Considerations for wildlife management). On moderately deep to deep loamy soils in southeastern Oregon, Evers [196] identified four general successional phases as follows: 1) early-seral phase: grasses and forbs dominant with mountain big sagebrush seedlings present (<1% mountain big sagebrush cover); 2) mid-seral, open phase: grasses and forbs dominant with mountain big sagebrush subdominant (1%-10% mountain big sagebrush cover); 3) late-seral, open phase: mountain big sagebrush, grasses, and forbs codominant (10%-30% mountain big sagebrush cover); and 4) late-seral, closed phase: mountain big sagebrush dominant (>30% mountain big sagebrush canopy cover).
The time required for mountain big sagebrush communities to develop late-seral attributes is influenced by a number of factors and therefore varies substantially among sites. To model succession in mountain big sagebrush communities in southeastern Oregon, Evers [196] assumed that a mountain big sagebrush population began with a single plant and doubled every 2 years and that it took 6 years for a seedling to reach reproductive maturity. The modeled communities reached the late-seral, closed phase in about 31 years in the absence of disturbance. When disturbance effects from insects, voles, freeze-kill, snow mold, drought, and fire were added to the model, it took 33 years to reach that phase [196]. Our review and analysis of mountain big sagebrush postfire recovery data showed that few sites had fully recovered (i.e., had mountain big sagebrush canopy cover that was equal to or greater than that on similar, unburned sites) before 25 years. While most sites studied (67%) had fully recovered after 25 years, sample size was small and variability was high for burns >25 years old, and about a third of them had not fully recovered. Computations of postfire recovery are complicated by variability in mountain big sagebrush canopy cover on unburned sites, which ranged from 4% to 71% (see Plant response to fire for details) and is influenced by a number of variables, including site characteristics, land use history, and successional stage.
Fire or other disturbance is not necessary to maintain mountain big sagebrush stands in a "relatively young, productive state" or to prevent them from becoming "decadent". Mean plant age of mountain big sagebrush on 33 relatively undisturbed sites on the Northern Yellowstone Winter Range, where vegetation cover averaged 14% (range: 3%-39%), was 32 years, with only 12% of individuals >50 years old and 5% >60 years old. The average age of dead plants was 41 years old. The author concluded that mountain big sagebrush plants established, matured, died, and were replaced within about 50 years [267,680]. A 30-year study of a mountain big sagebrush stand in the Gravelly Mountains of southwestern Montana, showed that mountain big sagebrush stands may persist indefinitely in the absence of large-scale disturbances. Based upon observations of the stand from 31 to 61 years after initial establishment, the author concluded that a dense stand of mountain big sagebrush "had to age sufficiently to break down into a more open stand before conditions were right for new plants to come in", and that "many plants died before the stand was open enough for the sunlight to reach the ground and permit seedlings to come in". No mountain big sagebrush seedlings were observed until 51 years after initial establishment [355]. In undisturbed stands, seedling establishment may be episodic [497,498] (see Seedling establishment).
Woodlands can expand into mountain big sagebrush communities when the interval between fires becomes long enough for trees to establish, mature, and dominate a site [419]. However, in Utah and adjacent Nevada, Kitchen and Weisberg [320] noted that conifer establishment in mountain big sagebrush communities was "minimal" on many sites even after apparently long periods without fire. Conifer expansion is most common in sagebrush communities on cool to warm, relatively moist sites (table 7, figure 18), including mountain big sagebrush communities at high elevations [303,416,419] (see Woodland expansion). Where soil temperature and moisture regimes are suitable, mountain big sagebrush and other sagebrush plants can act as nurse plants, providing safe germination and establishment sites for junipers and pinyons [109,185,240,421].
Miller et al. [413,426] categorized woodland succession in mountain big sagebrush communities in three phases:
A fourth late-successional, closed stage or "mature" phase, is sometimes included in woodland succession where trees are dominant, shrubs and herbaceous plant cover is minimal or absent, and shrubs are >90% dead [423,430]. The period of time required to transition between phases in woodland succession is variable. In southeastern Oregon and southwestern Idaho, the development of western juniper woodlands in mountain big sagebrush and low sagebrush communities from the time of tree establishment to dominance (shift from Phase I to Phase III) varied from 80 years on cool, moist sites to >120 years on warm, dry sites [303]. Based on tree age chronology, the minimum time for western juniper stands to approach dominance on mesic mountain big sagebrush/Idaho fescue sites in Oregon and California was 60 to 70 years and nearly 80% of western juniper trees established within a 30-year period [422,423]. Based on this and other data, Johnson and Miller [303,416] developed a chart for mountain big sagebrush communities with varying productivity that hypothesizes the time necessary to transition from initial western juniper establishment to development of late-seral woodlands (figure 3). The transition from mid-seral to late-seral woodlands causes a shift from shrub and herbaceous fuels to a predominance of tree canopy fuels, which influences fire behavior and severity [178,416,425,539,750,765] (see Fuels).
Numerous state-and-transition models have been proposed that describe successional processes—including the establishment and spread of junipers and pinyons into mountain big sagebrush communities—and model community transitions following natural and human-caused disturbances in mountain big sagebrush communities (e.g., [43,121,122,195,258,300,503,600,622,717,721]). For more information, see State-and-transition models.
 |
| Figure 3—Hypothesized time periods from initial western juniper establishment (early Phase I) to development of late-successional woodland (Phase III), and estimated maximum density and canopy cover of western juniper for stands developing on mountain big sagebrush sites with varying elevation and insolation exposure (i.e., a gradient of relatively cool/moist to warm/dry sites) [303,416]. |
Woodland succession: Mountain big sagebrush occurs in all successional stages of pinyon-juniper woodlands, typically establishing soon after fire and other disturbances, increasing in size and density for several decades, then declining as tree cover increases. Plant succession following fire in western juniper-mountain big sagebrush woodlands generally starts with an annual stage, followed by a perennial grass-forb stage, a perennial grass-forb-shrub stage, a perennial grass-forb-shrub-young juniper stage, a shrub-juniper stage, and finally, juniper woodland [35]. Five postfire successional stages were described for burned singleleaf pinyon-western juniper/mountain big sagebrush woodlands in California and Nevada, each with a representative cover of mountain big sagebrush: early succession (1-year-old burns, 0% cover mountain big sagebrush), early midsuccession (4- to 8-year-old burns, 15% cover), midsuccession (15- to 17-year-old burns, 36% cover), late midsuccession (22- to 60- year-old burns, 88% cover), and late succession (>60-years-old, 30% cover) [329]. Wright et al. [745] reported that shrubs such as big sagebrush and broom snakeweed begin establishing around the sixth year after fire in pinyon-juniper woodlands and gradually become denser. Trees begin to overtop them after about 100 years, and shrub growth is gradually suppressed as the woodland matures. After about 300 years, the pinyon-juniper forest has an understory composed of a few scattered shrubs and perennial and annual grasses [745]. Timing of tree dominance varies, but some estimates and observations suggest 30 to 50 years for western juniper [98], 46 to 71 years for Utah juniper [35], and ~60 years for pinyon [94,102,620]. See FEIS Species Reviews for information on postfire succession in pinyon-juniper communities.
Forest Succession: Mountain big sagebrush typically establishes early in succession in a variety of forest communities, such as Douglas-fir, limber pine, lodgepole pine, ponderosa pine, and subalpine fir communities (see Plant communities), often within a few years after fire or other disturbance (e.g., [53,54,263,300,589,590,640,734,736]). Its cover and density peak prior to shading by taller shrubs and trees. In late succession, when the tree canopy is closed, mountain big sagebrush is sparse or absent [553,590]. In Idaho, mountain big sagebrush is a common early-seral shrub in Douglas-fir communities, declining in later succession. The phase dominated by mountain big sagebrush may be "relatively short lived" [590], although regeneration lags or frequent fire may result in "semi-persistent" mountain big sagebrush dominance in some areas [29,492,640]. For example, some sagebrush-dominated areas in the Bighorn Mountains, Wyoming, may have resulted from successive high-severity fires in Rocky Mountain Douglas-fir stands [492]. In the limber pine series in Wyoming and Idaho, mountain big sagebrush may be dominant in early postfire succession, but is gradually shaded out by limber pine [589]. In late succession, mountain big sagebrush may persist in forest canopy openings [292,504], and its cover and density may increase in old-growth forest as openings are created by dying trees in the forest canopy [396,591].
| FIRE EFFECTS: |  |
| Figure 4—Fire in Kennedy Meadows, California, with mountain big sagebrush in foreground. Photo courtesy of Wildlandfire.com. |
Immediate fire effects:
Immediate effects on plants: Mountain big sagebrush branches are low to the ground; the wood, bark, and foliage are highly flammable [90,448,462,703]; and plants are highly susceptible to injury from fire [7,28,65,66,100,131,162,391,462]. Fire typically kills mountain big sagebrush; thus, its cover and density are reduced by fire (e.g., [7,28,65,333,349,391,518,664]). Clark and Starkey [131] stated that a fire intense enough to scorch the foliage kills big sagebrush plants. In Fremont and Clark counties, Idaho, mountain big sagebrush was nearly always killed by fire, regardless of severity rating: low-severity fires consumed sagebrush leaves and left small branches and twigs unburned; moderate-severity fires consumed small branches and twigs, but large branches were unburned; and high-severity fires consumed the entire sagebrush plant, including the trunk and main stem. Even after low-severity fires, many of the mountain big sagebrush plants that initially appeared to survive were actually injured "beyond recovery" [65]. Low-severity fires tend to leave patches of unburned vegetation, and more mountain big sagebrush plants survive on low-severity than high-severity burns [65,486]. Following a spring prescribed fire in a mountain big sagebrush/Idaho fescue community in central Idaho, mean density of mountain big sagebrush plants was reduced from 30 plants/20 m² to 8 plants/20 m², and mean canopy cover was reduced from 26% to 6% in low-severity burn patches. In high-severity burned patches, only a few scattered mountain big sagebrush plants survived [333].
Immediate effects on seeds: Fire can kill many mountain big sagebrush seeds in the soil seed bank [10], but some seeds may survive [10,124,125]. Mountain big sagebrush seeds lack a thick seed coat or any other adaptations to survive fire [703], and they have hypocotyl hairs that help facilitate germination and seedling establishment on the soil surface (Young and Martens 1991 cited in [34]), where they are exposed to higher temperatures during fires [703].
Few data are available for quantitatively assessing fire effects on mountain big sagebrush seed banks. Mountain big sagebrush seed anatomy, dispersal characteristics, and postfire seed bank studies indicate that fire is likely to reduce the abundance of germinable mountain big sagebrush seeds in soil seed banks [10,703,760], although one frequently cited study suggested that fire may stimulate germination of mountain big sagebrush seeds in the soil [125]. Mountain big sagebrush emergence in a greenhouse was lower in soils collected from unburned plots (12 seedlings/m²) than in soils collected from either low-severity (32 seedlings/m²) or high-severity (20 seedlings/m²) burned plots (P < 0.05) that were burned in situ with a propane burner prior to collecting 2-inch (5.1 cm) deep soil samples. The low-severity burn reached a maximum soil surface temperature of 219 °F (104 °C) after 30 seconds, and the high-severity burn reached a maximum soil surface temperature of 781 °F (416 °C) after 60 seconds [124,125]. Nonetheless, other evidence suggests that mountain big sagebrush seeds in the soil are likely to be injured or killed by wildfire.
Most mountain big sagebrush seeds are located in the litter under parent shrubs [255,661,760] (see Seed dispersal). This litter is typically consumed during fire, and the soil surface can reach lethal temperatures even during low-severity fire [10]. In a singleleaf pinyon-Utah juniper/mountain big sagebrush community in central Nevada, mountain big sagebrush seed density in soils declined by 78% after an early May prescribed fire. One week prior to burning, mountain big sagebrush seeds comprised 44% of the germinable seed bank, and most (90%) were located under adult mountain big sagebrush shrubs. Maximum temperature at the soil surface under shrubs reached 696 °F (369 °C) during the fire, and litter under shrubs was largely consumed. One week after burning, mountain big sagebrush comprised 9% of the germinable seed bank, and most seeds were still in microsites under shrubs. Two years after burning, mountain big sagebrush comprised 2% of the germinable seed bank in burned areas in fall and 15% in unburned areas. The authors noted that a higher severity wildfire in late summer or fall would likely result in greater reductions of the germinable seed bank [10].
Large, severe wildfires may leave few to no germinable mountain big sagebrush seeds in soil seed banks [461]. No mountain big sagebrush seedlings emerged from 2.5-inch (6.4 cm) deep samples of litter and soil collected from burned areas in September after a large August wildfire near Reno, Nevada. An average of only 0.001 mountain big sagebrush seedling/m² emerged in 25 plots in the burned area at the end of the first postfire growing season, indicating that either there was a very small reserve of viable seeds in the soil after the fire or that animals dispersed some seeds to the burn. Few details on the fire were provided, except that the fire consumed all standing woody material [760]. Fourteen years after the Grandine Fire in Idaho, Welch and Criddle [703] noted no mountain big sagebrush seedlings in the interior of a large burn.
Fire effects on seeds depend on location of seeds, including depth of burial, and the magnitude and duration of soil temperatures reached during fire, which are spatially heterogeneous due to spatial variability in fuel loads [41,330]. Buried seeds are usually more protected from lethal temperatures during fire than seeds in the litter or on the soil surface [51,226]. Based on the data in table 6, Miller et al. [416] concluded that mountain big sagebrush seeds in soil seed banks may be least likely to survive fire in pinyon-juniper woodlands and that seeds located on the soil surface or in the litter have a high probability of being consumed or exposed to lethal temperatures during fire. Lethal temperatures for plant tissues generally range from about 104 to 158 °F (40-70 °C), although some seeds can survive exposure to higher temperatures. Lower temperatures can be lethal if tissues are exposed for a longer duration (e.g., [288,350,658]).
| Table 6—Average and peak soil surface and subsurface temperatures measured in big sagebrush, cheatgrass, and pinyon-juniper communities in the western United States. | |||||
| Location | Plant community | Fire characteristics | Site characteristics | Average soil surface temperatures, unless otherwise noted | Source |
| Idaho | mountain big sagebrush | October prescribed fire | tree or shrub interspaces | 174-399 °F (79-204 °C) | [41] |
| tree canopy litter | 399-1,292 °F (204-704 °C) | ||||
| Nevada | mountain big sagebrush and Wyoming big sagebrush | October and mid-November prescribed fires | bare ground | 590 °F (310 °C) | [330] |
| under grasses | 585 °F (307 °C) | ||||
| under shrubs | 718 °F (381 °C) | ||||
| Utah | bare ground | 487 °F (253 °C) | |||
| under grasses | 570 °F (299 °C) | ||||
| under shrubs | 639 °F (337 °C) | ||||
| Utah | double-chained and seeded pinyon-juniper community | September prescribed fire | under grasses | 369 °F (187 °C) | [226] |
| <131 °F (55 °C) at 1.0-inch (2.5 cm) deep |
|||||
| under juniper and pinyon debris pile | >1,431 °F (777 °C) | ||||
| >550 °F (288 °C) at 1.0-inch (2.5 cm) deep |
|||||
| Utah and Washington | "near monocultures" of cheatgrass in big sagebrush steppe | June and July prescribed fires | under grasses | 248-293 °F (120-145 °C) peak temp on surface |
[51] |
| 140-167 °F (60-75 °C) peak temp at 0.4-inch (0.1 cm) deep |
|||||
Postfire regeneration strategy [598]:
Shrub without adventitious buds and without a sprouting
root crown
Ground residual colonizer (on site, initial community)
Initial off-site colonizer (off site, initial community)
Secondary colonizer (on- or off-site seed sources)
Fire adaptations: Mountain big sagebrush plants are poorly adapted to survive fire [533,678,698,738], but postfire seedling establishment can be abundant [65,111,177,391,420,471,486,518,532,664,763,767]. Mountain big sagebrush plants are easily killed by fire; they do not sprout [7,65,66,100,131,391,462,485,628]. Wildfires in mountain big sagebrush communities are typically stand-replacement fires that kill or top-kill most vegetation (e.g., [31,398,527,533,681,698,767,768]); however, fires can be patchy, due to variation in fuels, topography, and weather, and can result in a landscape mosaic of burned and unburned areas [31,415,601]. The soil seed bank and surviving plants in and adjacent to burns are important seed sources for postfire establishment [767]. Mountain big sagebrush seed production is highly variable, and depends on site and plant characteristics (e.g., size, age, and genetics) [339], but surviving plants may produce abundant seeds [43,161,255,692,713,763]. Most lightning-caused wildfires in sagebrush communities occur in summer and early fall before mountain big sagebrush seeds are ripe [767], so seeds from surviving plants are dispersed by wind, water, and animals [233,255,404,495,628,760] in fall and winter after wildfire [52,66,383,628,760] (see Seasonal development). Most mountain big sagebrush seeds fall within 10 feet (3 m) of parent plants [213,255,628,661,701,760]. Mountain big sagebrush seeds may also germinate from soil seed banks [447,729,767,768] if they survive fire and are not buried too deeply. Viable mountain big sagebrush seeds must be within 0.6 inch (15 mm) of the soil surface to germinate [290,760], where they are vulnerable to lethal temperatures during fire.
Hybrids: Observations of a plains silver sagebrush × mountain big sagebrush hybrid in the field show that at least some of these hybrids can root sprout after fire. The hybrid "showed more morphological characteristics of silver sagebrush", which is a strong sprouter [255].
Plant response to fire:Due to concerns regarding habitat requirements for sagebrush obligates, the length of time necessary for mountain big sagebrush canopy cover to return to prefire or unburned values (i.e., postfire recovery time) has been the focus of numerous studies (table A3). Postfire recovery time is determined by comparing mountain big sagebrush abundance (usually canopy cover, but also density and height) on burned sites to its prefire abundance or that on similar, unburned sites over time. This presumably indicates the time needed for big sagebrush to "regain full coverage and maturity" after fire [31]. Estimates of postfire recovery time are therefore strongly influenced by variability in cover values on unburned sites. Some of this variability derives from site characteristics (e.g., poor growing conditions leading to low potential canopy cover, or the reverse), while some may be due to site degradation from past land uses (e.g., livestock grazing and related management that decreased shrub cover, or lengthened fire-free intervals that increased shrub cover) [31]. Thus, the assumption that unburned sites consistently represent full recovery may be inaccurate for some sites. This is an important consideration because postfire recovery time is sometimes used to estimate fire frequency in mountain big sagebrush communities, based on the premise that mountain big sagebrush communities did not burn, on average, more frequently than the time required for them to recover [31,744]. See the FEIS synthesis on fire regimes of mountain big sagebrush communities for more details.
Analysis of postfire recovery studies:
| Our analyses found that mountain big sagebrush canopy cover and postfire recovery increased fairly consistently during the first 30 years after fire (figure 6), and that sites began reaching full recovery at around 26 to 30 years after fire, on average, when mountain big sagebrush canopy cover averaged around 28%. Mountain big sagebrush postfire canopy cover and postfire recovery may be faster on some sites and in some ecoregions than in others (figure A1); however, differences in unburned cover values (i.e., the recovery threshold) among study sites and in the number of study sites among ecoregions complicate comparisons of postfire recovery within and among ecoregions. |
In order to synthesize information on mountain big sagebrush postfire recovery time, we obtained data on mountain big sagebrush canopy cover and postfire recovery from 306 burned sites (figure 5) examined in 20 published and unpublished studies (table A3). We obtained most data directly from publications, but for seven studies we obtained site-level data directly from the authors [2,3,4,5,347,459,686]. In most studies, researchers compared mountain big sagebrush cover on burned sites to cover on nearby unburned sites with the assumption that unburned sites were representative of fully recovered sites. Burned sites ranged from 1 to 69 years since fire, with mean mountain big sagebrush cover values ranging from 0% to 57%. Unburned sites also had a wide range of mean cover values (4%-71%) and averaged 26% cover, overall [2,5,169,206,246,272,280,347,391,398,432,459,671,681,686,767]. Most unburned sites showed no evidence of fire. However, some "unburned" paired sites were actually burned sites with longer time-since-fire than their burned comparisons. For example, Nelson's [459] burned sites ranged from 3 to 40 years since fire, while nearby, paired unburned sites ranged from 17 to 164 years since fire. In four studies, postfire mountain big sagebrush cover was compared with prefire values [3,4,246,654]. For one study that examined 136 burned sites in the Uinta Mountains and Tavaputs Plateau [230], we used 25% canopy cover to represent full recovery based on the author’s observation that “the data show mountain big sagebrush strongly trending toward >25% crown cover in the absence of fire or other disturbance”.
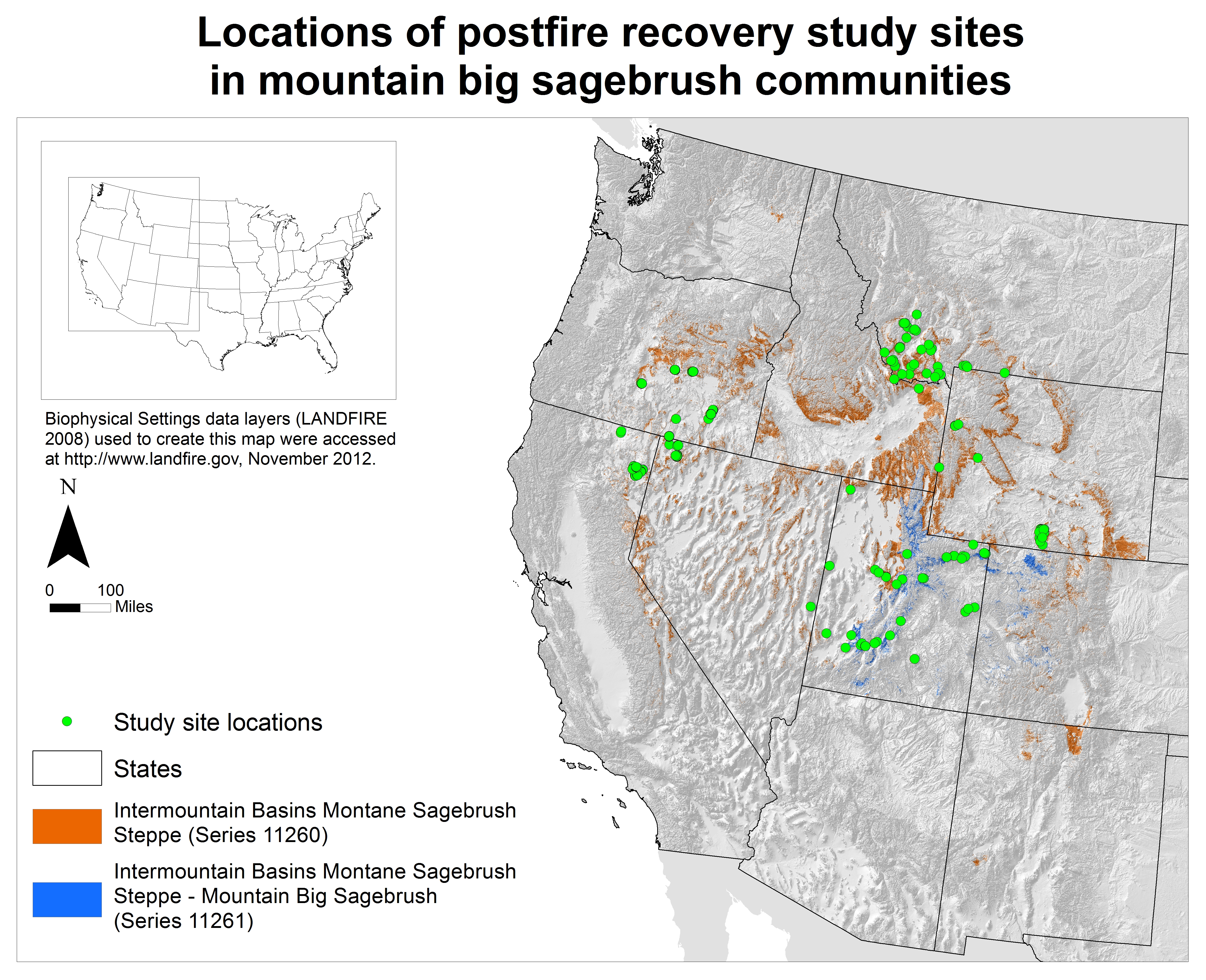 |
| Figure 5—Locations of postfire recovery study sites in mountain big sagebrush communities. In some cases, multiple study sites in close proximity (e.g., those of Moffet et al. [432] and Goodrich et al. [230]) are represented by a single point due to scale. Distribution of mountain big sagebrush Biophysical Settings is from LANDFIRE data [341]. Click on the map for a larger image. |
Methods: To assess changes in mean canopy cover and postfire recovery across all 306 sites over time, we:
Our analyses showed that, overall, mountain big sagebrush canopy cover increased consistently during the first 26 to 30 postfire years, then was more variable (figure 6A); a similar pattern was observed in some of the individual studies (e.g., [432,767]). At the US Sheep Experiment Station, for example, mountain big sagebrush cover increased with time-since-fire for the first 18 years, but after 18 years, cover was unrelated to time-since-fire. Over the 1- to 69-year postfire chronosequence, cover increased by 0.9% during the first postfire year, by 2.9% from postfire year 17 to 18, and then it plateaued at 31% during subsequent years (i.e., the "steady-state" phase of recovery) [432]. In our analysis, high variability in canopy cover on burns older than 26 to 30 years was influenced, in part, by small sample sizes of burns in those age classes. Of the 306 burned sites examined in our analysis, only 43 (14%) were >25 years old, and cover on those sites ranged from 7% in a 32-year-old burn to 57% in a 64-year-old burn.
Like canopy cover, postfire recovery generally increased with time-since-fire up to around 30 years, and then became more variable (figure 6B). When canopy cover values were averaged within 5-year time-since-fire bins, full recovery began around 26 to 30 years after fire, on average, when mountain big sagebrush canopy cover averaged around 28%. Few burns (10%) had recovered within 25 years, but most burns (67%) had recovered after 25 years. Of the 263 sites <25 years old, only 27 had recovered; five of these recovered sites had relatively low canopy cover (<20%) for their unburned comparisons [272,391,459], which might explain their rapid recovery. Of the 43 burns >25 years old, 29 had recovered. Of the 14 burns >40 years old, 12 had recovered, and the other two were 60% recovered after 41 years [767] and 87% recovered after 67 years [432].
On average, a given site has a low chance of recovery (12%) within 15 years, a 50% chance of recovery in 29 years, and a high chance of recovery (95%) in 49 years (binary logistic regression, P < 0.0001) (figure 7); however, the certainty around these probabilities varies. Based upon 95% confidence intervals (CI) shown by the gray area in figure 7, uncertainty of recovery is greatest for sites about 25 to 50 years after fire, due to a similar number of recovered and unrecovered sites in that age range. For example, at 40 years, the average chance of recovery is 84%, but the 95% CI ranges from about 68% to 94%. Overall, the model was better at predicting unrecovered sites (n = 250) than recovered sites (n = 56), an influence of the substantially larger number of unrecovered sites that had less variability in time-since-fire. Of the 250 unrecovered sites, time-since-fire averaged 12 years (range = 66 years), and postfire canopy cover averaged 6% with a low of 0% and a high of 47%. Of the 56 recovered sites, time-since-fire averaged 31 years (range = 91 years), and postfire canopy cover averaged 31% with a low of 8% and a high of 57%.
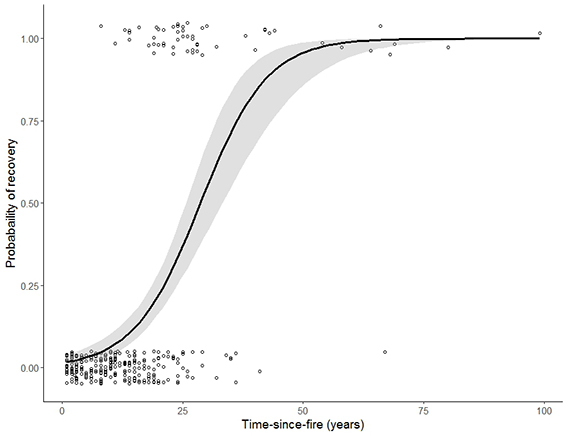 |
| Figure 7—Binary logistic regression analysis of mountain big sagebrush postfire recovery as a function of time-since-fire (n = 306 burned sites). Circles around the "1" line on the y-axis indicate recovered sites, and circles around the "0" line indicate sites that had not recovered to unburned canopy cover. The solid line represents the probability function derived from the prediction equation, and the gray area shows the 95% confidence interval. |
Considering the widespread distribution and importance of mountain big sagebrush communities, postfire recovery studies are limited geographically. Mountain big sagebrush steppe communities, as represented by LANDFIRE's Biophysical Settings [340], occur in 21 of the Level III ecoregions mapped by Omernik and Griffith [475] (figure 5). Postfire recovery data were available from eight of these ecoregions, although the number of burned study sites in each varied from nine in the Eastern Cascades Slopes and Foothills to 153 in the Wasatch and Uinta Mountains, and only three ecoregions had more than 20 study sites each.
When mountain big sagebrush canopy cover and postfire recovery were plotted against time-since-fire for each ecoregion, postfire recovery appeared to be faster in some ecoregions than in others (figure A1). However, differences in number of study sites and unburned cover values (i.e., the recovery threshold) make it difficult to compare recovery among ecoregions. For example, postfire recovery appears faster in the Snake River Plain than in other ecoregions because canopy cover of mountain big sagebrush was greater than unburned cover (23%, based on only one site) on 11 of the 12 burns >15 years old (figure A1.a). However, if 31% canopy cover (the point where postfire canopy cover plateaus on these sites [432]) was used to represent full recovery rather than 23%, then only 7 of 12 burns >15 years old would be considered recovered. In contrast, it appears that sites in the Wyoming Basin were relatively slow to recover because none of the sites had reached unburned cover values even 25 years after fire. However, because these sites were highly productive, unburned cover values averaged 56%, and most sites (60%) reached or exceeded 20% cover between 13 and 25 years after fire (figure A1.c). These results further emphasize the influence of unburned cover values on estimates of postfire recovery time and highlight the importance of examining site characteristics (e.g., site productivity) and land uses that may lead to variability in cover among unburned sites used for estimating postfire recovery. Regardless of postfire recovery estimates, mountain big sagebrush canopy cover appears to reach adequate levels to provide suitable winter habitat for sage-grouse (20% canopy cover [23,141,150,535]) around 16 to 20 years after fire on sites in the Snake River Plain and 11 to 15 years after fire on sites in the Wyoming Basin, on average (figure 8), assuming that sagebrush is tall enough to reach above snow depth [25].
Sites in the Middle Rockies appeared slowest to recover (figure A1). Only 1 of 63 burned sites had recovered, and only 15 sites ever exceeded 5% canopy cover within 35 years after fire (figure A1.b). Unburned canopy cover averaged 24% on these sites, which is similar to most ecoregions and suggests similar site productivity. Thus, slow recovery, overall, was not driven by high unburned cover values (i.e., highly productive comparison sites, as in the Wyoming Basin). Rather, several studies attributed the very low (<2%) mean mountain big sagebrush canopy cover up to 19 years after fire in and around Yellowstone National Park [267,398,654,681] to heavy postfire browsing by wild ungulates [267,398,681].
 |
| Figure 8—Mean mountain big sagebrush canopy cover by ecoregion versus time-since-fire in 5-year bins. The 20% canopy cover value is highlighted to represent typical sagebrush cover in sage-grouse winter habitat [23,141,150,535], and the 26% canopy cover value is highlighted to represent the average mountain big sagebrush canopy cover on unburned sites across all sites and ecoregions (n = 306). |
Not all sites in the Middle Rockies ecoregion were slow to recover. On one site, for example, mountain big sagebrush reached 20% canopy cover 15 years after fire. This site appeared to have a sprouting hybrid of mountain big sagebrush, which would lead to relatively faster postfire recovery. The identity of the hybrid was not determined, but silver sagebrush occurred nearby [1] and is known to sprout following top-kill and to hybridize with mountain big sagebrush (see Vegetative regeneration). These results emphasize the importance of prefire plant community composition as well as prefire canopy cover in estimating postfire recovery times. While this site was not considered recovered because mean unburned cover was 30% on the paired unburned site [2], it had adequate cover to provide suitable winter habitat for greater sage-grouse [1] (figure 9).
 |
| Figure 9—Greater sage-grouse winter habitat in a mountain big sagebrush community 15 years after a fall prescribed fire on Blacktail Butte, Grand Teton National Park, Wyoming. Photo courtesy of Diane Abendroth. |
In a similar analysis, Baker [31] reviewed 11 postfire recovery studies with cover and density data from >56 mountain big sagebrush sites in the Middle Rockies and Snake River Plain. He found that 16 sites showed nearly full recovery 25 to 30 years after fire; he classified these as "fast track" sites. The remaining sites were not yet recovered up to 35 years after fire. Baker estimated that those >40 sites would require 75 or more years to reach full recovery; he classified these as "slow track" sites. While acknowledging that postfire recovery time may be best represented as a continuum rather than just two tracks, he suggested that fast track recovery was more likely on relatively small burns, burns with high survival of mountain big sagebrush seed, and burns on sites with relatively more precipitation. He suggested that slow track recovery was more likely on large, uniformly high-severity burns, particularly if seed survival was low and seed dispersal from distant unburned areas was required for postfire establishment. Baker [31] did not examine differences between the ecoregions, and he used different criteria for inclusion of studies in his analyses than we did, so his results are not directly comparable to ours. Only six studies [206,280,349,398,671,681] were common to both analyses, and Baker included two studies [287,455] in his analyses that we could not use because site-specific canopy cover and time-since-fire values were not given. He also included three studies that reported density values that we did not use because our focus was on canopy cover.
Overall, our analyses of mountain big sagebrush recovery indicate a high degree of variability and uncertainty regarding postfire recovery rates. The following sections discuss factors that influence postfire recovery including prefire plant community and seed bank composition; fire characteristics such as fire severity, season, pattern, and size; postfire weather; and postfire herbivory [415,416,768].
Vegetation and site characteristics: Cold and cool, relatively moist sites characterized by mountain big sagebrush and mixed mountain shrubland communities tend to be more resilient to fire and other disturbances and more resistant to postfire nonnative annual grass invasion than warm, dry sites typically characterized by Wyoming big sagebrush communities (table 7, figure 10) [122,414,415]. This is because sites with greater moisture availability are more favorable for native plant growth and reproduction, and because nonnative annual grasses—primarily cheatgrass—are more invasive on warm, dry sites. Cool, moist sites typically occur at higher elevations than warm, dry sites, but this relationship is modified by slope and aspect, due to their influence on soil temperature and moisture regimes [121,122,416].
| Table 7—Sagebrush ecological types and their resilience to disturbance and resistance to nonnative annual grass invasion ([122], modified from [414,415]). Ecological types characterized by mountain big sagebrush are highlighted in yellow. | ||||
| Ecological type (soil temperature/moisture regimes) |
Mean annual precipitation | Typical shrubs | Resilience to disturbance | Resistance to nonnative annual grass invasion |
| Cold and moist (cryic/xeric) |
>14 inches (356 mm) |
mountain big sagebrush, snowfield big sagebrush, snowberry, serviceberry, silver sagebrush, and/or low sagebrush | Moderately high. Precipitation and productivity are generally high. Short growing seasons can decrease resilience on the coldest sites. | High. Climate suitability for nonnative annual grasses is low. |
| Cool and moist (frigid/xeric) |
12-22 inches (305-569 mm) |
mountain big sagebrush, antelope bitterbrush, snowberry, and/or low sagebrush (potential for juniper and pinyon expansion in some areas) | Moderately high. Precipitation and productivity are generally high. | Moderate. Climate suitability for nonnative annual grasses is moderate. |
| Warm and moist (mesic/xeric) |
12-16 inches (305-406 mm) |
mountain big sagebrush, Wyoming big sagebrush, Bonneville big sagebrush, and/or low sagebrush (potential for juniper and pinyon expansion in some areas) | Moderate. Precipitation and productivity are moderately high. | Moderately low. Climate suitability for nonnative annual grasses is moderately high. |
| Cool and dry (frigid/aridic) | 6-12 inches (152-305 mm) |
Wyoming big sagebrush, black sagebrush, and/or low sagebrush | Low. Effective precipitation limits site productivity. | Moderate. Climate suitability for nonnative annual grasses is moderate. |
| Warm and dry (mesic/aridic, bordering on xeric) |
8-12 inches (203-305 mm) |
Wyoming big sagebrush, black sagebrush, and/or low sagebrush | Low. Low effective precipitation limits site productivity. | Low. Climate suitability for nonnative annual grasses is high. |
Some evidence suggests that mountain big sagebrush sites recover more rapidly after fire than Wyoming big sagebrush sites, and basin big sagebrush sites recover most rapidly of the three [39,349,461,681]. However, on the Northern Yellowstone Winter Range, while basin big sagebrush canopy cover recovered most rapidly, recovery of mountain big sagebrush and Wyoming big sagebrush canopy cover were slow [681].
In general, prefire cover of big sagebrush influences postfire recovery rates [111,230,518,532] because big sagebrush cover is positively correlated with seed abundance in soil seed banks [9,256,370]. Mountain big sagebrush recovered more rapidly after fire in northeastern Utah on sites with >20% mountain big sagebrush prefire cover than sites with <20% prefire cover [230]. One year after a late September prescribed fire in a mountain big sagebrush-grass community in Wyoming, mountain big sagebrush seedling density was higher on an eastern than a western aspect, likely because of greater mountain big sagebrush cover on the eastern aspect before and after fire [518]. On the Deerlodge National Forest, Montana, postfire cover of mountain big sagebrush seedlings was greatest on relatively moist sites that had high prefire mountain big sagebrush cover [111].
As prefire cover of native shrubs and herbs decreases and that of conifers increases, postfire recovery of mountain big sagebrush communities becomes slower [42], and postfire invasion by cheatgrass becomes more likely [43]. In mountain big sagebrush communities in southeastern Oregon, mean mountain big sagebrush cover reached unburned levels more quickly after cut-and-burn treatments in midsuccessional western juniper woodlands with 18% tree canopy cover than in late-successional western juniper woodlands with 26% and 31% tree canopy cover [42]. In a related study, the herbaceous layer in midsuccessional western juniper woodlands was dominated by native plants before and 3 years after treatments, while the herbaceous layer in late-successional western juniper woodlands shifted from dominance by native plants before treatments to dominance by cheatgrass 3 years after treatments. Prefire cover of native shrubs was 3.5 times greater and of native herbs was 2.5 times greater in midsuccessional than late-successional woodlands [43].
Although cheatgrass is problematic in all big sagebrush ecosystems, it is less problematic in mountain big sagebrush than Wyoming big sagebrush ecosystems (see Considerations for nonnative invasive plants). Resistance of Great Basin sagebrush ecosystems to cheatgrass invasion depends on site characteristics [123], particularly soil temperature and moisture regimes and cover of perennial grasses prior to and immediately after fire [416]. Low soil temperatures constrain cheatgrass germination, growth, and reproduction in mountain big sagebrush and mixed mountain shrublands; however, soil temperatures are optimal in relatively warm, dry Wyoming big sagebrush communities. Slope, aspect, and soil characteristics modify soil temperature and moisture and influence resistance of sagebrush communities to cheatgrass establishment and spread at landscape to plant community scales [33,121,122,123,139,416] (figure 10B). For example, 3 years after fire at a high-elevation site (7,900 feet (2,400 m)) in Wyoming, cheatgrass cover was considerably greater on a southwestern aspect (20%) where Wyoming big sagebrush occurred than on eastern aspects (<3%) where mountain big sagebrush occurred [144]. In the Uinta Mountains, Utah, cheatgrass was generally uncommon in mountain big sagebrush communities on plateaus above 8,040 feet (2,450 m), but it was "conspicuous" on south-facing aspects, particularly on steep slopes (>40%) [227].
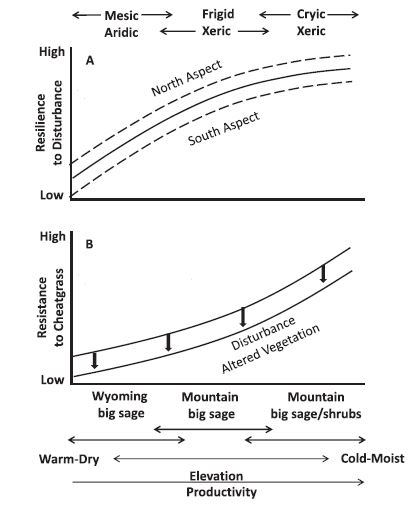 |
| Figure 10—A conceptual model of A) resilience to disturbance and B) resistance to cheatgrass invasion as they relate to soil temperature and moisture regimes, elevation, and productivity gradients in the Great Basin. Predominant sagebrush types that occur along this continuum include Wyoming big sagebrush on warm, dry (mesic/aridic) sites; mountain big sagebrush on cool, moist (frigid/xeric) sites; and mixed mountain shrublands with mountain big sagebrush and sprouting shrubs on cold, moist (cryic/xeric) sites. As environmental gradients move from left to right, resilience, resistance, and biomass (i.e., fuels) increase ([122,416], adapted from [120,121]). |
Although low soil temperatures constrain cheatgrass establishment and spread in mountain big sagebrush communities, cheatgrass may dominate the understory in the first few postfire years [36,246] and impede mountain big sagebrush establishment [235,760] (see Interference and competition). On productive sites, cheatgrass cover may diminish over time [246]. For example, nonnative annual grasses, primarily cheatgrass, dominated plant communities at two sites during the first 3 years following late-summer prescribed fires in mountain big sagebrush communities in northeastern California. Prior to burning, the sites had scattered western juniper and an understory of perennial grasses and forbs with patches of cheatgrass. By postfire year 10, nonnative annual grass cover was surpassed by native perennial grass cover, which peaked 20 years after fire. The authors suggested that early postfire domestic sheep grazing may have reduced the persistence and density of nonnative annual grasses. Recovery of mountain big sagebrush and other shrubs was slow, with only minimal shrub cover observed 10 to 20 years after fire. Mountain big sagebrush canopy cover returned to prefire levels (~30%) within 30 years after fire [246]. Long-term, postfire response of cheatgrass in mountain big sagebrush communities is poorly understood, partly because long-term studies are lacking and partly because cheatgrass cover fluctuates with precipitation, and it is difficult to separate the effects of climate and time-since-fire on cheatgrass cover [416]. For more information about fire effects on cheatgrass, see Considerations for nonnative invasive plants, Miller et al. [416], and the FEIS Species Review on cheatgrass.
Greater cover of native perennial grasses prior to and immediately after fire increases resistance to nonnative annual grass establishment [50,121,122,416,526] and may improve postfire recovery of mountain big sagebrush [139]. In mountain big sagebrush and Wyoming big sagebrush communities in Nevada and Utah, Chambers et al. [123] found that 2 years after treatment, cheatgrass biomass and seed production increased 2- to 3-fold after killing native perennial herbs with herbicide, 2- to 6-fold after prescribed fire, and 10- to 30-fold after herbicide and fire combined [123]. Six years after a large wildfire in late-successional singleleaf pinyon-Utah juniper communities with small pockets of mountain big sagebrush shrublands in the Toiyabe Range of central Nevada, postfire perennial herbaceous cover was positively correlated with postfire mountain big sagebrush cover and was negatively correlated with postfire cheatgrass cover [139]. Implementing a program of grazing rest or deferment after fire may promote perennial herb recovery in mountain big sagebrush communities [657] (see Managing postfire livestock grazing).
Topography may affect mountain big sagebrush recovery by influencing the distribution of mountain big sagebrush seedling recruitment. After a wildfire on a mountain ridge in central Nevada, the pattern of postfire recruitment of mountain big sagebrush suggested that a ridge formed a topographic barrier to seed dispersal. Six years after the fire, most mountain big sagebrush seedlings occurred within 33 feet (10 m) of the burn edge, and seedling frequency decreased with distance from the burn edge (P = 0.018) on the ridge. Where the burn edge fell behind the ridge, few mountain big sagebrush seedlings occurred, and no seedlings occurred within 33 feet of the burn edge [138].
Mountain big sagebrush plants occur on all aspects [398,423], but high temperature variation and low available moisture may limit mountain big sagebrush establishment on southern aspects. For example, in a postfire chronosequence of burned singleleaf pinyon-western juniper/mountain big sagebrush woodlands, mountain big sagebrush was most frequent on northern, eastern, and western aspects [329]. Time-since-fire and incident radiation, a relative index of aspect, explained 66% of the variation in postfire mountain big sagebrush cover (P = 0.001) on 16 large, high-severity burned sites in California, Oregon, and Nevada. Recovery was greatest on steep, cool, northern aspects [767]. In central Oregon, mountain big sagebrush cover was 3% on northern aspects, but 0% on southern aspects, 4 years after a high-severity, late-September prescribed burn in mountain big sagebrush stands in midsuccessional stages of western juniper woodland expansion [163].
Fire characteristics: As of 2017, few studies examined the effects of fire characteristics on mountain big sagebrush postfire recovery. However, because mountain big sagebrush postfire regeneration is by seed, fire characteristics that affect the amount of soil-stored seed consumed by fire (fire severity and timing), the number and distribution of surviving adult plants (fire severity and pattern), and the distance downwind from parent plant seed sources to the burn (fire pattern and size) have a strong influence on the rate of mountain big sagebrush postfire recovery [469].
Fire severity: Wildfires in mountain big sagebrush communities are typically stand-replacement fires that kill or top-kill most vegetation (e.g., [31,398,527,533,681,698,767,768]); however, fires are often patchy, due to variation in fuels, topography, and weather, resulting in a landscape mosaic of unburned patches and patches burned at varying severities [31,415,601]. Thus, postfire mortality of mountain big sagebrush plants and seeds may vary across a burned landscape [65,546]. Fire severity patterns affect mountain big sagebrush recovery because they affect the amount of soil-stored seed and the number and distribution of adult plants that survive, and because they influence the postfire response of the herbaceous layer. Differences in fire severity may, in part, account for variable mountain big sagebrush recovery after fire [745].
Although studies comparing the effects of different fire severities on mountain big sagebrush postfire recovery are lacking, and many studies of fire effects on mountain big sagebrush do not report fire severity, it is likely that high-severity fires slow establishment and recovery of big sagebrush by killing seed-producing plants and consuming seeds in the soil [42,100]. Bunting et al. [100] suggested that mountain big sagebrush density and cover on productive sites in the northern Great Basin may recover to prefire values in 15 to 20 years, but recovery would be longer (30 years) after severe fires. Baker [31] proposed that postfire recovery of mountain big sagebrush may take >75 years in situations where initial postfire recruitment conditions are poor, such as after large, high-severity fires. Mountain big sagebrush density 15 years after wild and prescribed fires in Fremont County, Idaho, averaged 69%, 61%, and 57% of nearby unburned areas on low-severity, moderate-severity, and high-severity burned areas, respectively, suggesting somewhat slower postfire recovery after high-severity fire. However, 15 years after prescribed fire in Clark County, Idaho, mountain big sagebrush density did not show this pattern. Density averaged 29%, 14%, and 21% of that on nearby unburned areas on low-, moderate-, and high-severity burned areas, respectively. Differences in recovery rates between sites may be attributed to higher average annual precipitation at the Fremont County site (16 inches (406 mm)) than at the Clark County site (11 inches (279 mm)) [65].
Because prescribed fires are often less severe than wildfires [66,100,300], mountain big sagebrush recovery may be faster after prescribed fires. However, only one study examined this relationship and it found no difference in postfire recovery between sites burned by wildfire and those burned under prescription at 28 sites in southwestern Montana (P = 0.15), even though there were often more surviving mountain big sagebrush plants in sites burned under prescription [349].
Fire size and pattern: Fire size and pattern vary in mountain big sagebrush communities. Fires in mountain big sagebrush communities range from <25 acres (10 ha) to >2,500 acres (1,000 ha) [169,201,270,272,640,767] and from patchy [31,65,392,455,516,532,746] to uniformly high-severity [31,392,527,698,746,767,768]. Although it is difficult to reconstruct the sizes and shapes of past fires in big sagebrush communities [28,31], fire-size distributions estimated from General Land Office survey records in Colorado [96], Idaho, Nevada, Oregon, and Wyoming [97] suggested that historical fires in big sagebrush communities were mostly small (less than ~1,200 acres (500 ha)) and large fires were infrequent.
Big sagebrush communities are likely to recover slowest after large, uniformly stand-replacement fires and fastest after small, patchy fires because most big sagebrush seeds come from unburned parents or are dispersed onto burns from off-site sources [320,447,485,628,696]. Baker [31] concluded that mountain big sagebrush canopy cover could recover to >85% of unburned values within 25 to 35 years after a "small" fire, but would take >75 years after a "large", uniformly high-severity fire.
If mountain big sagebrush seeds are not present within the burn perimeter, then distance and direction from seed sources at the burn edge are important in determining the rate of establishment of mountain big sagebrush in burned areas [139]. If mountain big sagebrush seeds are present in the burn, the rate of mountain big sagebrush seedling establishment in burned areas may be relatively fast, even in large burns [319,698] (see Postfire seedling establishment). Fifteen years after fire in Idaho, there were fewer mountain big sagebrush plants in burned plots far from seed sources in unburned areas than plants close to unburned areas (r = -0.138, P < 0.05) [65]. Mountain big sagebrush canopy cover diminished along a 3,300-foot (1,000 m) transect from the edge to the interior of a ~360-acre (146 ha), 14-year-old, high-severity burn near Stone, Idaho. Canopy cover of mountain big sagebrush was 30% in the unburned area, 3% at 600 feet (183 m) into the burn, and 0% along the remainder of the transect in the interior of the burn. The author determined that the rate of spread of mountain big sagebrush into the interior of the burn from west to east (downwind) was 42 feet/year (13 m/year), from east to west (upwind) and from north to south was near zero, and from south to north was about 21 feet/year (6 m/year). Based on these measurements, it would take at least 71 years for mountain big sagebrush to establish 3,000 feet (900 m) into the burn, and longer to reach full canopy recovery. However, widely scattered mountain big sagebrush plants within the burn interior, which were probably established from seed in the soil seed bank, may speed the recovery rate [698].
Postfire weather: Mountain big sagebrush stands may recover more quickly after fire if above-average precipitation occurs the fall and winter after burning [460]. Mountain big sagebrush recovery was positively related to postfire precipitation from October to March in 36 low-elevation sites burned between 1971 and 2007 in the Great Basin and Colorado Plateau in Utah and eastern Nevada (R = 0.41, P < 0.0001). This was probably because deep soil-water recharge that persists throughout the growing season enhances first-year seedling survival [460]. Raper et al. [518] reported dense mountain big sagebrush seedlings (2,691 seedlings/ha) in a mountain big sagebrush-grass community in Wyoming, 1 year after a late September prescribed fire that was preceded and followed by years of above-average precipitation.
Postfire herbivory: After fire, heavy browsing may injure mountain big sagebrush and slow its recovery (e.g., [267,398,671,674,681,682]) by concentrating browsing on surviving or reestablishing shrubs [267]. Nineteen years after a wildfire in the Gardiner Basin, canopy cover of mountain big sagebrush, basin big sagebrush, and Wyoming big sagebrush in burned stands was between 1% and 20% of that in adjacent, unburned stands, which were already in decline from historically heavy browsing. Recovery of Wyoming big sagebrush canopy cover was less than that of mountain big sagebrush and basin big sagebrush [398]. Nine to 15 years after prescribed fires in mountain big sagebrush communities in the Gardiner Basin, mountain big sagebrush mean canopy cover on unburned sites was 12 times greater and density 15 times greater than that on burned sites, where ungulate browsing was classified as “high” [267]. Ten years after a 1988 wildfire in a mountain big sagebrush-bluebunch wheatgrass-Idaho fescue community on the Black-tailed Deer Plateau of the Northern Yellowstone Winter Range, mountain big sagebrush cover averaged 20% in areas protected from elk and 10% in unprotected areas (P ≤ 0.01). Mountain big sagebrush density, however, was similar between protected and unprotected sites [682].
However, mountain big sagebrush density may increase following heavy livestock grazing of associated grasses in burned areas. Mountain big sagebrush cover ranged from 12% to 36%, 25 to 42 years after fire in northeastern Utah. The authors hypothesized that livestock grazing may have contributed to increased rates of mountain big sagebrush recovery at some sites, but these data were not analyzed because of a lack of sites without a history of livestock use [230]. At the US Sheep Experiment Station, sagebrush density was greater on a heavily grazed 9-year-old prescribed burn (55 sagebrush plants/100 ft²) than on a "conservatively" grazed 12-year-old prescribed burn (5 sagebrush plants/100 ft²). Lower sagebrush density on the conservatively grazed site was attributed to increased competition for soil water and other resources with dense grasses and forbs [486]. For more information, see Livestock grazing.
 |
| Figure 11—Old fireline separating mountain big sagebrush and bluebunch wheatgrass communities, Lassen County, California. Photo courtesy of the PRBO Conservation Science Shrubsteppe Monitoring Program. |
Fuels: Sagebrush foliage is highly flammable [90,462]. Foliar heat content (maximum amount of energy that can be generated by burning leaves) increases during the growing season, coinciding with decreased live fuel moisture content [517]. Pickett et al. [507] provide information on mass release rates at ignition and maximum flame height for live mountain big sagebrush leaves that may be useful in operational wildland fire spread models. Wright and Prichard [742,746] proposed models for predicting shrub, nonshrub, and total aboveground biomass consumption based on data from spring and fall prescribed fires in mountain big sagebrush, Wyoming big sagebrush, and basin big sagebrush communities in California, Nevada, Oregon, and Montana.
Mountain big sagebrush sites are among the most productive sagebrush sites [227,417,553], and mountain big sagebrush stands often have enough fuels to carry fire [66] (see Fire management considerations for fuels). Biomass, and thus fuel loading and often fuel continuity, generally increase along an environmental gradient from warm, dry Wyoming big sagebrush sites to cool, moist mountain big sagebrush sites and cold, moist mountain shrub sites (figure 10) [416]. Surface fuels in mountain big sagebrush shrublands were historically characterized by a relatively dense herbaceous layer. The predominance of Mollisols in mountain big sagebrush communities shows long-term (hundreds to thousands of years) dominance of grasses on those sites [227] (see Soils).
Late-successional communities produce the most vegetative biomass [398]. In a review, Miller and Eddleman [417] reported total annual herbaceous production in mountain big sagebrush stands ranging from 625 to 2,454 pounds/acre (700 to 2,750 kg/ha), while basin big sagebrush ranged from 774 to 2,098 pounds/acre (868 to 2,352 kg/ha) and Wyoming big sagebrush ranged from 393 to 691 pounds/acre (440 to 775 kg/ha). Goodrich et al. [227,228,230,232] reported total annual herbaceous production in mountain big sagebrush communities ranging from 373 to 2,100 pounds/acre (418 to 2,354 kg/ha). The low end of the range (373 pounds/acre) was from near Dubois, Idaho, where sites were likely near the ecotone with Wyoming big sagebrush [227].Mountain big sagebrush sites have mostly small-diameter fuels. In the Bighole Valley, Montana, shrubs, herbs, litter and duff accounted for 96% of the total surface fuel loadings in mountain big sagebrush/bluebunch wheatgrass stands. Surface fuels were typically <1 inch (2.5 cm) in diameter. Surface fuel loading averaged 0.005 kg/m² for 1-hour fuels, 0.009 kg/m² for 10-hour fuels, 0.004 kg/m² for 100-hour fuels, and 0 kg/m² for 1,000-hour fuels [314]. These correspond to diameter size classes of <0.25 inch (0.62 cm), 0.25 to 1 inch (0.62–2.54 cm), 1 to 3 inches (2.54–7.62 cm), and >3 inches (7.62 cm), respectively [524]. Dead stemwood is common in old mountain big sagebrush plants [90,673,741] (see Botanical description).
When trees establish and spread in mountain big sagebrush communities (see Shrubland succession and Woodland expansion), fuel characteristics change because cover of live mountain big sagebrush and herbaceous plants decreases as tree cover increases, and communities succeed from sagebrush steppe to late-successional conifer woodland (e.g., [35,329,413,426,523,617,688,745,750,751,765]). Fuel loads increase and peak as trees establish in early woodland succession, then decline from early to midsuccession. In late woodland succession, 10-hour fuel loads decrease, and 100-hour fuel loads increase (figure 12). In mountain big sagebrush steppe and during the initial stage of conifer establishment (Phase I), fuel loads are dominated by herbaceous plants and shrubs, the midsuccessional stage (Phase II) by shrubs and litter, and late-successional stage (Phase III) by living trees and downed dead trees and branches [688,717,751]. Dead sagebrush becomes increasingly common throughout succession. In the Sheep Rock Mountains, Utah, the percent of dead mountain big sagebrush plants in western juniper-singleleaf pinyon woodlands increased from none in <10-year-old stands to 16.2% in a 46-year-old stand, and 66.6% in stands >100 years old [35].
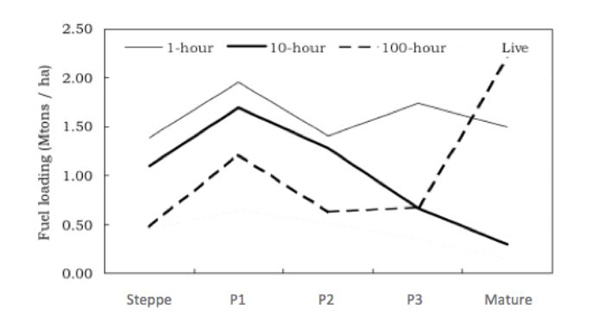 |
| Figure 12—Changes in fuel loading associated with western juniper establishment in mountain big sagebrush steppe (Steppe), early-successional woodland (P1), midsuccessional woodland (P2), late-successional open woodland (P3), and late-successional closed woodland (Mature). Figure from [688], as adapted from Yanish [750]. Image used with permission. |
As mountain big sagebrush steppe succeeds to woodland, tree crowns increase in size, continuity of crown fuels increases, and surface fuel abundance, continuity, and packing ratios decrease. These changes in fuel structure reduce the potential for surface fires burning under moderate weather conditions and increase the potential for crown fires burning under extreme conditions (i.e., high wind, high atmospheric instability, low humidity, and high temperatures) [416,425,601]. On sites where big sagebrush or similar large shrubs dominate, shrubs are more likely than trees to carry fire, especially if trees are widely spaced. As big sagebrush and herbaceous species decline during succession, trees become more important in carrying fire [539]. Simulations suggest that high winds (>15 miles/hour (25 km/hour)) are needed to carry fire through a singleleaf pinyon-California juniper woodland canopy that contains 2.2 tons/acre (5.4 tons/ha) of available canopy fuels, and flame lengths in the woodlands exceed those in big sagebrush-rubber rabbitbrush shrublands [178]. A landscape modeling study suggests that fire size increases as the area of midsuccessional woodland increases on the landscape (r² = 0.785) and decreases as the area of late-successional woodland increases on the landscape (r² = 0.758) [750]
The presence of woodland vegetation can increase fire severity in mountain big sagebrush communities. An analysis of a 46,680-acre (18,890 ha) July wildfire in southwestern Idaho showed that remotely-sensed burn severity in sagebrush steppe measured 1 and 2 years after fire was negatively correlated with the amount of mountain big sagebrush and low sagebrush steppe and early-successional woodlands nearby (approximate range of r-values = -0.5 to -0.7) and positively correlated with the amount of late-successional woodlands nearby (approximate range of r-values = 0.5-0.7, P < 0.05 for all comparisons) (figure 13) [601].
 |
| Figure 13—Photographs of a) mountain big sagebrush steppe before fire, b) midsuccessional woodlands before fire, c) late-successional woodlands before fire, d) mountain big sagebrush steppe 1 year after fire, e) midsuccessional woodlands 1 year after fire, f) late-successional woodlands 1 year after fire, g) mountain big sagebrush steppe 2 years after fire, h) midsuccessional woodlands 2 years after fire, and i) late-successional woodlands 2 years after fire. The photographs are not taken in the exact same location each year, but in the same potential vegetation type and structural stage. Images from Strand et al. [601] and used with permission. |
When nonnative annual grasses establish and spread into big sagebrush communities, the abundance and continuity of fine surface fuels is likely to increase—especially in years following those with abundant precipitation—which can increase fire activity on invaded sites (see Consequences of annual grass invasion). While mountain big sagebrush communities are less susceptible to annual grass invasion than most other sagebrush communities, fire may spread into mountain big sagebrush communities from adjacent, cheatgrass-dominated sites. In the Great Basin, 80% of multi-date fires occurring from 2000 to 2009 that started in cheatgrass grasslands spread into adjacent communities, some of which were likely mountain big sagebrush communities [32]. Figure 14 shows fire spreading from a cheatgrass grassland into a mountain big sagebrush community.
 |
| Figure 14—Fire spreading from a cheatgrass grassland (a site likely formerly dominated by Wyoming big sagebrush and perennial grasses) into a mountain big sagebrush community during the 2011 Constania Fire, Long Valley, California. Photo by Nolan Preece. |
Fire regimes: Fire is one of many natural disturbances that affected mountain big sagebrush communities historically [196,505] (see Successional status). Estimates of historical fire regime parameters in mountain big sagebrush communities can be used as an assessment tool for comparison with current fire regimes and trends and to provide general guidelines for ecological restoration [653].
Presettlement fires in the sagebrush biome were both lightning- and human-caused [27,239,243,569,652,727,730]. Peak fire season occurred between April and October and varied geographically [353]. Wildfires were high-severity with regard to mountain big sagebrush, and therefore stand-replacing [31]. Fire frequency was influenced by sites characteristics, and frequency estimates range from decades to centuries, depending on the applicable scale, methods used, and metrics calculated. Because mountain big sagebrush communities occur over a productivity gradient driven by soil moisture and temperature regimes, fire regimes likely changed across the gradient, with more frequent fire on more productive sites that supported more continuous fine fuels [122,416]. Sites dominated by mountain big sagebrush burned more frequently than sites dominated by Wyoming big sagebrush because the former tends to produce more fine fuels [313,679]. Mountain big sagebrush communities adjacent to fire-prone forest types (e.g., ponderosa pine) may have had more frequent fires [313] than those adjacent to less fire-prone types (e.g., pinyon-juniper) and those far from woodlands and forests. Most fires were likely small (less than ~1,200 acres (500 ha)), and large fires were infrequent [31,96,97]. Large fires were most likely after one or more cool, wet years that allowed fine fuels to accumulate and become more continuous [31,425,428]. Mountain big sagebrush hybridizes with other sagebrush taxa [209,210,220,231,235,382,385,386,542,574,602,695], and sites occupied by parent taxa and their hybrids are distinct from one another [211,379,381,409,684], but no studies to date examined fire history of hybrid communities. Only one study examined postfire recovery rate of a community with what appeared to be a sprouting hybrid of mountain big sagebrush, and this study reported relatively fast postfire recovery [1].
Since European-American settlement, fuel and fire regime characteristics in many big sagebrush communities have shifted outside the range of historical variation. Settlement generally began in the mid-1800s and caused changes in ignition patterns and fuel characteristics, although the timing and magnitude of these changes varied among locations [424]. Since then, fuels and fire regimes in many sagebrush ecosystems have changed due to a combination of interrelated factors, including land alteration for agriculture, rangeland, and energy, urbanization and infrastructure development, proliferation of nonnative invasive plants, woodland expansion, overgrazing by livestock, and climate changes [66,79,141,245,326,419,424,428,439,699].
For more detailed information about fire regimes in mountain big sagebrush communities, including information on fire ignition, season, frequency, severity, pattern, and size during presettlement and contemporary times, see the FEIS synthesis on fire regimes of mountain big sagebrush communities. To find fire regime information for other plant communities in which mountain big sagebrush may occur, enter "mountain big sagebrush" in the FEIS home page under "Find Fire Regimes".
FIRE MANAGEMENT CONSIDERATIONS:A 1987 review included the following objectives of prescribed burning in mountain big sagebrush and other sagebrush-grassland communities: 1) reduce sagebrush cover and density, 2) increase herbaceous plant productivity, 3) increase habitat diversity and edge, 4) reduce conifers, 5) alter herbivore distribution, 6) enhance palatability and nutritional value of vegetation, and 7) prepare for seeding desirable species [100]. In 2017, we contacted several federal resource managers, and many reported that prescribed fire was still commonly used in mountain big sagebrush communities for these purposes (e.g., [252,508,543,549,568,659]), although some have curtailed prescribed burning in recent years due to concern over widespread losses of big sagebrush communities from wildfire and other causes [464,586], including the spread of nonnative invasive species that can occur after fire [104,543,592].
Disagreement about the historical distribution and relative abundance of sagebrush communities and their historical fire regime characteristics occurs throughout the published literature and has led to opposing recommendations about the use of fire in mountain big sagebrush communities [319,398,698]. One view maintains that grassland steppes were not abundant in presettlement landscapes, and, with the exception of lands converted to other uses, the distribution of big sagebrush steppe is similar to presettlement times. While this view acknowledges changes in big sagebrush dominance in response to livestock grazing, altered fire regimes, and other disturbances, it denies the theory that contemporary big sagebrush stands are the result of widespread succession of grasslands to shrublands due to fire exclusion. Advocates of this view (e.g., ([264,305,651,699]) generally oppose management efforts to reduce big sagebrush density. Another view maintains that grassland steppes were abundant during presettlement times and that excessive livestock grazing and the suppression of wildfires have resulted in greater densities of big sagebrush, not only in big sagebrush steppes, but also in grassland steppes. Advocates of this view (e.g., [18,127,145,285,597]) generally consider big sagebrush to be an indicator of grassland degradation, and they support continued efforts to reduce the density of big sagebrush to increase herbaceous plant productivity. The disagreement is likely to continue because reliable historical information is lacking and because of the likelihood that neither model applies across the entire range of big sagebrush [319,398]. Kitchen and McArthur [319] noted that the overall area of big sagebrush-dominated landscapes has decreased since European-American settlement began because of land use conversion, woodland expansion, and more frequent fire associated with nonnative invasive annual grasses (see Other management considerations).
Regardless of the disagreement, managing mountain big sagebrush communities based on historical distribution and fire regime characteristics may not be realistic in landscapes impacted by human development, nonnative plant invasions, and climate change [34,419,653,721]. West [721] stated that resource managers should manage for a mix of desired plant communities in what remains of sagebrush steppe communities, noting that historical sagebrush steppe communities are likely "unrecoverable" because 1) the lack of Native American hunting, gathering, and burning; 2) a warmer and drier climate in contemporary than presettlement times; 3) increased atmospheric carbon dioxide during the past ~100 years; and 4) the presence of nonnative invasive plants [721]. In addition, the large scale of the changes and limited financial and logistical resources make it unlikely that resource managers can return most big sagebrush communities to presettlement conditions [419].
Numerous wildlife species use sagebrush communities, and their needs are important considerations when choosing appropriate management actions in mountain big sagebrush communities (see Considerations for wildlife management). In addition, sagebrush communities differ in their resilience to stress and disturbance and their resistance to nonnative plant invasions along climatic and elevational gradients (see Vegetation and site characteristics), and authors have recommended implementing management actions that will increase resilience and resistance (e.g., [83,86,120,121,122]) under site-specific conditions [230,320]. Managing mountain big sagebrush communities for resilience and resistance primarily involves considerations for managing conifers; promoting native plants and preventing the establishment and spread of nonnative plants; and managing postfire livestock grazing. Tools are available to help managers decide which management options are the most appropriate at a particular site. In general, fire is considered an effective management tool in mountain big sagebrush communities only where sagebrush is abundant, an understory of native perennial grasses and forbs are present, and cheatgrass and other nonnative plants are absent or sparse [169]. Fuel and fire characteristics are important considerations when using prescribed fire in mountain big sagebrush communities. Techniques for using prescribed fire in sagebrush communities, including fireline construction, use of natural fuelbreaks, firing methods, fire weather, and safety are discussed by many authors (e.g., [66,100,647,725,745]) and are not detailed in this review. Refer to other FEIS Species Reviews for discussions of fire management considerations for associated taxa including Wyoming big sagebrush, basin big sagebrush, antelope bitterbrush, bunchgrasses, pinyons, and junipers.
Considerations for wildlife management: General: In a review of the role of fire in sagebrush habitats, Knick et al. [327] stated that the use of prescribed fire to manipulate wildlife habitats is one of the most common yet contentious issues in managing big sagebrush ecosystems. Fire can injure and kill wildlife, although direct impacts are typically minimal, except perhaps during erratic, high-intensity, fast moving wildfires and perhaps in spring during nesting and when young are less mobile [466,624]. The most important influences of fire on wildlife in big sagebrush communities result from impacts on vegetation used for forage and cover [624]. Fires that create habitat mosaics that provide diverse, productive forage near security and thermal cover are often considered beneficial to wildlife, including a broad range of ground, foliage, and aerial feeding birds [21,192,478,502,624], small mammals [393], and wild ungulates [624,749]; however, the effects depend on the ratio of forage to cover over time [131,241,398,624]. Maintaining big sagebrush and native herbaceous species and reducing opportunities for nonnative plant establishment and spread are important in managing big sagebrush communities for birds [258,279,478], small mammals [344,476], and wild ungulates [624,630,753]. Bukowski and Baker [97] suggested that due to the extensive fragmentation of contemporary sagebrush communities and the threat of the establishment and spread of nonnative annual grasses (see Other management considerations), management efforts for wildlife should focus on preserving or rehabilitating large landscapes composed of a mosaic of patches of dense and scattered sagebrush [97].Management objectives for reducing big sagebrush often include increasing the abundance of native perennial grasses and forbs, particularly for sage-grouse (e.g., [142,149,203,259,391,516,747]) and wild ungulates (e.g., [95,144,262,357,358,567]). Although the abundance of native perennial grasses and forbs may increase after fire, abundance of big sagebrush, which also provides food and cover for many wildlife species, particularly in winter, is reduced by fire for many years (e.g., [30,39,166,246,262,753]). A literature review of the effects of prescribed burning in mountain big sagebrush communities concluded that prescribed burning can result in increases in canopy cover, frequency, relative abundance, and production of herbaceous species in the short term (<10 years), but these changes may not persist long-term (>10 years). All studies reported short- and long-term decreases in mountain big sagebrush canopy cover [48]. For this reason, management guidelines for sage-grouse suggest avoiding prescribed burning and suppressing wildfires in critical wintering areas [140,191,556,679].
On the other hand, cover of herbs [7,54,239,418,448,461,720] and mountain big sagebrush [240,413,423,460,540,553,589] decline with increasing conifer cover (see Successional status). Fire can prevent woodland succession in mountain big sagebrush communities [413] and help maintain desirable plants on the landscape [40]. Conifer establishment in mountain big sagebrush communities is generally detrimental to sagebrush obligates, but may be beneficial to facultative wildlife species, when tree density is low enough to support a healthy understory of shrubs and grasses. Low densities of western juniper tend to increase the abundance, diversity, and richness of bird and small mammal populations in shrub-steppe, and small, scattered stands of dense trees may provide thermal cover for wintering ungulates. However, as tree abundance increases, wildlife abundance, species richness, and diversity decline [413]. When using prescribed fire and other methods to reduce conifer establishment and dominance in big sagebrush communities, Holmes and Robinson [272] recommended that residual habitat be maintained at levels suitable to support shrub-dependent wildlife while treated areas recover.
The following discussion provides information on the importance of big sagebrush communities for food and cover, and fire management considerations for different classes and species of wildlife. Many studies of the relationship between wildlife and their sagebrush habitats do not identify sagebrush taxa to species or subspecies, but for many wildlife species, the specific sagebrush taxa may be less important than its height, density, cover, and patchiness [478,731].
Birds: Numerous bird species use mountain big sagebrush for food and cover [699], and their needs are important to consider when managing these communities with fire. More than 90 bird species have a facultative relationship with big sagebrush ecosystems [697], while sage-grouse, sage sparrow, and Brewer's sparrow [269,697] are sagebrush obligates that frequently use mountain big sagebrush communities. For example, the highest densities of Brewer's sparrows occurred in mountain big sagebrush habitats in and around Great Basin National Park, and the total number of both individual birds and bird species appeared to be greatest in study plots with a substantial component of mountain big sagebrush [397].
The importance of sagebrush in the diet of adult sage-grouse is impossible to overstate. Numerous studies have documented its year-round use by sage-grouse (e.g., [47,113,114,322,482,554,576,665,666]). Brewer's sparrows, sage sparrows, and sage thrashers are insectivorous or omnivorous sagebrush obligates that use mountain big sagebrush communities [478]. Brewer's sparrows are shrub-foraging insectivores that glean insects mostly from foliage and bark of big sagebrush and other shrubs or trees in big sagebrush communities. They also eat seeds, particularly in winter, and mainly from the ground [274]. Sage sparrows are ground-foraging omnivores that eat insects, seeds, fruits, and succulent vegetation mainly gleaned from the ground near or under big sagebrush and other shrubs in big sagebrush communities [275]. Sage thrashers are ground-foraging insectivores, with a small portion of their diet consisting of vegetation and fruits found in big sagebrush communities [110].
Sage-grouse [80,141,482], Brewer's sparrows [269,397,525], sage sparrows [525], and sage thrashers [269,525] select mountain big sagebrush communities with relatively high shrub canopy cover. According to a review, these species prefer big sagebrush canopy cover ranging from 20% to 36% [699]. Sage-grouse use sagebrush communities with different heights and canopy cover depending on the season, ranging from 10 to 31 inches (25-80 cm) tall and 12% to 43% canopy cover [141]. Nonobligate bird species using big sagebrush communities do not require canopy cover as dense as that required by obligate species [697].
Fire in big sagebrush communities has varied effects on sage-grouse populations. Some studies report that fires have no short-term effects on sage-grouse population size [223,373,537]. However, sage-grouse populations may decline after fire in big sagebrush communities, particularly when fires occur before or during drought (e.g., [140,582]). Fire pattern affects sage-grouse populations because when sage-grouse use burns, they may rely on unburned patches within burn perimeters [59]. Large, homogenous fires are likely more detrimental to sage-grouse populations than small or patchy fires [192,302,487]. In a simulation of greater sage-grouse population dynamics within mountain big sagebrush communities in southeastern Idaho, Pedersen et al. [487] found that fire was generally detrimental to greater sage-grouse. Populations only increased if fires were very small and infrequent (i.e., burned 1% of the area with 60 years between fires) and left large unburned areas. Their models indicated that large, contiguous fires (10% of the breeding habitat) burning at high frequency (17 years between fires) may lead to the extinction of greater sage-grouse populations. Medium-sized fires (5% of the breeding habitat) burning at medium frequency (25 years between fires) with heavy grazing (consumption of 50% of available forage) may also lead to the extinction of greater sage-grouse populations [487]. Sage-grouse populations are likely to decline when mountain big sagebrush communities succeed to conifer woodlands because sage-grouse avoid conifer communities during breeding, nesting, and brood-rearing [22,37,116,135].
In general, until shrubs recover, shrub-nesting bird populations are reduced by fire in big sagebrush communities, including mountain big sagebrush communities, and ground-nesting bird populations increase or are unaffected (e.g., [21,93,117,183,192,269,272,317,327,468,544,693]). For example, of 12 bird species studied in mountain big sagebrush communities in northwestern Nevada, density of six species (sage sparrow, spotted towhee, gray flycatcher, green-tailed towhee, American robin, and brown-headed towhee) declined up to 20 years after fire, while density of 2 species (horned lark and vesper sparrow) increased; the remaining 4 species showed no clear association between estimated density and time-since-fire [272]. A comparison of disturbed and undisturbed big sagebrush and black sagebrush sites at 5,922 to 9,147 feet (1,805-2,788 m) in Rich County, Utah, revealed that birds that prefer shrubby habitats (Brewer's sparrows, sage thrashers, and green-tailed towhees) tended to have higher density on undisturbed sites, and birds that prefer open, grassy sites (vesper sparrows and horned larks) had higher density on disturbed sites. Shrub cover was lower on disturbed sites, which were disturbed from the 1970s to the 2000s by wildfires or by management to remove shrubs and other vegetation using prescribed fires, herbicides, or mechanical methods [93]. Knick et al. [327] provide a summary table of available literature on the response of breeding birds to fire in sagebrush habitats.
Maintaining sufficient big sagebrush cover in unburned patches within burn perimeters is important to sagebrush obligates [71,192,269,394]. In Oregon, a prescribed fire in October resulted in a mosaic of burned and unburned patches within 1,040 acres (420 ha) of big sagebrush habitat. Brewer's sparrow and sage thrasher abundance was similar on unburned sites and those where the fire removed ≤35% of the shrub canopy, while their abundance on sites where the fire removed >70% of the shrub canopy was 55% and 68% lower, respectively, than on unburned sites (P < 0.05) [269]. Sage sparrow and sage thrasher densities were relatively unchanged after a September prescribed fire in big sagebrush habitat in southeastern Idaho. Brewer's sparrow density declined immediately after the fire but recovered dramatically during postfire year 2. The lack of short-term response of these species was likely because the fire was patchy, burning only about 45% of the study area [502]. In some cases a lack of a short-term response to burning may be attributable to site fidelity of breeding adults [327] or because prefire and postfire habitats are degraded [82].
A 1999 literature review on habitat requirements of birds in sagebrush communities recommended large areas (>100 to several 1,000s of acres, depending on species) of contiguous sagebrush habitat with a mosaic of sagebrush stands of different ages, species, height, and canopy cover to support bird populations. This review stressed the importance of natural disturbances, such as fire, in creating and maintaining this mosaic. The authors recommended that management activities 1) allow herbaceous cover to conceal nests through the first incubation period for birds that nest on the ground or low in shrubs; 2) maintain current season's growth through mid-July; and 3) retain >50% of the annual vegetative growth through the following nesting season [478].
Bird guilds shift as junipers and pinyons establish in mountain big sagebrush communities. As tree canopy cover and density increase, shrub and herb cover and density decrease, and shrub- and ground-nesting bird populations decline. On the other hand, woodland bird species (e.g., ash-throated flycatcher, pinyon jay, American robin, mountain bluebird, juniper titmouse, and western kingbird) colonize mountain big sagebrush steppe once sufficient woodland structure is provided by establishing conifers. In central Oregon, density of ground-nesting birds was highest in 5-year-old burns in grassland steppe, density of shrub-nesting birds was highest in early-successional mountain big sagebrush-Idaho fescue steppe, and density of tree- and cavity-nesting species was highest in old-growth western juniper/Idaho fescue woodlands [525]. In eastern Oregon, Brewer's sparrow, vesper sparrow, and sage thrasher abundance were negatively related to western juniper density and area occupied by western juniper (P < 0.05). Sage thrasher abundance declined sharply at very low western juniper densities. However, the number of bird species was greater in early- and midsuccessional western juniper/mountain big sagebrush woodlands than in mountain big sagebrush communities without trees because tree-nesting species—including chipping sparrow, flycatchers, Cassin's finch, and house finch—increased [467]. Following removal of western juniper trees from mountain big sagebrush steppe communities that were in middle stages of woodland succession in Lake County, Oregon, densities of Brewer's sparrow, green-tailed towhee, and vesper sparrow were higher in treated than untreated areas, while density of gray flycatcher, a species that commonly occurs in the shrubland-woodland ecotone, was lower in treated than untreated areas [271].
Small mammals: Many small mammals are dependent on big sagebrush communities for food and cover. Among these, pygmy rabbit and sagebrush vole have obligate relationships, and are therefore most likely to be affected by fires in mountain big sagebrush communities. In addition, nearly 80 species of small mammals, including black-tailed jackrabbits, squirrels, chipmunks, kangaroo rats, voles, shrews, and mice have facultative relationships. Although small mammals use habitat with a range of big sagebrush canopy cover (0%-68%), a review concluded that the canopy cover of big sagebrush that appears to support the greatest number of small mammal species is between 20% and 50% [697].
Because loss of big sagebrush can decrease both food and cover for pygmy rabbits [222], small, patchy fires are less detrimental to pygmy rabbit persistence than large, homogenous fires [343]. Pygmy rabbits occur in dense stands of big sagebrush and other shrubs on deep alluvial soils, which provide forage, escape cover, and conditions favoring burrow construction [689]. The primary food of pygmy rabbits is big sagebrush, including mountain big sagebrush, which may comprise 99% of their winter diet. They also eat grasses and forbs from mid- to late summer [77,222,237,238,726]. Pygmy rabbits are colonial and tightly clumped in distribution, which makes the species vulnerable to fires that remove shrubs at the colony site. Furthermore, fragmentation of shrub communities may pose a threat by limiting dispersal into favorable habitats [624,689]. Loss of big sagebrush cover after fire can also increase their vulnerability to predation. After an August prescribed fire in a Wyoming big sagebrush-grassland community in Idaho, 6 of 12 radio-collared pygmy rabbits abandoned their home range; 2 of these established new home ranges and the other 4 were predated. Of the 6 that remained on the burned site, only 1 survived through winter, 4 were predated, and 1 was not located [222].
The effects of fire in big sagebrush communities on sagebrush vole populations is relatively unknown. One study found no relationship between time-since-fire and sagebrush vole densities at two sites on the Sheldon National Wildlife Refuge [270,273]. Sagebrush voles occur in big sagebrush-grasslands with 0% to 27% big sagebrush cover and generally require an understory of dense herbaceous vegetation. Grass cover, not big sagebrush cover, appears most limiting for sagebrush vole populations [137,375,377,473]. A conservation assessment indicated that low-severity, patchy, infrequent fires in big sagebrush communities probably have little effect on sagebrush vole populations, except where native grasses are replaced by nonnative plants. Frequent or high-severity surface fires would be detrimental to sagebrush vole populations during the year of the fire by reducing grass cover, which is required for forage and hiding cover. Additionally, complete kill of shrubs would remove the vertical habitat structure sagebrush voles prefer [137]. Vole populations are known to be cyclic, but their population cycles are difficult to predict [21]. A better understanding of both the effects of fire and the factors that trigger eruptions in vole populations would be valuable to mountain big sagebrush management [21].
Patchy fire in big sagebrush and other shrub types may benefit black-tailed jackrabbits by increasing grass and forb production adjacent to shrub cover. Reducing shrub cover over large areas, however, can increase mortality from predation [222,624]. Black-tailed jackrabbits do not burrow, and they require tall shrubs such as big sagebrush to provide adequate cover [470]. Big sagebrush is a primary forage species of black-tailed jackrabbits, and it is used throughout the year [13,198], but especially in fall and winter, while grasses and forbs are mostly eaten in spring and summer [13,198,200,378,685]. In Idaho, black-tailed jackrabbit density was higher in big sagebrush plots burned under prescription than in unburned plots in two of four surveys (P < 0.01), but density was similar on burned and unburned plots in the other two surveys [222,624].
In a study of prescribed fire effects on small mammals, McGee [393] concluded that "ideal" sagebrush management should create a mosaic of various successional stages, so that "no small mammal species, or group of similar species, would be significantly displaced in space and time". Soon after fire, abundance of herbivores decreases, especially on high-severity burns, while abundance of granivores and omnivores may increase because fire consumes vegetation and litter and increases the availability of seeds and invertebrates [262]. Granivores such as deer mice [26,270,276,321] and Great Basin pocket mice [257,359] tend to increase soon after fire in big sagebrush habitats. As the abundance of seeds decreases over time, so does the population of granivores. However, by the third postfire year, new seed-producing vegetation establishes and granivores again increase [262]. The first growing season after a high-severity, August prescribed fire in mountain big sagebrush habitat on the Bridger-Teton National Forest, small mammal density was higher in burned than unburned habitat largely due to increased densities of deer mice and Uinta ground squirrels, both of which eat seeds and tolerate openings. Density of other species, including western jumping mouse and vagrant shrew, declined due to the reduction in total cover. After a patchy, June prescribed fire nearby, small mammal density declined the first growing season but was similar to unburned habitat after three growing seasons [393]. At two mountain big sagebrush sites on the Sheldon National Wildlife Refuge, Nevada, small mammal densities were compared between unburned areas and three, 7- to 19-year-old burns. Belding's ground squirrel densities were approximately 10 times greater on burned than unburned sites regardless of time-since-fire, while Great Basin pocket mouse, sagebrush vole, and least chipmunk densities did not differ between burned and unburned sites. Deer mice densities differed between burned and unburned sites, but the pattern was not consistent. Overall, effects of fire on small mammals in this study were relatively small, perhaps because of rapid recovery of the mountain big sagebrush canopy [270,273]. Welch [697] provides a summary table listing the effects of big sagebrush reduction or removal by fire or mechanical methods on individual species of small mammals in big sagebrush communities, which indicates that most are reduced in abundance by treatments.
The effects of conifer establishment in mountain big sagebrush communities are likely to vary among small mammal populations. Many small mammals, including deer mice, yellow-pine chipmunks, golden-mantled ground squirrels, dusky-footed woodrats, mountain cottontails, and black-tailed jackrabbits, use western juniper foliage and/or female cones for food during part of the year [374,557]; however, increased cover of western juniper also decreases mountain big sagebrush and herb cover, which would likely have a negative impact on many small mammal populations [413]. Several studies in the Intermountain West have shown that small mammal numbers generally increase when western juniper and Utah juniper are either thinned or completely removed, provided that the slash remains (e.g., [189,472,559,732]).
Wild ungulates: Several wild ungulates use big sagebrush ecosystems, including pronghorn, mule deer, elk, and bighorn sheep [697], and Paige [478] considered pronghorn a big sagebrush obligate. Mountain big sagebrush is highly palatable and nutritious browse for these and other wild ungulates [697,712], especially during winter when other forage is less nutritious or unavailable [674,694]. Many studies found that mountain big sagebrush is preferred by mule deer and elk over other sagebrush taxa (e.g., [500,565,566,672,673,675,709,710]). Big sagebrush habitats also provide important cover for ungulates [674,694].
Large-scale burning in sagebrush habitats is detrimental to pronghorn because they rely on sagebrush for food [66,372,465,579,697] and cover, especially during winter [8,223,268,318,323]. Shrubs such as big sagebrush are particularly important during fawning, when does seek areas with greater than average shrub cover and height [24]. However, pronghorn are generally benefitted by fires that create openings in dense sagebrush habitats [262,318,754], because they also require areas of open cover that provide long-range visibility to escape predation [513]. Trainer et al. [634] reported that pronghorn rarely used western juniper woodlands during the winter or spring in eastern Oregon, preferring more open shrub-steppe communities or stands with only scattered trees. In Yellowstone National Park, where mountain big sagebrush dominates pronghorn summer range [558], pronghorn used burned and unburned summer range similarly after the 1988 fires, although burns were preferred over unburned range in some areas [578]. In the Hart Mountain National Antelope Refuge, Oregon, pronghorn did not use an area where sagebrush cover was >50% and shrubs averaged >30 inches (76 cm) tall, until 1 year after a wildfire that converted the range from sagebrush steppe to a grassland-forb community with small stands of sagebrush. Pronghorn continued to use the burn for at least 11 years after the fire [174]. Similarly, pronghorn did not use an area in the Long Valley of California and Nevada where big sagebrush cover averaged 60% and height averaged 23 inches (58 cm). However, within a few years of a series of wildfires that burned about 38,000 acres (15,200 ha), pronghorn moved into the burned areas from adjacent ranges, and they continued to use burned areas until at least 7 years after fire, when cover of big sagebrush and other shrubs averaged 19% and height averaged 17 inches (43 cm) [754]. In the Great Basin, pronghorn prefer sagebrush-steppe habitats where vegetation cover averages ~50%, with about equal proportions grasses, forbs, and shrubs [318,755]. Yoakum [752,755] recommended that prescribed fires in pronghorn habitat be <1,000 acres (400 ha) and leave 5% to 10% shrub cover remaining within fire perimeters.
Mule deer benefit from a mosaic of burned and unburned sagebrush habitats that provide both hiding cover and abundant forage [95,131,144,262,594]; however, fires that result in large expanses of homogeneous vegetation are detrimental [262,361,562,642]. Fire can increase mule deer forage quality and quantity in the short term [58,262,560,562]. Mule deer prefer forbs and grasses when green and succulent and switch to browse when forbs and grasses dry [63,363,474,668], and several researchers reported increases in herb and shrub productivity and nutritional quality after fires on sagebrush rangelands [144,584,641]. Because mule deer use forests or dense shrublands such as mountain big sagebrush habitats for cover, particularly during fawning, and nearby open habitats for foraging [181,224,363,667], most authors recommend a mosaic of burned and unburned habitats within a landscape to benefit mule deer (e.g., [95,131,144,262,567]). Stevens [594] stated that diversity of food and cover over short distances is key to enhancing mule deer populations in big sagebrush areas. Because fire reduces cover, several authors cautioned against using prescribed fire in mule deer habitats where cover is limiting, particularly on winter rangelands [58,199,262,560,562,608], where mule deer browse heavily on big sagebrush [332]. Klebenow [323] noted that mule deer avoided large burned areas on sagebrush-grassland winter range until shrubs recovered. Regardless of habitat, small burns are often considered better for mule deer because portions of large burns may be left entirely unused by mule deer [14,58,63,562,627]. However, small burns may be heavily browsed, which may reduce or eliminate preferred sprouting trees and shrubs [562]. For this reason, Brown [91] suggested burning multiple small areas within a landscape to disperse animals. Alternatively, a single, large fire that creates a mosaic of vegetation may create favorable mule deer habitat while still dispersing animals [91].
Elk are generally associated with a mosaic of open areas for foraging and forested areas for cover [581], and fire tends to maintain this mosaic [92,262,360]. Fire reduces big sagebrush forage, which is considered valuable elk forage in winter and fall [38,331,488]; however, like mule deer, elk eat a variety of plant species and prefer grasses and forbs over browse when available [173]. Elk use of sagebrush communities may increase when palatable grasses and forbs increase and less palatable shrubs decrease after burning [655]. Fire may also enable unpalatable or invasive plants to establish, which can reduce elk forage availability and thus elk use [630,631,633]. Elk rely on a combination of shrubs—including big sagebrush—topography, and areas with little human disturbance to meet cover requirements when preferred forest cover is not available [547]. Big sagebrush provides important cover for elk during calving [301]. Elk most likely benefit from patchy fires that create early-successional habitats that provide forage while leaving interspersed patches of forests and shrublands that provide cover. Elk are not likely to benefit from fires that result in large expanses of homogeneous vegetation [184,262,360,577,615,655,660]. The postfire successional stage when herbaceous plant cover has built up and before tree and shrub canopies close is generally considered "optimum" for elk [581,611]. However, elk use of burned areas varies widely among locations, plant communities, and seasons due to variation in postfire vegetation growth rates, rates of succession, adjacent habitat, and prefire elk density and movements [483,632,654,655]. Having a variety of sizes of burned areas within a landscape may be most beneficial to elk [357,358]. However, small burns may be especially vulnerable to overbrowsing by elk, especially in areas with large elk populations, such as elk winter rangelands [92].
Information is lacking regarding bighorn sheep use of mountain big sagebrush communities after fire. Bighorn sheep prefer habitats generally free of visual obstruction, including mountain grasslands, big sagebrush steppe, and pinyon-juniper woodlands [315,489,585,624,656] that are near escape terrain (e.g., cliffs, rock rims, rock outcroppings, and bluffs with sparse cover of trees or shrubs) [126,248,624,656]. In general, fires that reduce visual obstruction in habitats near escape terrain benefit bighorn sheep by improving visibility and potentially increasing forage [126,490,585,624,663,740]. Bighorn sheep primarily graze green grasses and forbs, but browse woody plants when herbs are unavailable [126], and big sagebrush can be an important part of their winter diet [312,315,372,697]. Fires in mature conifer stands adjacent to escape terrain may maintain or establish bighorn sheep winter range. Early spring fires, particularly on southern and southwestern aspects, may provide more spring forage than would otherwise be available for bighorn sheep [663,740]. Burning immature forests and shrublands adjacent to bighorn sheep winter range could also provide migration corridors between winter and summer ranges [604]. Fire can negatively affect bighorn sheep habitat when range condition is poor and forage species cannot recover, when nonsprouting forage species are eliminated, or when too much area is burned and forage is inadequate until the next growing season [490].
Follow links in table A2 to FEIS Species Reviews for available fire effects information on animal species mentioned in this section.
Managing conifers: Removal of juniper and pinyon trees to increase forage for livestock or improve wildlife habitat has historically been a primary objective of prescribed burning in mountain big sagebrush communities; however, this is not always appropriate because the trees can be an important part of ecosystems in the West [16,45,225,413,539] (see Considerations for wildlife management). Authors stated that if management goals are to restore plant communities to their historical conditions, it is important for managers to collect site-specific information about historical disturbance regimes and landscape dynamics prior to treatment [97,416,539]. Authors generally only advocate for conifer removal in woodlands where trees were historically absent and the density of trees has increased since presettlement times [97,168,205,413,416,539].
Several authors recommended that priority for conifer removal be given to mountain big sagebrush sites in the early stages of woodland succession, before trees become dominant, because communities at this stage are likely to be more resilient, and restoration to mountain big sagebrush dominance is more likely to be successful than on sites in later stages of woodland succession [43,99,101,225,416,425] (see State-and-transition models). Mountain big sagebrush communities in early- to midsuccessional stages of woodland succession that have relatively intact understories of native shrubs and herbs are more likely to be dominated by native than nonnative vegetation after fire [43]. Communities in late-successional stages have depleted understories and shift from a predominance of shrub and herbaceous fuels to tree canopy fuels [416]. This results in a shift from surface fires under moderate weather conditions to crown fires under extreme conditions [751] (see Fuels). Mountain big sagebrush communities are less resilient and less resistant to nonnative annual grass invasions after fires at this stage [43] (see State-and-transitions models).
Prescribed fire, mechanical manipulations, and hand thinning of conifers have been recommended and implemented throughout the range of mountain big sagebrush to slow the establishment and spread of conifers into mountain big sagebrush communities as well as to reduce the risk of high-intensity crown fires [42]. Gentilcore [225] and Miller et al. [414] review the advantages and disadvantages of conifer removal methods. Many of these methods are currently used by federal land managers (as of 2017) to reduce conifers in mountain big sagebrush communities (e.g., [252,508,543,549,659]). Prescribed fire can remove nearly all vegetation and easily covers large areas. However, it can also be unpredictable, hard to control, may burn nontarget species, and often results in greater risk of cheatgrass establishment and spread after treatment. Prescribed fire is most appropriate in areas that are unlikely to convert to cheatgrass, such as cool and cold, relatively moist mountain big sagebrush sites [225,414] (see table 7). Selecting a treatment requires understanding site conditions (e.g., vegetation, soils, climate, and topography), the mechanisms driving woodland expansion, and the effects of the various treatment types at a particular location [16,225,319,539]. See table A1 for links to FEIS reviews available for conifer species of interest.
Considerations for nonnative invasive plants:Consequences of annual grass invasion: Of the nonnative plant species present in mountain big sagebrush ecosystems, annual grasses pose the biggest threat because they alter fuel characteristics in invaded communities and have the potential to increase the frequency, size, spread rate, duration, and severity of wildfires [15,32,325,352,419], such that big sagebrush cannot regenerate [648] and a grass/fire cycle establishes [32,88,156,328,610]. Nonnative invasive annual grasses of concern in big sagebrush ecosystems include cheatgrass, medusahead, and ventenata [149,544]; among these, cheatgrass has been the most harmful [570], and large areas of big sagebrush have been converted to cheatgrass monocultures as a consequence of frequent wildfires [32,156,328,610]. Although mountain big sagebrush communities are less susceptible to annual grass invasion than most other sagebrush types [122,414,415] (table 7), it is a growing concern, especially on relatively warm, dry mountain big sagebrush sites [123,188].
Increased fire activity following cheatgrass invasion has been documented by several researchers (e.g., [62,501,724]) and quantified by others (e.g., [32,325,352,399]). A 1999 land cover map shows that cheatgrass grasslands dominated about 6% of the land cover in the Great Basin and adjacent Mojave Desert, and analyses of burned area data from 1980 to 2009 found that cheatgrass grasslands consistently had more frequent, faster spreading, and longer duration fires and the largest proportional area burned relative to five native cover classes. Fire intervals for cheatgrass-dominated grasslands in 251,000 miles² (650,000 km²) of the Great Basin (calculated as the inverse of the proportion of the total area in the cheatgrass grassland cover class that burned annually) averaged 78 years from 2000 to 2009. Fires were 2 to 25 times more frequent in the cheatgrass grassland cover class than in other land cover classes, including 2.2 times more frequent than in the mountain big sagebrush cover class, where fire intervals averaged 169 years [32]. The likelihood of ignition was greater in cheatgrass grasslands [32], fire spread increased with increasing cheatgrass cover [352], and the number (R² = 0.22) and size (R² = 0.27) of cheatgrass fires were positively correlated with precipitation during the preceding calendar year. Long-duration, multi-day fires between 2000 and 2009 were more likely to start in cheatgrass grasslands than in other cover classes; however, 80% of these fires burned in multiple cover classes. Increased fire size, duration, and spread in sagebrush communities may be caused by cheatgrass invasion in and adjacent to these communities [32].
Preventing nonnative plant invasions: In addition to invasive annual grasses, some deep-rooted, nonnative perennials are becoming increasingly detrimental to sagebrush communities (see Other Management Considerations: Nonnative invasive plants). Preventing invasive plants from establishing and spreading into new areas is the most effective and least costly management approach, and must be considered in fire management plans (Box 1).
Prescribed fire is not recommended in areas where cheatgrass or other nonnative plants are present and likely to spread or become dominant after fire [225,327,366]. Warm, dry sagebrush sites are at greatest risk of cheatgrass establishment and spread, while cool and cold, relatively moist sites have the least risk [36,121,139,419,610,621,739] (table 7). Although mountain big sagebrush communities are among the least susceptible of the big sagebrush communities to annual grass invasion [122,414,415], cheatgrass does dominate the herbaceous layer of some low- and midelevation mountain big sagebrush communities [123,188]. These and other relatively warm, dry mountain big sagebrush sites may be at risk of cheatgrass invasion. In 2005, more than half of the area occupied by mountain big sagebrush and low sagebrush-mountain big sagebrush communities in the Great Basin region of Nevada, Utah, and California was estimated to be at moderate to high risk of displacement by cheatgrass. The sites at highest risk were generally warm, dry sites at low elevations and on south- and west-facing slopes. Sites considered to be at low risk were those at high elevations and on north- and east-facing slopes, which were generally colder and wetter [610]. Cheatgrass usually does not dominate the understory of mountain big sagebrush communities above 4,920 feet (1,500 m) in the northern portion of the sagebrush biome and above 5,250 feet (1,600 m) in the southern portion [139], but it may be present on warm, dry aspects. For example, in mountain big sagebrush communities in the Uinta Mountains, cheatgrass was uncommon on plateaus above 8,040 feet (2,450 m) as of 2005, but it was "conspicuous" in these communities on southern aspects, particularly on steep slopes (>40%) [227]. Ongoing and predicted climate changes are likely to make mountain big sagebrush sites more vulnerable to cheatgrass establishment and spread [479].
Mountain big sagebrush communities with greater than ~20% perennial herb cover have greater resistance to cheatgrass establishment and spread and typically have better recruitment of mountain big sagebrush after fire [121,123,139]. Cheatgrass may initially dominate mountain big sagebrush communities after fire, especially if postfire cover of perennial grasses is low and weather conditions are favorable for cheatgrass establishment and growth (i.e., weather is relatively wet and warm in fall and spring [283,284]) [36,123,246]; however, cheatgrass cover may decline over time, as native perennial vegetation increases [246]. Overgrazing by livestock and infilling of juniper and pinyon trees are likely to decrease cover of native perennial grasses and subsequently decrease resistance to the establishment and spread of nonnative annual grasses [86] (see Vegetation and site characteristics).
In areas where native perennial plant cover is depleted, seeding after fire may help stabilize soils, speed recovery of sagebrush and other shrubs, and prevent establishment and spread of nonnative species [60,148,161,415,536,576,762]. However, seeding sagebrush communities after fire has had low success rates, particularly on warm, dry sites [168,515,546]. For example, analyses of 101 postwildfire seeding sites in Oregon, Idaho, Nevada, and Utah from 1990 to 2003 indicated that postfire seeding was more successful in areas with relatively low annual temperatures (especially cool springs and falls) and relatively high total and spring precipitation. Because mountain big sagebrush communities often occur on sites with these climate characteristics (e.g., at high elevations), postfire seeding may be more successful than in other sagebrush types [17], and may not be needed if native plant species are able to recover naturally after fire [232]. In Utah, natural revegetation (no seeding) was most successful on moist, high-elevation (>6,000 feet (1,820 m)) sites, especially on northern and eastern aspects [362]. Miller et al. [415] provide guidelines for determining when and where to seed sagebrush stands (figure 15). For more information, see Field guides.
On areas where cheatgrass is already abundant, special measures may be necessary to prevent recurrent fires [66] that prevent sagebrush from reestablishing [648]. Grazing management to reduce fuel loads and greenstripping are two methods employed to prevent large, recurrent fires in areas dominated by cheatgrass [494,613]. Greenstrips are fuel breaks placed at strategic locations on the landscape that are seeded with fire-resistant vegetation such as bunchgrasses or other plants that remain green, cure later, or have a caespitose growth form that does not carry fire as easily as cheatgrass. Greenstripping is only recommended in areas where there is a high threat of annual grass invasion and a high threat to high-value sagebrush sites, because greenstrips fragment sagebrush habitat and can increase nonnative plants if the seeding is unsuccessful [478].
Box 1—Preventing the establishment of invasive plants in burned areas can be accomplished through early detection and eradication, careful monitoring and follow-up, and limiting dispersal of invasive plant seed. Specific recommendations from these sources [20,87,234,644] include:
|
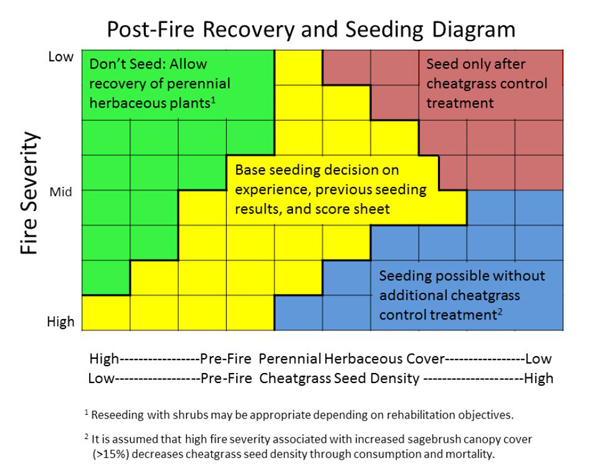 |
| Figure 15—Considerations for postfire seeding in sagebrush communities based on relationships among prefire perennial herbaceous cover, prefire cheatgrass seed density, and wildfire severity. The red blocks represent areas where cheatgrass seed mortality was limited as a result of low to moderate fire severity, and successful postfire seeding requires cheatgrass control. The blue blocks represent areas where postfire cheatgrass seed density is low as a result of high fire severity, and a 1-year window for seeding with minimal competition from cheatgrass typically occurs. The yellow blocks represent areas where vegetation response is less predictable and information on past seeding results, local experience, and use of a score sheet is needed to evaluate resilience to disturbance and resistance to nonnative annual grasses. The green blocks represent areas where prefire perennial herbaceous cover was high and prefire cheatgrass seed density was low or postfire cheatgrass seed density is low as a result of high fire severity, and seeding is not needed for perennial herbaceous plant recovery [415]. |
Managing postfire livestock grazing: Livestock tend to concentrate on revegetating burned areas [130,206,720], including newly burned areas in communities with mountain big sagebrush [129,219]. In southwestern Idaho, cattle selected lightly to moderately burned areas in spring for 5 postfire years after fall prescribed fire in a mesic mountain big sagebrush community; whereas before the fire, cattle had been neutral towards these areas [129]. Grazing in the early growing season immediately following burning may accelerate sagebrush reestablishment at the expense of native perennial herb productivity [486]. This is particularly true in areas with dense sagebrush and low production of grasses before fire [346,749].
Many authors recommend excluding livestock from recent burns in big sagebrush communities for at least the first 1 or 2 years to protect regenerating graminoids and forbs (e.g., [59,66,100,415,486,647,657,725,745]). Additional recommendations include a nongrazing period of 2 to 3 years after fire [59], periodic growing-season rest from grazing for up to 25 years after fire [157], and excluding livestock from burned areas until perennial grasses have recovered and are producing viable seeds equal to prefire or unburned values [114,657]. According to Miller et al. [415,416], the length of time to defer livestock grazing depends on prefire vegetation, site characteristics, fire severity, weather, and whether the burned area was reseeded.
Excluding livestock from recent burns in sagebrush communities for the first 2 years is probably adequate where:
Grazing too soon after fire can promote the spread of nonnative annual grasses by reducing native perennial plants and disturbing biological soil crusts [49,149,168,346,526,646]. Once cheatgrass is established, complete protection from grazing or other disturbances will not usually reduce cheatgrass persistence. A mountain big sagebrush/bluebunch wheatgrass site in southwestern Idaho that was invaded by cheatgrass and subsequently protected from further disturbance continued to be dominated by cheatgrass for decades. Perennial grasses recovered slowly in protected areas, requiring more than 45 years to increase in cover from about 1.4% to 6.7% [435].
In addition to delaying grazing after fire, authors recommend reducing livestock impacts on burned areas by spreading use throughout numerous small, well-distributed burns. Small burns tend to be disproportionately trampled by livestock, and burns more than 0.5 mile (0.8 km) from water have less chance of being "camped on" by livestock [366]. Miller et al. [415] suggested that managers also consider impacts by wildlife, which can be heavy, when determining livestock grazing in burns (see Wildlife browsing).
Decision tools: There is a growing body of literature and tools to help managers manage or rehabilitate mountain big sagebrush and other sagebrush communities. These include the following: Management guidelines:State-and-transition models: Increasingly, managers rely on state-and-transition models to determine recovery potentials and management alternatives for sagebrush communities [395] and to explore how management alternatives may interact with natural disturbances and affect the potential long-term trajectory of the ecosystem [195]. Numerous state-and-transition models have been proposed for mountain big sagebrush communities that describe plant community states and model transitions between states resulting from natural and human-caused disturbances (e.g., [43,121,122,195,245,258,300,503,600,622,717]). Recent models incorporate information on resilience to disturbance and resistance to nonnative annual grasses (e.g., [121]). Changes in disturbance regimes and the establishment and spread of nonnative species can cause plant communities to decline in resilience, with the potential for crossing a threshold to a new state that differs in plant composition, structure, and function. Once a threshold is crossed, a return to the former state is often difficult (and often expensive) to achieve because of altered species composition and site attributes [43,345]. More degraded states are typically less likely to be successfully rehabilitated or restored than healthier states [121]. Wildfire can either help to maintain ecosystem function within a desirable ecological state, or move the ecosystem to a less desirable ecological state, such as one dominated by nonnative annual grasses [622]. Careful assessment of site condition is necessary to determine the relevance of a particular state-and-transition model, the suitability of a site for management, and the most appropriate treatment(s) for the site [121].
Several authors [622,717] developed a state-and-transition model for the Great Basin that describes mountain big sagebrush ecological states and shows how management affects these states (figure 16). The "healthy sagebrush" state (MBS-1a) is dominated by mountain big sagebrush, native and nonnative grasses, and forbs. Wildfire occurring at 60-year intervals maintains this state. Without fire for extended periods, this state may shift to the "pinyon-juniper and mature shrubs" state (MBS-1b). If wildfire occurs within 50 years of transitioning to this state, the MBS-1b state returns to the healthy MBS-1a state. Treatment (i.e., prescribed fire, mechanical treatments, and seeding of desired species) success is 100% in the MBS-1a and MBS-1b states. Without fire for a longer period, the MBS-1b state succeeds to the "closed canopy pinyon-juniper" state (MBS-2). Returning to the healthy MBS-1a state is difficult, with only a 50% treatment success rate, and treatment costs are high. Mechanical treatments are generally used to treat closed-canopy conifer systems. When fires occur in the MBS-2 state, they tend to be high-severity crown fires that kill both conifers and shrubs and result in the "annual grass dominated" state (MBS-3). Fire occurring in the MBS-3 state tends to maintain this state. Treatment is costly and failure is likely in this state. The authors concluded that the probability of treatment success is strongly influenced by the degree of conifer dominance and the relative abundance of native perennial grasses and nonnative annual grasses [622,717].
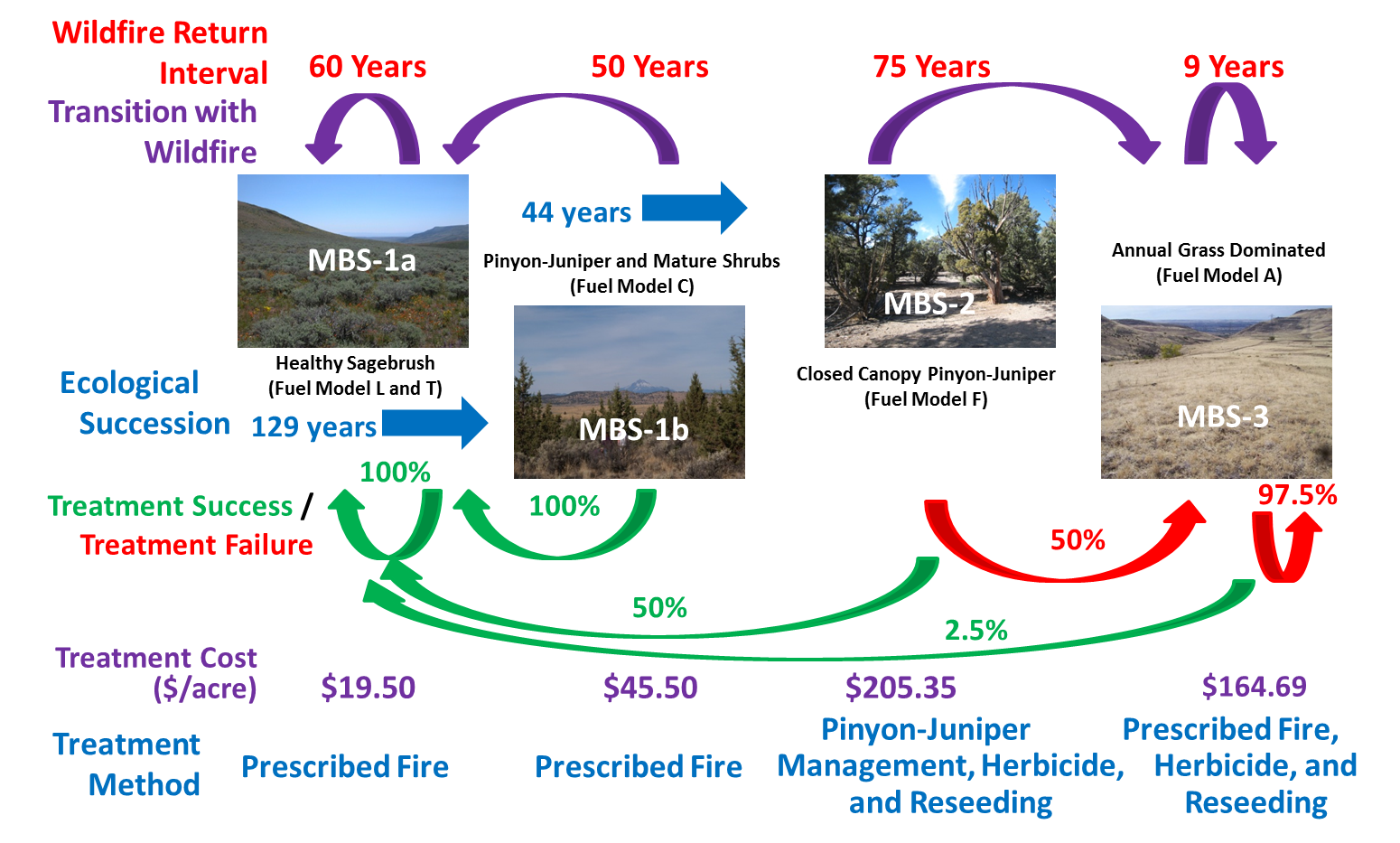 |
| Figure 16—Economic cost of management for alternative states in a mountain big sagebrush plant community [717]. Image used with permission. |
Analyses of state-and-transition models for mountain big sagebrush communities in Utah, Nevada, California, and Oregon [121] and southeastern Oregon [43] suggest that prescribed fire and cutting treatments aimed at reducing conifers are most effective early in woodland succession and for sites with adequate native perennial grass and forb cover. In Utah, Nevada, California, and Oregon, prescribed fire and cut-and-leave treatments in mountain big sagebrush communities in early- to midsuccessional stages of woodland succession aided recovery to a desirable state, and tended to increase native perennial grass and forb cover and shrub recruitment on sites with frigid soil temperature regimes and xeric soil moisture regimes. On sites with mesic soil temperature regimes and xeric soil moisture regimes, such as sites where Wyoming big sagebrush and mountain big sagebrush intermix, treatments tended to increase native perennial grass and forb cover and shrub recruitment, but large increases in nonnative annual grass cover occurred on some sites. On these sites, cut-and-leave treatments were more likely to aid recovery to a desirable state than prescribed fire. Native perennial grass and forb cover of ~20% prior to treatments appeared necessary to prevent substantial increases in cheatgrass and other nonnative annual grass after treatments [121]. In southeastern Oregon, mountain big sagebrush sites in midsuccessional stages of western juniper expansion were dominated by native perennial and annual vegetation up to 9 years after cut-and-prescribed burn treatments, while late-successional woodlands were dominated by cheatgrass. Perennial grass and forb densities greater than 1 and 5 plants/m², respectively, appeared to reduce the chance of a site becoming dominated by nonnative annual grasses after fire [43]. Detailed state-and-transition models applicable to big sagebrush communities in Sage-grouse Management Zones III, IV, V, and VI are provided by Chambers et al. [122]. See figure 18 for a map of these management zones.
While state-and-transition models developed for mountain big sagebrush communities can be informative tools for resource managers, they may "overemphasize (or be misinterpreted as to) the need for treatments while minimizing the relatively stable essence of many sagebrush communities" [723] (see Successional status). Provencher and Blankenship [514] suggested that mountain big sagebrush sites with and without the potential to succeed to conifers in the absence of fire should be differentiated in state-and-transition models.
Considerations for fuels:
Overview: Mountain big sagebrush communities often have enough fine fuel to carry fire [66,84,738] and can be burned under prescription. However, sites where fine fuels are limited do not carry fire well [749] and burn only under severe weather conditions [533]. These include some high-elevation sites, windswept ridges, and sites with shallow soils. A few older studies describe fuel and weather conditions necessary to enable fire spread in big sagebrush ecosystems (e.g., [46,66,85,90,486,647,745]). Very dense sagebrush "monocultures" (often with cover exceeding 60%) may be difficult to burn safely [131].
Field sampling methods to estimate fuel characteristics of mountain big sagebrush were developed in the 1980s [90,172], and more recently, photo series guides were developed to estimate fuels in mountain big sagebrush communities (e.g., [74,477,588,645,743]). Mountain big sagebrush fine fuel (twig) and foliage biomass can be estimated using crown area and height [172]. The strong relationship between these variables (R² = 0.71) makes crown area and height suitable for modeling fuel characteristics of mountain big sagebrush [90]. Procedures for estimating biomass and fuel loads of common shrubs, grasses, and forbs in singleleaf pinyon-Utah juniper/mountain big sagebrush woodlands in Nevada are available [524]. Photo guides that quantify fuels in all strata (e.g., [74,588]) can be used to predict vegetation and fuel response to various treatments, assess target conditions, set management objectives, help choose management activities to meet objectives, and determine treatment effectiveness [74].
Brown [90] quantified fuel properties and modeled fire behavior for mountain big sagebrush and Wyoming big sagebrush in Montana and Idaho and showed how rate of spread and fireline intensity vary with big sagebrush height, percent cover, foliage moisture, and fraction of dead stemwood. See Brown [90] for additional information on metrics and considerations for modeling fire behavior in mountain big sagebrush communities.
Fuel characteristics and subsequent fire behavior change when mountain big sagebrush communities succeed to woodlands [750] (figure 12), and when big sagebrush communities are invaded by nonnative annual grasses (see Considerations for nonnative invasive plants). Fuel models have been developed to predict fire behavior under varying moisture conditions and fuel characteristics during woodland succession [750]. See Fuels for more information.
Grazing management to reduce fuel loads: While impacts of livestock grazing on big sagebrush communities vary (see Livestock grazing), strategic livestock grazing can be used under some conditions to reduce fine fuel biomass and continuity and thereby decrease the risk, size, and severity of wildfires and possibly increase effectiveness of fire suppression activities [167,168]. This is particularly important in areas with cheatgrass-dominated understories [175,190,443,493], and seasonally targeted grazing may be an important tool for breaking the grass/fire cycle [168]. Ungrazed mountain big sagebrush, Wyoming big sagebrush, and low sagebrush plots in southeastern Oregon had ~2 times more perennial bunchgrass cover (P < 0.01) and 1.5 times more total herbaceous cover (P = 0.01) than grazed plots. Standing fine fuel biomass was ~3 times greater in ungrazed than grazed plots (P < 0.01), and total fine fuel biomass (standing biomass plus litter) was ~2 times greater in ungrazed than grazed plots (P < 0.01). Ungrazed plots had more perennial grass cover without fuel gaps (P < 0.01) and smaller fuel gaps (P = 0.03) than grazed plots [167]. Diamond et al. [175] suggest that strategic grazing in nonnative annual grasslands could reduce fuel loads and continuity enough to prevent a flame front from carrying across treated areas even under peak fire conditions, and Davison [170] provides more detailed information on using livestock grazing to reduce fuel loads in cheatgrass-dominated rangelands.
Considerations for fire characteristics: Fire frequency: Miller et al. [419] stated that fires in sagebrush communities should not be more frequent than the amount of time required for sagebrush canopy cover and density to fully recover. Our review and analysis of mountain big sagebrush postfire recovery found that few sites had fully recovered within 25 years after fire, while most burns older than 25 years had fully recovered (figure 6B). However, postfire recovery varied within and among ecoregions (see Plant response to fire), likely because it is influenced by several interacting variables including prefire vegetation and site characteristics, fire characteristics, postfire weather, and postfire herbivory.On the other hand, authors also recommended that fires be frequent enough to prevent tree establishment and succession to conifer woodland (e.g., [320]). Although conifers do not establish in all mountain big sagebrush sites (e.g., [320]), junipers and pinyons can establish, mature, and dominate some sites in as little as 50 years [99,109,320,413,425,759] (see Successional status). Management recommendations include reducing tree dominance in mountain big sagebrush communities in the early to intermediate stages of woodland succession before canopy cover substantially reduces cover of native perennial herbs [139].
Estimates of historical fire frequency in mountain big sagebrush communities can be used as an assessment tool for comparison with current fire regimes and trends and to provide general guidelines for ecological restoration [653]. For more information about historical and contemporary fire frequency in mountain big sagebrush communities, see the FEIS synthesis on fire regimes of mountain big sagebrush communities.
Fire size and pattern: Postfire recovery of mountain big sagebrush communities after prescribed burning is likely to be faster if fires are small or have unburned plants within burn perimeters [139,320] (see Fire characteristics). Mangan and Autenrieth [366] recommended 2.5- to 4.9-acre burns (1-2 ha) to encourage rapid reestablishment of mountain big sagebrush from seeds of unburned plants along burn perimeters.
Mountain big sagebrush shrublands often occur within a mosaic of forests, woodlands, and grasslands [208,369,452,640], and fires historically spread across these community types [32,44,416,601] (see Fuels). For this reason, Kitchen and Weisberg [320] suggested that managing fires at landscape scales and allowing fires to cross vegetation boundaries would best mimic historical fire regime patterns in mountain big sagebrush communities of the eastern Great Basin and Colorado Plateau.
Moffet et al. [432] suggested a shrub management plan with an alternating, mosaic-type repeated-burn strategy when mountain big sagebrush cover exceeds 25%. In their 13,290-acre (5,380 ha) study area at the US Sheep Experiment Station, a postfire recovery model of mountain big sagebrush indicated that a mosaic-type burn could be repeated in the study area at postfire year 17 [432]. Guidelines from Wyoming in 2002 suggested that prescribed fires be used in mountain big sagebrush to create a mixture of successional stages on the landscape, with ~13% of the area in early-successional stages with 0% to 5% mountain big sagebrush canopy cover, ~33% in midsuccessional stages with 5% to 20% mountain big sagebrush canopy cover, and ~53% in late-successional stages with >20% mountain big sagebrush canopy cover. These guidelines also suggested that areas not be retreated until mountain big sagebrush canopy is >20%, ground cover is >70%, and >20 herbaceous species are established [749]. However, Baker [31,96] asserted that most contemporary sagebrush landscapes with mountain big sagebrush, Wyoming big sagebrush, and other sagebrush taxa are highly heterogeneous due to fragmentation from land uses and natural disturbances, and that rest, recovery, and preservation are more appropriate management objectives than further increasing heterogeneity.
Fire season: If the objective is to produce a patchy burn that reduces big sagebrush and increases herbaceous plant production, authors recommended burning when plants are dormant, either in the spring or fall [46,66,100]. For example, Beardall and Sylvester [46] recommended prescribed burning before or just after plants have broken dormancy in spring but not after new growth on grasses reaches 2 inches (5 cm) tall. Burning when soils are wet in spring can result in a patchy burn that allows some big sagebrush plants to survive [46,139,300,596,698]. However, the short time between snowmelt and green-up in mountain big sagebrush communities makes it difficult to burn when plants are dormant [100,366]. This window could last anywhere from a few days to 2 weeks on cool, high-elevation mountain big sagebrush sites [46]. Some burning objectives in big sagebrush are better achieved with prescribed burning in early fall [66,745], although some mountain big sagebrush sites may be too green to burn easily in early fall [703]. Fall fires in big sagebrush communities are generally more severe than spring fires [746].
Managers generally avoid burning mountain big sagebrush sites in summer because conditions are dry and fires tend to be more severe than fires in other seasons [66,100,300]. In southeastern Idaho, mountain big sagebrush, basin big sagebrush, and Wyoming big sagebrush plants burned easily during a winter prescribed fire. Fire weather and equations for a winter ignition index, rate of fire spread, and flame reach were provided by Neuenschwander [462].
Considerations for climate change: Places where conditions are becoming less suitable for big sagebrush may benefit from management actions that promote sagebrush seedling survival, including fire exclusion, managing postfire livestock grazing, reducing conifers, and preventing nonnative plant invasions [196,552]. A review by Finch et al. [202] suggested that managers could facilitate adaptation to climate change impacts by maintaining landscape connectivity to ensure that species can move from unsuitable habitat to colonize more suitable sites. Long-term strategies for dealing with climate change impacts on big sagebrush ecosystems included identifying areas that will maintain sagebrush communities in the future and limiting anthropogenic development in these areas, and accepting and promoting sagebrush expansion into other communities at the leading edge of climate-driven big sagebrush distribution shifts [202]. |
|
| Figure 17—Brewer's sparrow perched on big sagebrush. Photos courtesy of the PRBO Conservation Science Shrubsteppe Monitoring Program. |
FEDERAL LEGAL STATUS:
None
OTHER LEGAL STATUS:
None
IMPORTANCE TO WILDLIFE AND LIVESTOCK:
Cover and food value of mountain big sagebrush for several classes and species of wildlife are discussed in Considerations for wildlife management.
Cattle, domestic sheep, and feral horses and burros use mountain big sagebrush communities extensively [697,757].
Because of its importance as ungulate forage, a method for estimating mountain big sagebrush forage production using plant crown measurements was developed by Dean et al. [172]. Wambolt et al. [670] modeled winter forage production of mountain big sagebrush in southwestern Montana.
See table A2 for links to available FEIS Species Reviews on animals mentioned in this section.
Palatability: The palatability of mountain big sagebrush varies by season, species, subspecies, local populations (accessions), and even individuals [451,701]. Various wildlife species consume all parts of big sagebrush, including the leaves, stems, pollen, seeds, and roots [703]. Mountain big sagebrush is considered highly palatable to most wildlife browsers [541], and domestic sheep may use it extensively, but it has low palatability for cattle [66,179,564,628,701,712]. During the growing season, most herbaceous species are more palatable to livestock than big sagebrush [49,149,186,556,628] (see Livestock grazing).
Sage-grouse, voles, and ungulates browse mountain big sagebrush primarily in winter, when mountain big sagebrush is more palatable than other available vegetation [697]. Mountain big sagebrush sometimes escapes winter browsing by sage-grouse [25,158] and wild ungulates [128,143,579,683]) because of deep snow, while deep snow may increase vole browsing [214,444,695]. Wyoming big sagebrush may be more available than mountain big sagebrush in winter due to shallower snowpack in Wyoming big sagebrush communities [579] (see Climate).
The aromatic smell of big sagebrush is from volatile oils in the leaves (e.g., terpenoids and coumarins) that serve as a chemical-defense mechanism to limit herbivory [748] and protect against rapid temperature changes and water loss (Adams and Billinghurst 1927, cited in [566]). Wildlife species such as pronghorn and sage-grouse that ingest large quantities of sagebrush have developed digestion systems to cope with these compounds [674,695,748]. Sagebrush taxa differ in their concentrations of these compounds. For example, in Gardiner, Montana, crude terpenoid concentrations were lowest in mountain big sagebrush, intermediate in black sagebrush and Wyoming big sagebrush, and greatest in basin big sagebrush [603]. Total monoterpenoids, expressed as a percentage of dry-matter, varied from 0.93% to 1.41% for Wyoming big sagebrush, 0.95% to 1.91% for basin big sagebrush, and 1.02% to 2.95% for mountain big sagebrush plants collected from 20 locations and grown in common gardens [706].
The role of terpenoids in determining preferred browse among sagebrush taxa or local populations of the same taxa is unclear [709]. Some studies concluded that these compounds are responsible for determining browse preferences (e.g., [451,500,566]), while others concluded they are unimportant or less important than other factors in determining preferences (e.g., [134,555,708,726]). For a review of studies on big sagebrush plant chemistry, see Welch [695]. For a review of studies on big sagebrush plant chemistry on the Northern Yellowstone Winter Range, see Wambolt [674]
Palatability of sagebrush species to sage-grouse varies within and among taxa, with mountain big sagebrush being among the most palatable [541]. Among the big sagebrush subspecies, sage-grouse prefer mountain big sagebrush and Wyoming big sagebrush over basin big sagebrush, which is less nutritious and higher in terpenes [25]. In a common garden study in Utah, Welch et al. [714] found sage-grouse preferred mountain big sagebrush over both Wyoming big sagebrush and basin big sagebrush, but the birds shifted to the less preferred subspecies when leaves and buds of mountain big sagebrush became limited.
All sagebrush taxa are potentially valuable forage; however, wild ungulates often show a preference among taxa [675,695]. Several studies found that mountain big sagebrush is preferred by mule deer and elk over other sagebrush taxa (e.g., [500,565,566,672,673,675,709,710]), although preference may vary among mountain big sagebrush populations [726]. Preference for a population of mountain big sagebrush near Springville, Utah, led to the development of a cultivar of mountain big sagebrush ('Hobble Creek') [695,705,710,712]. Welch [695] provides additional information about mule deer and elk preferences among sagebrush taxa.
Nutritional value: Big sagebrush is a nutritious forage for many wildlife species, although nutritional content changes seasonally [695], and may vary among subspecies (e.g., [676,711]) and local populations (e.g., [711]). Big sagebrush nutritional value does not decline as plants age [165,316,505,676].
Mountain big sagebrush protein content and digestibility are typically higher than other available plants in winter [674,694]. Its nutritional value is lower than most associated plants in spring and summer, and it is not browsed as much during those seasons [541,579,694].
A review of studies on the chemistry of mountain big sagebrush, Wyoming big sagebrush, and basin big sagebrush provides data on seasonal variation in crude protein, in vitro digestibility, phosphorus, and calcium, and suggests that big sagebrush in general contains enough protein to meet the maintenance, gestation, growth, and lactation requirements of domestic sheep and cattle year-round; maintenance and gestation of horses; maintenance, gestation, and lactation of mule deer; and maintenance of birds. Winter crude protein content levels of big sagebrush seeds may meet the breeding and growing requirements of birds [694]. In vitro digestibility of big sagebrush ranges from 54% in winter to 59% in summer and fall. All are above the maintenance and gestation requirements of most animals, but do not meet lactation needs. Phosphorus levels of big sagebrush in current year's growth peaks in spring (0.30%) and summer (0.31%), declines in fall (0.25%), and reaches its lowest level in winter (0.20%). At the spring and summer levels, big sagebrush could meet the phosphorus needs for maintenance, gestation, and growth of domestic sheep, cattle, and deer; and maintenance of horses and birds. Calcium levels vary more seasonally than the other nutrients mentioned here, reaching a high of 0.72% in spring and a low of 0.54% in winter. Calcium levels meet the maintenance, gestation, growth, and lactation requirements of domestic sheep, cattle, horses, and deer year-round and the maintenance, gestation, and growth of small mammals, and the maintenance of birds. Only the calcium requirements of breeding birds, growing birds, and lactating small mammals are not met by the winter levels of calcium in big sagebrush [695].
Big sagebrush subspecies may differ in nutritional value [676,695,711]. For example, on the Gallatin National Forest, Montana, midwinter crude protein of mountain big sagebrush (8.34%) was less than that of Wyoming big sagebrush (11.25%), and basin big sagebrush (11.29%) (P < 0.05) [676]. All were above the maintenance requirements of mule deer (7.5%) [690,694]. In a common garden, digestible dry matter was greater in basin big sagebrush (62.1%) than both mountain big sagebrush (53.2%) and Wyoming big sagebrush (51.4%) [711].
Cover value: Mountain big sagebrush provides good cover for many wildlife species [171,179]. Many sagebrush obligate birds, such as sage-grouse [80,141,482], Brewer's sparrow [269,397,525], sage sparrow [525], and sage thrasher [269,525], select mountain big sagebrush communities for cover. Mountain big sagebrush communities provides important cover for many small mammals, including mice, shrews, voles, kangaroo rats, chipmunks, squirrels, and rabbits [697]. Mountain big sagebrush communities also provide important rangelands for wild ungulates, particularly during fawning and calving [24,185,301]. Mountain big sagebrush cover is reduced by burning [420] (see Plant response to fire). Cover and food value of big sagebrush for several classes and species of wildlife are discussed in the section on Fire management: Considerations for wildlife management.
VALUE FOR REHABILITATION OF DISTURBED SITES:
Mountain big sagebrush is easily propagated from seed under greenhouse, nursery, and common garden conditions [193,255,356,529,761] and has been successfully seeded directly into field sites, although results vary [118,147,324,529,564,761]. Mountain big sagebrush has also been planted in field sites using nursery-grown and wilding bareroot and containerized stock with high survival (e.g., [194,290,324,356,404,510,593,701]). Mountain big sagebrush cultivars, such as 'Hobble Creek', were developed for use in rehabilitation projects and on ungulate winter ranges (e.g., ([324,710,712]).
Recommendations for planting and seeding mountain big sagebrush, including rates, timing, and seed bed preparation, are available (e.g., [66,289,337,403,404,436,491,596,701,729]). Planting mountain big sagebrush is recommended in early spring, when moisture conditions are favorable [404]. In general, seeding big sagebrush in the fall and winter, at about the same time that seeds naturally disperse, is recommended [404,701]. Seeding big sagebrush onto snow over disturbed soil has been successful [324,436,596]. Seeding methods that distribute big sagebrush seeds on or near the soil surface result in the best germination [436,701]. Reciprocal transplant experiments indicate that germination response varies between populations of mountain big sagebrush growing in different sites [761] (see Germination). Nonadapted seeds germinate at inappropriate times, and seedlings may fail to emerge or persist [76,405]. Thus, it is important to match seeds and plant collections to sites similar to where they were collected [75,76,407,701]. Mahalovich and McArthur [364] stated that when determining a seed mix, it is more important to match a species to its native environment than to choose a subspecies or cultivar based on wildlife or livestock preference. They recommended that seeds or plants not be moved farther than 300 miles (483 km) from the place of origin to the planting site [364].
Livestock grazing may reduce big sagebrush recruitment in seeded areas. Welch [701] recommended protecting rehabilitation areas from livestock grazing for 2 to 3 years after seeding. Richardson et al. [529] measured the influence of varied seeding rates of grasses, forbs, shrubs, and mountain big sagebrush, and the effects of grazing on the establishment of mountain big sagebrush on mine disturbances. Plots protected from grazing yielded a greater number of mountain big sagebrush plants than unprotected plots.
OTHER USES:
Sagebrush is valued for its antifungal, antimicrobial, analgesic, and anesthetic properties in traditional herbal medicine [573]. Native Americans use big sagebrush leaves and branches for medicinal teas [277,480] and as a "smudge" plant in cleansing rituals [573], and have used big sagebrush plants for many purposes. Big sagebrush seeds were eaten [239]; the bark woven into ropes, mats, baskets, bags, and clothing [277,480]; and the plants used for fuel [573], bedding, and shelter [277,696].
Overview: The land area historically occupied by sagebrush communities has been reduced and altered by the cumulative and interacting effects of land development for agriculture and energy; urbanization and infrastructure development, such as roads and powerlines; proliferation of nonnative plants, particularly annual grasses; woodland expansion; overgrazing by livestock; altered fire regimes; climate changes; water extraction; and reservoir development [66,79,141,245,326,419,424,428,439,699]; however, few studies have compared historical and contemporary mountain big sagebrush distributions quantitatively. Miller et al. [419] estimated that only 55% of the area delineated on Kuchler's maps as potentially dominated by sagebrush was occupied by sagebrush in 2011. In an assessment of the area encompassing the Interior Columbia Basin and portions of the Klamath Basin and Great Basin, Hann et al. [245] estimated that cool shrub (i.e., mountain big sagebrush and mixed mountain shrub) communities occupied about 7.7% of the area from 1850 to 1900 and about 5.1% of the area from 1900 to 1997—a reduction of about 34%. Changes in area occupied by cool shrub communities varied among the Ecological Reporting Units, and ranged from a 1.19% increase in the Central Idaho Mountains to a 97% reduction in the Snake Headwaters (table 8) [245].
| Table 8—Comparison of cool shrub community distribution in the Interior Columbia Basin and portions of the Klamath Basin and Great Basin between two time periods. Cool shrub communities include mesic mountain big sagebrush and mixed mountain shrub communities. Asterisks indicate a significant difference in mean cover between time periods (P < 0.05) [245]. | |||||
| Ecological Reporting Unit | Area occupied (%) 1850–1900 |
Mean area occupied (%) 1900–1997 | Change in area occupied (%) |
||
| Min | Max | Mean | |||
| Blue Mountains | 9.93 | 11.66 | 11.66 | 6.07 | -47.89* |
| Central Idaho Mountains | 6.44 | 9.33 | 6.44 | 6.52 | 1.19 |
| Columbia Plateau | 7.27 | 7.77 | 7.77 | 1.61 | -79.32* |
| Lower Clark Fork | 0 | 0.11 | 0 | 0 | 0 |
| Northern Cascades | 1.05 | 2.59 | 2.59 | 0.28 | -88.99* |
| Northern Glaciated Mountains | 1.46 | 2.07 | 2.07 | 0.84 | -59.25* |
| Northern Great Basin | 4.13 | 5.31 | 5.31 | 2.73 | -48.58* |
| Owyhee Uplands | 19.33 | 21.98 | 21.98 | 20.86 | -5.09 |
| Snake Headwaters | 0.39 | 5.55 | 5.55 | 0.17 | -96.94* |
| Southern Cascades | 1.42 | 4.37 | 1.42 | 0.95 | -33.67* |
| Upper Clark Fork | 1.37 | 3.49 | 1.37 | 1.07 | -21.91* |
| Upper Klamath | 4.44 | 4.90 | 4.90 | 0.71 | -85.49* |
| Upper Snake | 4.91 | 5.95 | 5.95 | 3.14 | -47.28* |
| Total | 7.17 | 7.72 | 7.72 | 5.12 | -33.74* |
Much of the area occupied by mountain big sagebrush is at risk of conversion to conifer woodland or nonnative annual grassland. About a third of the land formerly occupied by sagebrush communities has been converted to other land cover types, including barren, burned, nonnative grassland, nonsagebrush shrublands, and juniper woodlands [419]; and additional areas of big sagebrush are under threat of conversion. Of the 20.4 million acres (8.3 million ha) of sagebrush in the Great Basin region of California, Nevada, and Utah present in 2005, 58% was estimated to be at moderate or high risk of displacement by cheatgrass during the next 30 years, while in 12.0 million acres (4.8 million ha) of sagebrush in the eastern Great Basin, 41% was estimated to be at moderate or high risk of pinyon-juniper expansion during the next 30 years. Mountain big sagebrush communities comprised only 13% of the total sagebrush area in the Great Basin and 9% was at moderate to high risk of displacement by cheatgrass, and mountain big sagebrush comprised 15% of the total sagebrush area in the eastern Great Basin and 60% of this area was at moderate to high risk of pinyon-juniper expansion. Combined, almost 90% of the area occupied by sagebrush communities in the eastern Great Basin, including 65% of the area occupied by mountain big sagebrush communities, was estimated to be at moderate or high risk from at least one of these threats [610]. Though woodland expansion and nonnative annual grass invasion mostly occur in different sagebrush plant community types, Davies et al. [168] concluded that areas prone to woodland expansion and nonnative annual grass invasion appear to be overlapping more commonly than they have in the past. These authors concluded that when sagebrush communities are at risk of both threats the priority should be to address annual grass invasion first [168].
Woodland expansion: Prior to European-American settlement in the mid- to late 1800s, juniper and pinyon-juniper communities were dynamic, expanding and contracting throughout the Holocene (last ~10,000 years) due largely to changing climate and fire patterns [185,413,427,646,745]. Historically, junipers and pinyons spread into nearby big sagebrush communities during periods when the interval between fires was long enough for seedlings to establish and trees to mature [419]. All pinyons and most junipers are killed by fire. Pinyons are intolerant of fire at all life stages, while the probability of western juniper trees being killed by fire decreases for trees >50 years old on high-productivity sites and >90 years old on low-productivity sites [98,109,416,422].
Since European-American settlement, density of junipers and pinyons has increased in many sagebrush and woodland communities [416,426,539], while it has not changed or has declined in others [539]. In areas where conifer expansion into big sagebrush communities has occurred, the peak rate of expansion occurred during a relatively wet and mild period between 1850 and 1916 (e.g., [185,218,303,413,416,425,426,539,618,727,745]). In seven study areas in Idaho, Oregon, Nevada, and Utah, the area occupied by singleleaf pinyon, western juniper, or Utah juniper increased by 125% to 625% between 1860 and 2001. Woodland expansion was not synchronous among all sites but occurred from 1880 to 1920 in Idaho and from 1900 to 1920 in Oregon, Utah, and Nevada [426]. In the central and northern Rocky Mountains, lodgepole pine [53] and Rocky Mountain Douglas-fir [18,19,218,240,242,260] densities have also increased in mountain big sagebrush stands, but these stands are less studied.
The greatest proportion of conifer expansion has occurred on cool to warm, relatively moist sagebrush sites (table 7, figure 18), particularly in mountain big sagebrush communities and low sagebrush communities on moderately deep soils. Conifer expansion has also occurred on relatively cool, moist sites in black sagebrush and Wyoming big sagebrush communities [303,416,419]. The probability of woodlands replacing these sagebrush communities increases on productive sites with nearby conifer seed sources, when fire-free intervals are >50 years [419].
 |
| Figure 18—Distribution of soil temperature and moisture regimes in Sage-grouse Management Zones (MZ I-VII) in the western United States [122]. |
The combined effects of climate variability and related variation in fire frequency were likely the primary drivers of juniper and pinyon range expansion and contraction during prehistoric times, and they continue to be driving forces on contemporary landscapes, along with other interacting effects of overgrazing by livestock and carbon dioxide fertilization [185,413,429,539]
Some authors have attributed juniper expansion since European-American settlement to the effects of a wet, mild climate in the late 20th century coincident with decreased fire frequency (e.g., [108,109,425]), while other authors debate the role of decreased fire frequency in explaining juniper expansion (e.g., [97,185]). Miller et al. [425] suggested that postsettlement western juniper expansion during the late 1880s and early 1900s was driven by mild temperatures and above-average precipitation that promoted conifer establishment and growth and decreased fire frequency that allowed western juniper trees to mature and dominate a site. Decreased fire frequency was attributed to the reduction in Native American burning and the removal of fine fuels by heavy livestock grazing [425]. Burkhardt and Tisdale [108,109] examined several possible causes of and contributing factors to succession of sagebrush-grasslands to western juniper woodlands, and concluded that it was directly related to the combined effects of variability in climate and reduced fire frequency and spread due to fire control and prevention, reduced fine fuel due to livestock grazing, and fragmentation of sagebrush communities due to development. Bukowski and Baker [97] stated that fire regimes in sagebrush communities are primarily controlled by weather or climate, and concluded that estimated fire rotations in mountain big sagebrush communities in four areas of Idaho, Nevada, Oregon, and Wyoming were generally too long for fire to be the only factor preventing conifers from establishing. Eddleman et al. [185] considered the effects of fire suppression to be insufficient to explain western juniper expansion until after World War II. Grove [240] stated that even if fire exclusion was not a primary cause of conifer expansion, it allows it to continue unabated. Juniper and pinyon-juniper woodlands are likely to continue expanding into big sagebrush communities on susceptible landscapes [103,426,610] (see Climate Change).
While conifer expansion is a concern in many big sagebrush communities, tree density and canopy cover have not changed or have declined in many pinyon-juniper communities in the western United States (e.g., [81,368,539]). At Dinosaur National Monument and the surrounding area, a comparison of historical vegetation reconstructed using General Land Office survey records from 1910 with modern records showed a net decline in pinyon-juniper woodlands and montane shrublands and an increase in sagebrush shrublands (a combination of mountain big sagebrush and Wyoming big sagebrush communities). Shorter fire rotations since European-American settlement appeared to be driving the decline. Pinyon-juniper woodlands converted mostly to sagebrush shrublands and grasslands. However, some pinyon-juniper expansion did occur near historical pinyon-juniper-sagebrush ecotones, particularly at 6,600 to 7,900 feet (2,000-2,400 m) and on 10% to 30% slopes [16].
Potential consequences of increasing tree dominance in sagebrush communities are: 1) an increase in woody fuel loads and change in fuel structure that increase the potential for high-intensity crown fires and subsequent establishment and spread of nonnative species; 2) changes in plant community composition and structure, including reduced cover of sagebrush, grasses, and forbs; 3) an increase in aboveground carbon and nutrient pools; and 4) a reduction in water infiltration and an increase in soil erosion [43,161,240,416,503,717]. Changes in plant community composition over time as mountain big sagebrush steppe succeeds to juniper woodland in the absence of fire are illustrated in figure 19 [416]. These changes result in plant communities that are less resilient to fire and other disturbances and less resistant to nonnative annual grass establishment and spread after fire [416].
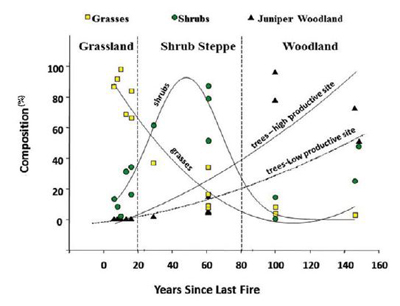 |
| Figure 19—Mountain big sagebrush steppe successional model based on canopy cover data collected from communities with frigid to mesic soil temperature regimes and xeric soil moisture regimes at Lava Beds National Monument [416]. |
Nonnative invasive plants: Mountain big sagebrush communities are not as susceptible to nonnative plant invasions as sagebrush communities at lower elevations and on warmer, drier sites [245,419] (table 7). However, many nonnative plant species have replaced native species in other sagebrush communities [15,32], and mountain big sagebrush communities that occur on sites at the warmer, drier end of their distribution may be similarly susceptible (see Vegetation and site characteristics). Invasive annual grasses—particularly cheatgrass, but also medusahead and ventenata—are considered the most problematic nonnative plants in sagebrush communities because they alter fuel characteristics on invaded sites and have the potential to lengthen the fire season and increase the frequency, size, spread rate, and duration of wildfires [15,32,325,352,419] (see Fire management: Considerations for nonnative invasive plants). As of 2015, deep-rooted, nonnative perennials such as Russian knapweed, squarrose knapweed, Dalmatian toadflax, and Canada thistle were becoming increasingly detrimental to sagebrush communities [15]. Earlier analyses ranked susceptibility of mountain big sagebrush communities as high only for Dalmatian toadflax in 2011 (table 9) [419], and only for cheatgrass and Dalmatian toadflax in 1997 [245]. Figure 20 shows the widespread presence of cheatgrass in parts of the Great Basin and Columbia Plateau.
| Table 9—Susceptibility of upland community types to nonnative plant establishment and spread. Susceptibility is ranked as H=high, M=moderate, L=low, and U=unknown (compiled by [419]). For more information on fire effects on these species, see Miller et al. [416] and the FEIS Species Reviews. | |||||
| Nonnative plant species | Mountain big sagebrush | Basin big sagebrush, Wyoming big sagebrush, and threetip sagebrush | Low sagebrush and black sagebrush | Salt desert shrub | Crested wheatgrass and other bunchgrasses |
| bull thistle | M | M | M | M | H |
| Canada thistle | M | M | M | M | H |
| cheatgrass | M | H | M | M | H |
| common crupina | M | L | L | L | M |
| Dalmatian toadflax | H | M | M | L | H |
| diffuse knapweed | M | M | M | L | H |
| dyer's woad | L | H | H | L | H |
| halogeton | M | M | M | H | M |
| heart-podded hoary cress | M | M | M | M | M |
| leafy spurge | L | M | M | M | M |
| meadow hawkweed | L | L | L | L | L |
| Mediterranean sage | M | H | U | L | H |
| medusahead | M | M | L | M | M |
| musk thistle | M | U | U | M | M |
| orange hawkweed | M | L | L | L | L |
| oxeye daisy | U | U | U | L | M |
| perennial pepperweed | L | L | L | L | L |
| poison hemlock | L | L | L | L | L |
| purple loosestrife | M | L | L | L | L |
| rush skeletonweed | M | M | U | L | M |
| Russian knapweed | M | M | U | M | M |
| Russian-thistle | M | M | L | M | M |
| Scotch cottonthistle | L | M | U | L | M |
| sowthistles | M | M | M | M | M |
| spotted knapweed | M | M | U | L | H |
| squarrose knapweed | M | M | M | M | M |
| sulfur cinquefoil | M | U | U | L | H |
| tansy ragwort | U | U | U | U | U |
| yellow starthistle | M | M | M | L | H |
| yellow toadflax | M | M | U | L | M |
 |
| Figure 20—Cheatgrass presence in parts of the Great Basin and Columbia Plateau as of 2015. Image courtesy of Bethany Bradley. |
Herbivory: Mountain big sagebrush has low tolerance to browsing [217,384,628], because all of its growing points extend above the crown where they are vulnerable to browsing animals [61,699]. Heavy browsing can reduce density, size, and seed production [61,215,346,538,661,662,699] and even kill plants [628], especially where big sagebrush densities are low (<1.5 plants/100 ft²) [215]. The percent of dead crown in live mountain big sagebrush, Wyoming big sagebrush, and basin big sagebrush plants increases in proportion to the overall amount of browsing received. For example, heavy elk and mule deer browsing killed 35% of mountain big sagebrush plants at two study sites near Gardiner, Montana, between 1982 and 1992, and the percent of dead crown in live plants for mountain big sagebrush, Wyoming big sagebrush, and basin big sagebrush, was 59%, 45%, and 30%, respectively [673].
Wildlife browsing:
Small mammals: In winter, long-tailed voles, mountain voles, and meadow voles eat the cambium layer of mountain big sagebrush plants, and in so doing, strip the bark off stems and branches buried in the snow [21,214,695]. When vole populations are large, their browsing can kill or cause extensive damage to big sagebrush plants [214,444,695], even over large areas, especially where herbaceous cover is seasonally high and snow cover is deep [214,444]. Evers [196] included vole damage in the modeled successional rate for cool, moist mountain big sagebrush communities where deep and persistent snow is frequent, and for successional stages with a mix of sagebrush and relatively high grass abundance. An assumed 4- to 5-year population outbreak cycle meant that during 100 years, 23% of the years would have vole populations large enough to thin mountain big sagebrush stands [196].
Black-tailed jackrabbits eat most plant species in sagebrush communities, and big sagebrush is a primary forage [13]. Because black-tailed jackrabbits eat the seedlings of most of these species, Barbour et al. [34] hypothesized that when black-tailed jackrabbit population density is high, seedling recruitment of big sagebrush and other plant species could be eliminated if adequate escape cover is available (e.g., [378]).
Wild ungulates: Heavy wild ungulate browsing decreases mountain big sagebrush canopy cover [673,699]. For example, declines in mountain big sagebrush canopy cover associated with heavy, long-term, mule deer and elk browsing on the Northern Yellowstone Winter Range are described by many researchers (e.g., [267,398,527,674,681,683]) (see Postfire herbivory). In addition to directly eliminating sagebrush, fire concentrates ungulate browsing on burned areas [674] (see Considerations for wildlife management).
Insects: Insects such as aroga moths (e.g., [221,697]) and grasshoppers [235] can damage and kill mountain big sagebrush plants, but they do not seem to pose a threat to mountain big sagebrush populations [697]. Outbreaks of aroga moth may cause nearly 100% mortality of mountain big sagebrush plants in a stand [414]. However, aroga moth populations are heavily parasitized during peak periods and usually decline abruptly after 1 or 2 years at peak level. The sagebrush stands then gradually recover [697]. Large aroga moth outbreaks occurred in southeastern Oregon and southwestern Idaho in the 1960s; most of the damage occurred in old stands [221]. In 2013, Evers et al. [195] stated that very little information was available on aroga moth outbreak frequency, size, and severity. Their models of mountain big sagebrush postfire successional rates suggested that aroga moth outbreaks may have played a much greater role than fire in mountain big sagebrush dynamics prior to 1850 by either affecting larger areas or by occurring more frequently [195].
Livestock grazing: Impacts of livestock grazing within big sagebrush stands can be positive, negative, or neutral to big sagebrush communities, depending on the type, timing, severity, duration, and vegetation composition [49,66,149,192,628,701,703]. Reviews of the effects of livestock grazing on big sagebrush communities are available (e.g., [49,66,628,701]).
Overgrazing by domestic livestock during the late 1800s and early 1900s coupled with severe drought triggered a rapid change in sagebrush communities, resulting in reduced cover and density of native herbaceous plants. The reduction in native herbaceous plants allowed establishment of nonnative annual grasses, particularly on warm, dry sites, and conifers, particularly on cool, relatively moist sites. However, overgrazing alone is not sufficient to explain vegetation changes throughout the sagebrush region over the past 150 years [424]. Added to the threat of livestock grazing itself was a policy of aggressive sagebrush reduction adopted by land management agencies in the 1950s and 1960s. Prescribed fire, herbicides, and mechanical methods were used to convert sagebrush types to grasslands for seasonal grazing by livestock [113,376,439]. These efforts included seeding with nonnative grasses, primarily crested wheatgrass, which affected approximately 6.4 million acres (2.6 million ha) of sagebrush lands by the 1970s [326]. As of 2017, federal resource managers in some regions continued to use prescribed fire and other methods to reduce big sagebrush cover and increase herbaceous plant production for livestock and wildlife (e.g., [252,508,568,659]), although prescribed burning on some federal lands had been curtailed due to concern over additional widespread losses of big sagebrush communities from wildfire, nonnative plant proliferation, and other causes [104,464,586,592]. Past management has had legacy effects on the composition and diversity of sagebrush communities, and many sites have not recovered [534]. Livestock grazing and associated habitat alterations have had the most widespread impact on western sagebrush ecosystems of any land use [326].
Livestock grazing often exacerbates impacts from invasive annual grasses such as cheatgrass; however, cheatgrass abundance may also be reduced by uniformly heavy spring grazing, because it is highly palatable to cattle in spring, and appropriately timed grazing reduces seed production [286,756]. In some cases intensive grazing can be used to reduce the abundance and continuity of fine fuels, thus impeding the annual grass/fire cycle [6] (see Fire management: Considerations for fuels). Heavy spring grazing may also increase cheatgrass abundance by weakening native cool-season perennial grasses [756]. For more information, see the FEIS Species Review of cheatgrass.
Biological soil crusts are an important component of the nutrient cycle in sagebrush ecosystems because they include nitrogen-fixing microbiota, sequester many nutrients—including phosphorus and potassium—and increase soil carbon storage. Livestock grazing disturbs biological soil crusts, and recovery rates of biological soil crusts are very slow [56]. As a result, modern nutrient cycling in sagebrush systems is probably substantially different from that historically, with greater leaching of nutrients from current systems [646]. Where crust communities are well established in sagebrush ecosystems, they help prevent the establishment and spread of nonnative annual grasses [89,122,526,646], and, consequently, also reduce the likelihood and slow the spread of wildfire [57].
When big sagebrush communities are heavily grazed by livestock, herbaceous plant abundance tends to decrease and big sagebrush density tends to increase because most of the herbaceous species are more palatable to livestock than big sagebrush, especially during the growing season [49,149,186,556,628]. However, trampling by livestock may reduce mountain big sagebrush densities on grazed sites [529]. In 18 plots in mountain big sagebrush communities in the Bodie Hills on the California-Nevada border, the highest shrub cover from 1979 to 2011 was associated with years having low precipitation and high livestock stocking densities, while low shrub cover was associated with years having high precipitation and low stocking densities [441].
The introduction of livestock grazing during the late 1800s may also have contributed to woodland expansion in mountain big sagebrush communities. However, evidence for a direct relationship between livestock grazing and woodland expansion is difficult to substantiate because the role of fire must also be accounted for, and the size and complexity of fires prior to livestock introduction are poorly understood. Patterns of woodland expansion in sagebrush steppe have not been related to grazed/ungrazed fenceline contrasts, distance to water, or ecological conditions that are often observed with livestock grazing [419].
Climate change: Climate is a primary control on the occurrence of wildfire in sagebrush ecosystems on annual to decadal time scales (e.g. [422,425,440]), as well as on centennial to millennial time scales (e.g., [282,431,458,571,572,629,718,728]).
Climate change models for the sagebrush biome predict increasing temperatures, increasing atmospheric carbon dioxide, more frequent severe weather (droughts and storms), decreasing snow pack, and variable changes in precipitation [119,419]. These predicted climate changes are likely to favor the establishment and spread of cheatgrass [367,402,419] and thus contribute to the grass/fire cycle, which includes more frequent fire. In addition to the potential for increased fire frequency, a warming climate has the potential to increase the frequency of insect outbreaks and decrease the frequency of snow mold, which could alter vegetation composition in sagebrush communities [119,202,700].
A review of paleobotanical studies showed that big sagebrush communities have been resilient to historical climate changes in many locations throughout the West [699]. This suggests that these communities may be resilient to future climate changes, in the absence of other contemporary changes not present historically (e.g., nonnative plant invasions). However, projected impacts of climate change on sagebrush are varied. Many projections predict widespread shifts in vegetation by the end of the century, with some locations becoming less suitable for sagebrush [78,154,419,454,551,552]. In addition, sagebrush ranges are expected to become increasingly fragmented and threatened by nonnative grasses (e.g., [154,212,454,479,551,561]).
While few studies have examined the effects of projected climate changes on mountain big sagebrush communities, some suggest that they will decrease in range. Mountain big sagebrush is predicted to lose as much as 68% of its currently suitable range by the end of the twenty-first century under modeled climates that included warmer winter temperatures. Substantial range losses are projected under all modeled future climate conditions, largely because in many locations mountain big sagebrush does not have many high-elevation sites to retreat into [621]. Climate change-fire models suggest that big sagebrush distribution is likely to decrease by 13%, with substantial decreases in southern parts of the species' current range and some increases in the northern parts and at high elevations [551]. Taylor [621] suggested that many of the projected losses may be due to the loss of mountain big sagebrush. Evers [196] modeled three potential future climate scenarios in southeastern Oregon: the first with warmer and drier conditions year-round, the second with warmer and wetter conditions in winter, and the third with warmer and wetter conditions in summer. All scenarios predicted a reduction or near elimination of the area covered by cool, moist mountain big sagebrush communities [196]. Simulations of climate change in montane mountain big sagebrush steppe, Wyoming big sagebrush-basin big sagebrush steppe, and Wyoming big sagebrush-basin big sagebrush shrublands throughout the Intermountain West predicted the greatest increases in spring soil moisture and the greatest decreases in summer soil moisture in montane mountain big sagebrush sites. The authors stated that warmer temperatures and wetter winter and spring conditions could result in montane mountain big sagebrush steppe sites becoming increasingly suitable to cheatgrass establishment and spread [479].
Sagebrush seedling establishment may be particularly affected by predicted climate changes [75,367,419]. Experimental warming studies in subalpine meadows at high elevations (9,580 feet (2,920 m)) in Gunnison County, Colorado, showed that mountain big sagebrush seedling establishment and plant growth rates may increase and populations may expand under warming future climates [253,336,354,496,563]. However, mountain big sagebrush plants may not be as able to adapt to increased winter temperature and earlier snowmelt at lower elevations, where these climate conditions are likely to lead to increased summer drought stress [338].
Mountain big sagebrush flowering phenology may be able to adapt to climate warming. A model using mountain big sagebrush, basin big sagebrush, and Wyoming big sagebrush plants grown in common gardens indicated that flowering phenology of all three major subspecies is plastic and can adapt to potential future climate changes expected from the middle to the end of the twenty-first century [530].
Several authors stated that polyploidy and inter- and intraspecific hybridization in big sagebrush is likely to help mountain big sagebrush adapt to potential future climate changes [75,381,387]. Hybridization is also a factor that could affect bioclimatic modeling results where subspecies overlap along ecotones [599], but hybridization has not been included in climate change models. Sagebrush densities may fluctuate during climate changes as older plants die and genotypes adapted to milder winter conditions migrate in [196].| Common name | Scientific name |
| Forbs | |
| agoseris | Agoseris spp. |
| avens | Geum spp. |
| balsamroot | Balsamorhiza spp. |
| buckwheat | Eriogonum spp. |
| bull thistle* | Cirsium vulgare |
| Canada thistle* | Cirsium arvense |
| cinquefoil | Potentilla spp. |
| common crupina* | Crupina vulgaris |
| Dalmatian toadflax* | Linaria dalmatica |
| desertparsley | Lomatium spp. |
| diffuse knapweed* | Centaurea diffusa |
| dyer's woad* | Isatis tinctoria |
| fleabane | Erigeron spp. |
| halogeton | Halogeton glomeratus |
| hawksbeard | Crepis spp. |
| heart-podded hoary cress* | Cardaria draba |
| Indian paintbrush | Castilleja spp. |
| leafy spurge* | Euphorbia esula |
| lupine | Lupinus spp. |
| meadow hawkweed* | Hieracium caespitosum |
| Mediterranean sage* | Salvia aethiopis |
| milkvetch | Astragalus spp. |
| musk thistle* | Carduus nutans |
| orange hawkweed* | Hieracium aurantiacum |
| oxeye daisy* | Leucanthemum vulgare |
| perennial pepperweed* | Lepidium latifolium |
| phlox | Phlox spp. |
| poison hemlock* | Conium maculatum |
| purple loosestrife* | Lythrum salicaria |
| rush skeletonweed* | Chondrilla juncea |
| Russian knapweed* | Acroptilon repens |
| Russian-thistle* | Salsola kali |
| Scotch cottonthistle* | Onopordum acanthium |
| spotted knapweed* | Centaurea stoebe subsp. micranthos |
| squarrose knapweed* | Centaurea virgata |
| sulfur cinquefoil* | Potentilla recta |
| tansy ragwort* | Senecio jacobaea |
| yarrow | Achillea spp. |
| yellow starthistle* | Centaurea solstitialis |
| yellow toadflax* | Linaria vulgaris |
| Graminoids | |
| bearded wheatgrass | Elymus caninus |
| bluebunch wheatgrass | Pseudoroegneria spicata |
| blue grama | Bouteloua gracilis |
| bottlebrush squirreltail | Elymus elymoides |
| bulbous bluegrass* | Poa bulbosa |
| California brome | Bromus carinatus |
| cheatgrass* | Bromus tectorum |
| crested wheatgrass* | Agropyron cristatum |
| Columbia needlegrass | Achnatherum nelsonii |
| Fendler bluegrass | Poa fendleriana |
| green fescue | Festuca viridula |
| Idaho fescue | Festuca idahoensis |
| intermediate needle-and-thread grass | Hesperostipa comata subsp. intermedia |
| medusahead* | Taeniatherum caput-medusae |
| melicgrass | Melica spp. |
| Parry's oatgrass | Danthonia parryi |
| prairie Junegrass | Koeleria macrantha |
| rough fescue, northern | Festuca altaica |
| saline wildrye | Leymus salinus |
| Sandberg bluegrass | Poa secunda |
| sedge | Carex spp. |
| slender wheatgrass | Elymus trachycaulus |
| Thurber fescue | Festuca thurberi |
| Thurber needlegrass | Achnatherum thurberianum |
| tufted hairgrass | Deschampsia cespitosa |
| western needlegrass | Achnatherum occidentale |
| western wheatgrass | Pascopyrum smithii |
| Shrubs | |
| antelope bitterbrush | Purshia tridentata |
| basin big sagebrush | Artemisia tridentata subsp. tridentata |
| big sagebrush | Artemisia tridentata |
| bitterbrush | Purshia spp. |
| black greasewood | Sarcobatus vermiculatus |
| black sagebrush | Artemisia nova |
| Bonneville big sagebrush | Artemisia tridentata subsp. × bonnevillensis |
| broom snakeweed | Gutierrezia sarothrae |
| low sagebrush | Artemisia arbuscula |
| Mojave big sagebrush | Artemisia tridentata subsp. parishii |
| mountain silver sagebrush | Artemisia cana subsp. viscidua |
| mountain snowberry | Symphoricarpos oreophilus |
| plains silver sagebrush | Artemisia cana subsp. cana |
| rabbitbrush | Chrysothamnus spp., Ericameria spp. |
| roundleaf snowberry | Symphoricarpos rotundifolius |
| rubber rabbitbrush | Ericameria nauseosa |
| sagebrush | Artemisia spp. |
| Saskatoon serviceberry | Amelanchier alnifolia |
| serviceberry | Amelanchier spp. |
| shrubby cinquefoil | Dasiphora fruticosa subsp. floribunda |
| silver sagebrush | Artemisia cana |
| snowberry | Symphoricarpos spp. |
| snowbrush ceanothus | Ceanothus velutinus |
| snowfield big sagebrush | Artemisia tridentata subsp. spiciformis |
| stiff sagebrush | Artemisia rigida |
| threetip sagebrush | Artemisia tripartita |
| true mountain-mahogany | Cercocarpus montanus |
| Utah serviceberry | Amelanchier utahensis |
| wax currant | Ribes cereum |
| wild crab apple | Peraphyllum ramosissimum |
| winterfat | Krascheninnikovia lanata |
| Wood's rose | Rosa woodsii |
| Wyoming big sagebrush | Artemisia tridentata subsp. wyomingensis |
| xeric big sagebrush | Artemisia tridentata subsp. xericensis |
| yellow rabbitbrush | Chrysothamnus viscidiflorus |
| Trees | |
| blue spruce | Picea pungens |
| California juniper | Juniperus californica |
| chokecherry | Prunus virginiana |
| Colorado pinyon | Pinus edulis |
| curlleaf mountain-mahogany | Cercocarpus ledifolius |
| Douglas-fir | Pseudotsuga menziesii var. glauca Pseudotsuga menziesii var. menziesii |
| Engelmann spruce | Picea engelmannii |
| Gambel oak | Quercus gambelii |
| Jeffrey pine | Pinus jeffreyi |
| juniper | Juniperus spp. |
| limber pine | Pinus flexilis |
| lodgepole pine | Pinus contorta var. latifolia Pinus contorta var. murrayana |
| pinyon | Pinus spp. |
| ponderosa pine | Pinus ponderosa var. ponderosa Pinus ponderosa var. scopulorum |
| quaking aspen | Populus tremuloides |
| Rocky Mountain juniper | Juniperus scopulorum |
| singleleaf pinyon | Pinus monophylla |
| subalpine fir | Abies lasiocarpa |
| Utah juniper | Juniperus osteosperma |
| water birch | Betula occidentalis |
| western juniper | Juniperus occidentalis |
| whitebark pine | Pinus albicaulis |
| white fir | Abies concolor |
For further information on fire ecology of these species, follow the highlighted links to FEIS Species Reviews.
| Birds | |
| greater sage-grouse | Centrocercus urophasianus |
| Gunnison sage-grouse | Centrocercus minimus |
| pinyon jay | Gymnorhinus cyanocephalus |
| Mammals | |
| bighorn sheep | Ovis canadensis |
| black-tailed jackrabbit | Cynomys ludovicianus |
| deer mouse | Peromyscus maniculatus |
| elk | Cervus elaphus |
| Great Basin pocket mouse | Perognathus parvus |
| mule deer | Odocoileus hemionus |
| pronghorn | Antilocapra americana |
| pygmy rabbit | Brachylagus idahoensis |
1. Abendroth, Diane. 2018. Personal communication [Email to Robin Innes]. 23 January. Unpublished reports and information regarding mountain big sagebrush postfire recovery. Moose, WY: U.S. Department of the Interior, National Park Service, Teton Interagency Fire Center. Documents on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92578]
2. Abendroth, Diane. 2018a. Personal communication [Email to Robin Innes]. 5 February. Unpublished 2013 report and information regarding the Blacktail Butte Area bitterbrush burns - 15 years post. Moose, WY: U.S. Department of the Interior, National Park Service, Teton Interagency Fire Center. Documents on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92592]
3. Abendroth, Diane. 2018b. Personal communication [Email to Robin Innes]. 5 February. Unpublished 2014 report and information regarding the Fremont Ridge prescribed burn - year 10. Moose, WY: U.S. Department of the Interior, National Park Service, Teton Interagency Fire Center. Documents on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92590]
4. Abendroth, Diane. 2018c. Personal communication [Email to Robin Innes]. 5 February. Unpublished report and information regarding the Spring Creek prescribed burns - year 20 post. Moose, WY: U.S. Department of the Interior, National Park Service, Teton Interagency Fire Center. Documents on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92593]
5. Abendroth, Diane. 2018d. Personal communication [Email to Robin Innes]. 5 February. Unpublished 2016 report and information regarding the Teton Science School prescribed burn - year 15 post burn. Moose, WY: U.S. Department of the Interior, National Park Service, Teton Interagency Fire Center. Documents on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92591]
6. Agee, James K. 1994. Fire and weather disturbances in terrestrial ecosystems of the eastern Cascades. In: Everett, Richard L., assessment team leader [Eastside forest ecosystem health assessment]; Hessburg, Paul F., science team leader and tech. ed. [Vol. 3: Assessment]. Gen. Tech. Rep. PNW-GTR-320. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 52 p. [23656]
7. Akinsoji, Aderopo. 1988. Postfire vegetation dynamics in a sagebrush steppe in southeastern Idaho, USA. Vegetatio. 78(3): 151-155. [6944]
8. Alldredge, A. William; Deblinger, Robert D.; Peterson, Jan. 1991. Birth and fawn bed site selection by pronghorns in a sagebrush-steppe community. The Journal of Wildlife Management. 55(2): 222-227. [15468]
9. Allen, Elizabeth A. 2006. Seed banks of pinyon-juniper woodlands: The effects of tree cover and prescribed burn. Reno, NV: University of Nevada, Reno. 77 p. [+ tables & figures]. Thesis. [91181]
10. Allen, Elizabeth A.; Chambers, Jeanne C.; Nowak, Robert S. 2008. Effects of a spring prescribed burn on the soil seed bank in sagebrush steppe exhibiting pinyon-juniper expansion. Western North American Naturalist. 68(3): 265-277. [75588]
11. Allred, B. W. 1941. Grasshoppers and their effect on sagebrush on the Little Power River in Wyoming and Montana. Ecology. 22(4): 387-392. [91049]
12. Anderson, Jay E.; Inouye, Richard S. 2001. Landscape-scale changes in plant species abundance and biodiversity of a sagebrush steppe over 45 years. Ecological Monographs. 71(4): 531-556. [39482]
13. Anderson, Jay E.; Shumar, Mark L. 1986. Impacts of black-tailed jackrabbits at peak population densities on sagebrush vegetation. Journal of Range Management. 39(2): 152-155. [322]
14. Anderson, Loren. 1994. Chapter VII - terrestrial wildlife and habitat. In: Miller, Melanie, ed. Fire effects guide. PMS 481/NFES 2394. Boise, ID: National Wildfire Coordinating Group, Prescribed Fire and Fire Effects Working Team: 1-16. [69984]
15. Anderson, Pete; Boyd, Chad; Chambers, Jeanne; Christiansen, Tom; Davis, Dawn; Espinosa, Shawn; Havlina, Doug; Hopkins, Todd; Ielmini, Michael; Kemner, Don; Kurth, Laurie; Maestas, Jeremy; Mealor, Brian; Mayer, Kenneth; Pellant, Mike; Pyke, David; Tague, Joe; Vernon, Jason. 2015. Invasive plant management and greater sage-grouse conservation: A review and status report with strategic recommendations for improvement. Cheyenne, WY: Western Association of Fish & Wildlife Agencies. 47 p. [89478]
16. Arendt, Paul A. 2012. GLO surveys show change over the past century in a semiarid landscape in the area of Dinosaur National Monument, Colorado/Utah. Laramie, WY: University of Wyoming. 67 p. [+ appendices]. Thesis. [90821]
17. Arkle, Robert S.; Pilliod, David S.; Hanser, Steven E.; Brooks, Matthew L.; Chambers, Jeanne C.; Grace, James B.; Knutson, Kevin C.; Pyke, David A.; Welty, Justin L.; Wirth, Troy A. 2014. Quantifying restoration effectiveness using multi-scale habitat models: Implications for sage-grouse in the Great Basin. Ecosphere. 5(3): 1-32. [90060]
18. Arno, Stephen F.; Gruell, George E. 1983. Fire history at the forest-grassland ecotone in southwestern Montana. Journal of Range Management. 36(3): 332-336. [342]
19. Arno, Stephen F.; Gruell, George E. 1986. Douglas-fir encroachment into mountain grasslands in southwestern Montana. Journal of Range Management. 39(3): 272-276. [343]
20. Asher, Jerry; Dewey, Steven; Olivarez, Jim; Johnson, Curt. 1998. Minimizing weed spread following wildland fires. In: Christianson, Kathy, ed. Western Society of Weed Science: Proceedings; 1998 March 10-12; Waikoloa, HI. In: Proceedings, Western Society of Weed Science. 51: 49. Abstract. [40409]
21. Astroth, Kirk A.; Frischknecht, Neil C. 1984. Managing Intermountain rangelands--research on the Benmore Experimental Range, 1940-84. Gen. Tech, Rep. INT-175. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 44 p. [361]
22. Atamian, Michael T.; Sedinger, James S.; Heaton, Jill S.; Blomberg, Erik J. 2010. Landscape-level assessment of brood rearing habitat for greater sage-grouse in Nevada. The Journal of Wildlife Management. 74(7): 1533-1543. [90141]
23. Autenrieth, Robert E. 1981. Sage grouse management in Idaho. Wildlife Bulletin No. 9. Federal Aid in Wildlife Restoration: Project W-125-R & W-160-R. Boise, ID: Idaho Department of Fish and Game. 238 p. [40588]
24. Autenrieth, Robert. 1976. A study of birth sites selected by pronghorn does and the bed sites of fawns. In: Autenrieth, Robert, compiler. Proceedings, 7th pronghorn antelope workshop; 1976 February 24-26; Twin Falls, ID. Boise, ID: Idaho Department of Fish and Game: 127-134. [25418]
25. Autenrieth, Robert; Molini, William; Braun, Clait, eds. 1982. Sage grouse management practices. Tech. Bull No. 1. Twin Falls, ID: Western States Sage Grouse Committee. 42 p. [7531]
26. Baker, Rollin H. 1968. Habitats and distribution. In: King, John Arthur, ed. Biology of Peromyscus (Rodentia). Special Publication No. 2. Stillwater, OK: The American Society of Mammalogists: 98-126. [25452]
27. Baker, William L. 2002. Indians and fire in the Rocky Mountains: The wilderness hypothesis renewed. In: Vale, Thomas R., ed. Fire, native peoples, and the natural landscape. Washington: Island Press: 41-76. [88508]
28. Baker, William L. 2006. Fire and restoration of sagebrush ecosystems. Wildlife Society Bulletin. 34(1): 177-185. [66367]
29. Baker, William L. 2009. Fire ecology in Rocky Mountain landscapes. Washington, DC: Island Press. 605 p. [88343]
30. Baker, William L. 2009. Fire effects on animals: From individuals to landscapes. In: Fire ecology in Rocky Mountain landscapes. Washington, DC: Island Press: 101-130. [88603]
31. Baker, William L. 2011. Pre-Euro-American and recent fire in sagebrush ecosystems. In: Knick, Stephen T.; Connelly, John W., eds. Greater sage-grouse: Ecology and conservation of a landscape species and its habitats. Studies in Avian Biology No. 38. Berkeley, CA: University of California Press: 185-201. [86464]
32. Balch, Jennifer K.; Bradley, Bethany A.; D'Antonio, Carla M.; Gomez-Dans, Jose. 2013. Introduced annual grass increases regional fire activity across the arid western USA (1980-2009). Global Change Biology. 19(1): 173-183. [86928]
33. Bansal, Sheel; Sheley, Roger L. 2016. Annual grass invasion in sagebrush steppe: The relative importance of climate, soil properties and biotic interactions. Oecologia. 181(2): 543-557. [90652]
34. Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. 2007. Terrestrial vegetation of California, 3rd ed. Berkeley, CA: University of California Press. 712 p. [82605]
35. Barney, Milo A.; Frischknecht, Neil C. 1974. Vegetation changes following fire in the pinyon-juniper type of west-central Utah. Journal of Range Management. 27(2): 91-96. [397]
36. Barrington, Mac; Bunting, Steve; Wright, Gerald. 1988. A fire management plan for Craters of the Moon National Monument. Draft. Subagreement No. 5 to Cooperative Agreement CA-9000-8-0005. Moscow, ID: University of Idaho, Range Resources Department. On file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 52 p. [1687]
37. Baruch-Mordo, Sharon; Evans, Jeffrey S.; Severson, John P.; Naugle, David E.; Maestas, Jeremy D.; Kiesecker, Joseph M.; Falkowski, Michael J.; Hagen, Christian A.; Reese, Kerry P. 2013. Saving sage-grouse from the trees: A proactive solution to reducing a key threat to a candidate species. Biological Conservation. 167: 233-241. [90144]
38. Baskin, Yvonne. 1999. Yellowstone fires: A decade later. Bioscience. 49(2): 93-97. [29468]
39. Bates, Jon D.; Rhodes, Edward C.; Davies, Kirk. 2011. The impacts of fire on sage-grouse habitat and diet resources. Natural Resources and Environmental Issues. 17(15): 111-127. [86086]
40. Bates, Jonathan D.; Davies, Kirk W.; Hulet, April; Miller, Richard F.; Roundy, Bruce. 2017. Sage grouse groceries: Forb response to pinyon-juniper treatments. Rangeland Ecology and Management. 70(1): 106-115. [91592]
41. Bates, Jonathan D.; Davis, Kirk W.; Sharp, Robert N. 2011. Shrub-steppe early succession following juniper cutting and prescribed fire. Environmental Management. 47(3): 468-481. [84612]
42. Bates, Jonathan D.; O'Connor, Rory; Davies, Kirk W. 2014. Vegetation recovery and fuel reduction after seasonal burning of western juniper. Fire Ecology. 10(3): 27-48. [89615]
43. Bates, Jonathan D.; Sharp, Robert N.; Davies, Kirk W. 2014. Sagebrush steppe recovery after fire varies by development phase of Juniperus occidentalis woodland. International Journal of Wildland Fire. 23(1): 117-130. [87793]
44. Bauer, John M. 2006. Fire history and stand structure of a central Nevada pinyon-juniper woodland. Reno, NV: University of Nevada, Reno. 113 p. [+ appendices]. Thesis. [90217]
45. Bauer, John M.; Weisberg, Peter J. 2009. Fire history of a central Nevada pinyon-juniper woodland. Canadian Journal of Forest Research. 39(8): 1589-1599. [83029]
46. Beardall, Louis E.; Sylvester, Vern E. 1976. Spring burning for removal of sagebrush competition in Nevada. In: Proceedings, Tall Timbers fire ecology conference: Fire and land management symposium; 1974 October 8-10; Missoula, MT. No. 14. Tallahassee, FL: Tall Timbers Research Station: 539-547. [406]
47. Beck, D. I. 1975. Attributes of a wintering population of sage grouse, North Park, Colorado. Fort Collins, CO: Colorado State University. 49 p. Thesis. [5757]
48. Beck, Jeffrey L.; Klein, J. Garrett; Wright, Justin; Wolfley, Kenneth P. 2011. Potential and pitfalls of prescribed burning big sagebrush habitat to enhance nesting and early brood-rearing habitats for greater sage-grouse. In: Wambolt, Carl L.; Kitchen, Stanley G.; Frisina, Michael R.; Sowell, Bok; Keigley, Richard B.; Palacios, Patsy; Robinson, Jill, comps. Proceedings--shrublands: Wildlands and wildlife habitats; 15th wildland shrub symposium; 2008 June 17-19; Bozeman, MT. Natural Resources and Environmental Issues, Volume XVI. Logan, UT: Utah State University, College of Natural Resources, S. J. and Jessie E. Quinney Natural Resources Research Library: 27-32. [83465]
49. Beck, Jeffrey L.; Mitchell, Dean L. 2000. Influences of livestock grazing on sage grouse habitat. Wildlife Society Bulletin. 28(4): 993-1002. [90146]
50. Beckstead, Julie; Augspurger, Carol K. 2004. An experimental test of resistance to cheatgrass invasion: Limiting resources at different life stages. Biological Invasions. 6(4): 417-432. [91651]
51. Beckstead, Julie; Street, Laura E.; Meyer, Susan E.; Allen, Phil S. 2011. Fire effects on the cheatgrass seed bank pathogen Pyrenophora semeniperda. Rangeland Ecology & Management. 64(2): 148-157. [91159]
52. Beetle, A. A. 1960. A study of sagebrush: The section Tridentatae of Artemisia. Bulletin 368. Laramie, WY: University of Wyoming, Agricultural Experiment Station. 83 p. [416]
53. Beetle, Alan A. 1961. Range survey in Teton County, Wyoming: Part 1. Ecology of range resources. Bull. 376. Laramie, WY: University of Wyoming, Agricultural Experiment Station. 42 p. [417]
54. Beetle, Alan A.; Johnson, Kendall L. 1982. Sagebrush in Wyoming. Bull. 779. Laramie, WY: University of Wyoming, Agricultural Experiment Station. 68 p. [421]
55. Beetle, Alan A.; Young, Alvin. 1965. A third subspecies in the Artemisia tridentata complex. Rhodora. 67: 405-406. [422]
56. Belnap, Jayne; Kaltenecker, Julie Hilty; Rosentreter, Roger; Williams, John; Leonard, Steve; Eldridge, David. 2001. Biological soil crusts: Ecology and management. Technical Reference 1730-2. Denver, CO: U.S. Department of the Interior, Bureau of Land Management, National Science and Technology Center, Information and Communications Group. 110 p. [40277]
57. Belnap, Jayne; Williams, John; Kaltenecker, Julie. 1999. Structure and function of biological soil crusts. In: Meurisse, Robert T.; Ypsilantis, William G.; Seybold, Cathy, tech. eds. Proceedings: Pacific Northwest forest and rangeland soil organism symposium; 1998 March 17-19; Corvallis, OR. Gen. Tech. Rep. PNW-GTR-461. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 161-178. [38449]
58. Bendell, J. F. 1974. Effects of fire on birds and mammals. In: Kozlowski, T. T.; Ahlgren, C. E., eds. Fire and ecosystems. New York: Academic Press: 73-138. [16447]
59. Benson, Lee A.; Braun, Clait E.; Leininger, Wayne C. 1991. Sage grouse response to burning in the big sagebrush type. In: Davis, P.; Rush, S.; Todd, J., eds.; Comer, Robert D., contributor. Issues and technology in the management of impacted wildlife: Proceedings of a national symposium; 1991 April 8-10; Snowmass Resort, CO. Boulder, CO: Thorne Ecological Institute: 97-104. [21766]
60. Beyers, Jan L. 2004. Postfire seeding for erosion control: Effectiveness and impacts on native plant communities. Conservation Biology. 18(4): 947-956. [50079]
61. Bilbrough, C. J.; Richards, J. H. 1991. Branch architecture of sagebrush and bitterbrush: Use of a branch complex to describe and compare patterns of growth. Canadian Journal of Botany. 69: 1288-1295. [15755]
62. Billings, W. D. 1994. Ecological impacts of cheatgrass and resultant fire on ecosystems in the western Great Basin. In: Monsen, Stephen B.; Kitchen, Stanley G., comps. Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 22-30. [24248]
63. Blackburn, W. H.; Beall, R.; Bruner, A.; Klebenow, D.; Mason, R.; Roundy, B.; Stager, W.; Ward, K. 1975. Controlled fire as a management tool in the pinyon-juniper woodland, Nevada. Annual Progress Report FY 1975. Unpublished report on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 77 p. [453]
64. Blaisdell, James P. 1949. Competition between sagebrush seedlings and reseeded grasses. Ecology. 30(4): 512-519. [461]
65. Blaisdell, James P. 1953. Ecological effects of planned burning of sagebrush-grass range on the upper Snake River Plains. Tech. Bull. 1975. Washington, DC: U.S. Department of Agriculture. 39 p. [462]
66. Blaisdell, James P.; Murray, Robert B.; McArthur, E. Durant. 1982. Managing Intermountain rangelands--sagebrush-grass ranges. Gen. Tech. Rep. INT-134. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 41 p. [467]
67. Blank, R. R.; Chambers, J.; Roundy, B.; Whittaker, A. 2007. Nutrient availability in rangeland soils: Influence of prescribed burning, herbaceous vegetation removal, overseeding with Bromus tectorum, season, and elevation. Rangeland Ecology & Management. 60(6): 644-655. [74042]
68. Blank, Robert R.; Allen, Fay; Young, James A. 1994. Extractable anions in soils following wildfire in a sagebrush-grass community. Soil Science Society of America Journal. 58(2): 564-570. [43173]
69. Blank, Robert R.; Allen, Fay; Young, James A. 1994. Growth and elemental content of several sagebrush-steppe species in unburned and post-wildfire soil and plant effects on soil attributes. Plant and Soil. 164(1): 35-41. [26887]
70. Board, David I.; Chambers, Jeanne C.; Wright, Joan G. 2011. Effects of spring prescribed fire in expanding pinyon-juniper woodlands on seedling establishment of sagebrush species. In: Wambolt, Carl L.; Kitchen, Stanley G.; Frisina, Michael R.; Sowell, Bok; Keigley, Richard B.; Palacios, Patsy; Robinson, Jill, comps. Proceedings--shrublands: Wildlands and wildlife habitats; 15th wildland shrub symposium; 2008 June 17-19; Bozeman, MT. Natural Resources and Environmental Issues, Volume XVI. Logan, UT: Utah State University, College of Natural Resources, S. J. and Jessie E. Quinney Natural Resources Research Library: 149-158. [83480]
71. Bock, Carl E.; Bock, Jane H. 1987. Avian habitat occupancy following fire in a Montana shrubsteppe. Prairie Naturalist. 19(3): 153-158. [2806]
72. Bonham, C. D.; Cottrell, T. R.; Mitchell, J. E. 1991. Inferences for life history strategies of Artemisia tridentata subspecies. Journal of Vegetation Science. 2(3): 339-344. [16599]
73. Booth, Gordon D.; Welch, Bruce L.; Jacobson, Tracy L. C. 1990. Seedling growth rate of 3 subspecies of big sagebrush. Journal of Range Management. 43(5): 432-436. [11005]
74. Bourne, Andrea; Bunting, Stephen C. 2011. Guide for quantifying post-treatment fuels in the sagebrush steppe and juniper woodlands of the Great Basin. Tech. Note 437. Denver, CO: U.S. Department of the Interior, Bureau of Land Management. 115 p. [88012]
75. Brabec, Martha M. 2014. Big sagebrush (Artemisia tridentata) in a shifting climate context: Assessment of seedling responses to climate. Boise, ID: Boise State University. 116 p. Thesis. [88557]
76. Brabec, Martha M.; Germino, Matthew J.; Richardson, Bryce A. 2017. Climate adaption and post-fire restoration of a foundational perennial in cold desert: Insights from intraspecific variation in response to weather. Journal of Applied Ecology. 54(1): 293-302. [91475]
77. Bradfield, Terry D. 1975. On the behavior and ecology of the pygmy rabbit. Pocatello, ID: Idaho State University. 43 p. Thesis. [23639]
78. Bradley, Bethany A. 2010. Assessing ecosystem threats from global and regional change: Hierarchical modeling of risk to sagebrush ecosystems from climate change, land use and invasive species in Nevada, USA. Ecography. 33: 198-208. [91466]
79. Braun, Clait E. 1998. Sage grouse declines in western North America: What are the problems? In: Proceedings, WAFWA; 1998, June 26-July 2; Jackson, WY. Cheyenne, WY: Western Association of Fish and Wildlife Agencies: 139-156. [35365]
80. Braun, Clait E.; Connelly, John W.; Schroeder, Michael A. 2005. Seasonal habitat requirements for sage-grouse: Spring, summer, fall, and winter. In: Shaw, Nancy L.; Pellant, Mike; Monsen, Stephen B., eds. Sage-grouse habitat restoration symposium proceedings; 2001 June 4-7; Boise, ID. Proc. RMRS-P-38. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 38-42. [63180]
81. Breshears, David D.; Cobb, Niel S.; Rich, Paul M.; Price, Kevin P.; Allen, Craig D.; Balice, Randy G.; Romme, William H.; Kastens, Jude H.; Floyd, M. Lisa; Belnap, Jayne; Anderson, Jesse J.; Myers, Orrin B.; Meyer, Clifton W. 2005. Regional vegetation die-off in response to global-change-type drought. Proceedings of the National Academy of Sciences. 102(42): 15144-15148. [91632]
82. Brewerton, Adam B. 2012. Avian response to post wildland fire reseeding treatments in Great Basin shrubsteppe. Logan, UT: Utah State University. 41 p. Thesis. [88042]
83. Briske, D. D.; Bestelmeyer, B. T.; Stringham, T. K.; Shaver, P. L. 2008. Forum: Recommendations for development of resilience-based state-and-transition models. Rangeland Ecology and Management. 61(4): 359-367. [91601]
84. Britton, Carlton M.; Clark, Robert G. 1985. Effects of fire on sagebrush and bitterbrush. In: Saunders, Ken; Durham, Jack; [and others], eds. Rangeland fire effects: Proceedings of the symposium; 1984 November 27-29; Boise, ID. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office: 22-26. [515]
85. Britton, Carlton M.; Clark, Robert G.; Sneva, Forrest A. 1981. Will your sagebrush range burn? Rangelands. 3(5): 207-208. [517]
86. Brooks, Mathew L.; Chambers, Jeanne C. 2011. Resistance to invasion and resilience to fire in desert shrublands of North America. Rangeland Ecology and Management. 64(5): 431-438. [84701]
87. Brooks, Matthew L. 2008. Effects of fire suppression and postfire management activities on plant invasions. In: Zouhar, Kristin; Smith, Jane Kapler; Sutherland, Steve; Brooks, Matthew L., eds. Wildland fire in ecosystems: Fire and nonnative invasive plants. Gen. Tech. Rep. RMRS-GTR-42-vol. 6. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 269-280. [70909]
88. Brooks, Matthew L.; D'Antonio, Carla M.; Richardson, David M.; Grace, James B.; Keeley, Jon E.; DiTomaso, Joseph M.; Hobbs, Richard J.; Pellant, Mike; Pyke, David. 2004. Effects of invasive alien plants on fire regimes. BioScience. 54(7): 677-688. [50224]
89. Brooks, Matthew L.; Pyke, David A. 2001. Invasive plants and fire in the deserts of North America. In: Galley, Krista E. M.; Wilson, Tyrone P., eds. Proceedings of the invasive species workshop: The role of fire in the control and spread of invasive species; Fire conference 2000: 1st national congress on fire ecology, prevention, and management; 2000 November 27 - December 1; San Diego, CA. Misc. Publ. No. 11. Tallahassee, FL: Tall Timbers Research Station: 1-14. [40491]
90. Brown, James K. 1982. Fuel and fire behavior prediction in big sagebrush. Research Paper INT-290. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 10 p. [543]
91. Brown, James K. 1985. Fire effects and application of prescribed fire in aspen. In: Saunders, Ken; Durham, Jack; [and others], eds. Rangeland fire effects: Proceedings of the symposium; 1984 November 27-29; Boise, ID. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office: 38-47. [3658]
92. Brown, James K. 1985. Role and use of fire in aspen. In: Foresters' future: Leaders or followers?: Proceedings of the 1985 Society of American Foresters national convention; 1985 July 28-31; Fort Collins, CO. SAF Publ. 85-13. Bethesda, MD: Society of American Foresters: 101-105. [5104]
93. Brown, Lindsay J. 2007. A historic perspective: The response of breeding passerines to rangeland alteration in Rich County, Utah. Logan, UT: Utah State University. 57 p. [+ appendices]. Thesis. [91422]
94. Bruner, Allen D.; Klebenow, Donald A. 1979. Predicting success of prescribed fires in pinyon-juniper woodland in Nevada. Res. Pap. INT-219. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 11 p. [3254]
95. Bryant, Fred C. 1991. Managed habitats for deer in juniper woodlands of west Texas. In: Rodiek, Jon E.; Bolen, Eric G., eds. Wildlife and habitats in managed landscapes. Island Press: Washington, DC: 56-75. [85242]
96. Bukowski, Beth E.; Baker, William L. 2013. Historical fire in sagebrush landscapes of the Gunnison sage-grouse range from land-survey records. Journal of Arid Environments. 98: 1-9. [87644]
97. Bukowski, Beth E.; Baker, William L. 2013. Historical fire regimes, reconstructed from land-survey data, led to complexity and fluctuation in sagebrush landscapes. Ecological Applications. 23(3): 546-564. [88857]
98. Bunting, Stephen C. 1984. Prescribed burning of live standing western juniper and post-burning succession. In: Proceedings--western juniper management short course; 1984 October 15-16; Bend, OR. Corvallis, OR: Oregon State University, Extension Service; Oregon State University, Department of Rangeland Resources; 69-73. [557]
99. Bunting, Stephen C. 1987. Use of prescribed burning in juniper and pinyon-juniper woodlands. In: Everett, Richard L., comp. Proceedings--pinyon-juniper conference; 1986 January 13-16; Reno, NV. Gen. Tech. Rep. INT-215. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 141-144. [4836]
100. Bunting, Stephen C.; Kilgore, Bruce M.; Bushey, Charles L. 1987. Guidelines for prescribed burning sagebrush-grass rangelands in the northern Great Basin. Gen. Tech. Rep. INT-231. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 33 p. [5281]
101. Bunting, Stephen C.; Kingery, James L.; Hemstrom, Miles A.; Schroeder, Michael A.; Gravenmier, Rebecca A.; Hann, Wendel J. 2002. Altered rangeland ecosystems in the interior Columbia Basin. Gen. Tech. Rep. PNW-GTR-553. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 71 p. (Quigley, Thomas M., ed.; Interior Columbia Basin Ecosystem Project: scientific assessment). [43462]
102. Bunting, Stephen C.; Peters, Erin F.; Sapsis, David B. 1994. Impact of fire management on rangelands of the Intermountain West. Scientific Contract Report: Science Integration Team, Terrestrial Staff, Range Task Group. Walla Walla, WA: Interior Columbia Basin Ecosystem Management Project. 32 p. [26452]
103. Bunting, Stephen C.; Strand, Eva K.; Kingery, James L. 2007. Landscape characteristics of sagebrush-steppe/juniper woodland mosaics under various modeled prescribed fire regimes. In: Masters, Ronald E.; Galley, Krista E. M., eds. Fire in grassland and shrubland ecosystems: Proceedings of the 23rd Tall Timbers fire ecology conference; 2005 October 17-20; Bartlesville, OK. Tallahassee, FL: Tall Timbers Research Station: 50-57. [69733]
104. Buranek, Shelley. 2017. Personal communication [Email to Robin Innes]. 19 January. Regarding use of Rx fire in big sagebrush communities within the John Day Fossil Beds National Monument. Casper, OR: U.S. Department of the Interior, National Park Service, John Day Fossil Beds National Monument. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [91633]
105. Burke, Ingrid C. 1989. Control of nitrogen mineralization in a sagebrush steppe landscape. Ecology. 70(4): 1115-1126. [7974]
106. Burke, Ingrid C.; Reiners, William A.; Olson, Richard K. 1989. Topographic control of vegetation in a mountain big sagebrush steppe. Vegetatio. 84(2): 77-86. [11178]
107. Burke, Ingrid C.; Reiners, William A.; Schimel, David S. 1989. Organic matter turnover in a sagebrush steppe landscape. Biogeochemistry. 7(1): 11-31. [11133]
108. Burkhardt, J. Wayne; Tisdale, E. W. 1969. Nature and successional status of western juniper vegetation in Idaho. Journal of Range Management. 22(4): 264-270. [564]
109. Burkhardt, Wayne J.; Tisdale, E. W. 1976. Causes of juniper invasion in southwestern Idaho. Ecology. 57(3): 472-484. [565]
110. Buseck, Rebecca S.; Keinath, Douglas A.; McGee, Matthew H. 2004. Species assessment for sage thrasher (Oreoscoptes montanus) in Wyoming. Cheyenne, WY: U.S. Department of the Interior, Bureau of Land Management, Wyoming State Office. 72 p. [91430]
111. Bushey, Charles L. 1985. Summary of results from the Galena Gulch 1982 spring burns (Units 1b). Missoula, MT: Systems for Environmental Management. Unpublished report on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 9 p. [567]
112. Caldwell, Martyn M.; Eissenstat, David M.; Richards, James H.; Allen, Michael F. 1985. Competition for phosphorus: Differential uptake from dual-isotope- labeled soil interspaces between shrub and grass. Science. 229: 384-385. [586]
113. Call, Mayo W. 1979. Habitat requirements and management recommendations for sage grouse. Denver, CO: U.S. Department of the Interior, Bureau of Land Management, Denver Service Center. 37 p. [591]
114. Call, Mayo W.; Maser, Chris. 1985. Wildlife habitats in managed rangelands--the Great Basin of southeastern Oregon: Sage grouse. Gen. Tech. Rep. PNW-187. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 30 p. [592]
115. Carpenter, Alan T.; West, Neil E. 1987. Indifference of mountain big sagebrush growth to supplemental water and nitrogen. Journal of Range Management. 40(5): 448-451. [3320]
116. Casazza, Michael L.; Coates, Peter S.; Overton, Cory T. 2011. Linking habitat selection and brood success in greater sage-grouse. In: Sandercock, Brett K.; Martin, Kathy; Segelbacher, Gernot, eds. Ecology, conservation, and management of grouse. Studies in Avian Biology, No. 39. Berkeley, CA: University of California Press: 151-167. [90143]
117. Castrale, John S. 1982. Effects of two sagebrush control methods on nongame birds. The Journal of Wildlife Management. 46(4): 945-952. [5254]
118. Chambers, Jeanne C. 1989. Native species establishment on an oil drillpad site in the Uintah Mountains, Utah: Effects of introduced grass density and fertilizer. Res. Pap. INT-402. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 9 p. [6885]
119. Chambers, Jeanne C. 2008. Climate change and the Great Basin. Gen. Tech. Rep. RMRS-GTR-204. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 32 pages. [91464]
120. Chambers, Jeanne C.; Bradley, Bethany A.; Brown, Cynthia S.; D'Antonio, Carla; Germino, Matthew J.; Grace, James B.; Hardegree, Stuart P.; Miller, Richard F.; Pyke, David A. 2014. Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America. Ecosystems. 17(2): 360-375. [91554]
121. Chambers, Jeanne C.; Miller, Richard F.; Board, David I.; Pyke, David A.; Roundy, Bruce A.; Grace, James B.; Schupp, Eugene W.; Tausch, Robin J. 2014. Resilience and resistance of sagebrush ecosystems: Implications for state and transition models and management treatments. Rangeland Ecology and Management. 67(5): 440-454. [89207]
122. Chambers, Jeanne C.; Pyke, David A.; Maestas, Jeremy D.; Pellant, Mike; Boyd, Chad S.; Campbell, Steven B.; Espinosa, Shawn; Havlina, Douglas W.; Mayer, Kenneth E.; Wuenschel, Amarina. 2014. Using resistance and resilience concepts to reduce impacts of invasive annual grasses and altered fire regimes on the sagebrush ecosystem and greater sage-grouse: A strategic multi-scale approach. Gen. Tech. Rep. RMRS-GTR-326. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 73 p. [89518]
123. Chambers, Jeanne C.; Roundy, Bruce A.; Blank, Robert R.; Meyer, Susan E.; Whittaker, A. 2007. What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum? Ecological Monographs. 77(1): 117-145. [66809]
124. Chaplin, M. R.; Winward, A. H. 1982. The effect of simulated fire on emergence of seeds found in the soil of big sagebrush communities. In: Society for Range Management Abstracts: Proceedings, 35th Annual Meeting of the Society for Range Management; [Date of conference unknown]; Calgary, AB. Denver, CO: Society for Range Management: 37. Abstract. [9800]
125. Chaplin, Mark R. 1982. Big sagebrush (Artemisia tridentata) ecology and management with emphasis on prescribed burning. Corvallis, OR: Oregon State University. 136 p. Dissertation. [9484]
126. Chapman, Joseph A.; Feldhamer, George A., eds. 1982. Wild mammals of North America. Baltimore, MD: The Johns Hopkins University Press. 1147 p. [21085]
127. Christensen, Earl M.; Johnson, Hyrum B. 1964. Presettlement vegetation and vegetational change in three valleys in central Utah. Brigham Young University Science Bulletin, Biological Series; 4(4). Provo, UT: Brigham Young University. 16 p. [627]
128. Christianson, David A.; Creel, Scott. 2007. Review of environmental factors affecting elk winter diets. The Journal of Wildlife Management. 71(1): 164-176. [83214]
129. Clark, Patrick E.; Lee, Jaechoul; Ko, Kyungduk; Nielson, Ryan M.; Johnson, Douglas E.; Ganskopp, David C.; Pierson, Frederick B.; Hardegree, Stuart P. 2014. Prescribed fire effects on resource selection by cattle in mesic sagebrush steppe. Part 1: Spring grazing. Journal of Arid Environments. 124: 78-88. [89948]
130. Clark, Patrick E.; Lee, Jaechoul; Ko, Kyungduk; Nielson, Ryan M.; Johnson, Douglas E.; Ganskopp, David C.; Pierson, Frederick B.; Hardegree, Stuart P. 2016. Prescribed fire effects on resource selection by cattle in mesic sagebrush steppe. Part 2: Mid-summer grazing. Journal of Arid Environments. 124: 398-412. [89947]
131. Clark, Robert G.; Starkey, Edward E. 1990. Use of prescribed fire in rangeland ecosystems. In: Walstad, John D.; Radosevich, Steven R.; Sandberg, David V., eds. Natural and prescribed fire in Pacific Northwest forests. Corvallis, OR: Oregon State University Press: 81-91. [46959]
132. Clarke, Sharon E.; Garner, Mark W.; McIntosh, Bruce A.; Sedell, James R. 1997. Section 3--Landscape-level ecoregions for seven contiguous watersheds, northeast Oregon and southeast Washington. In: Clarke, Sharon E.; Bryce, Sandra A., eds. Hierarchical subdivisions of the Columbia Plateau and Blue Mountains ecoregions, Oregon and Washington. Gen. Tech. Rep. PNW-GTR-395. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 53-113. [28539]
133. Cleary, M. B.; Pendall, E.; Ewers, B. E. 2008. Testing sagebrush allometric relationships across three fire chronosequences in Wyoming, USA. Journal of Arid Environments. 72(4): 285-301. [69951]
134. Cluff, L. K.; Welch, B. L.; Pederson, J. C.; Brotherson, J. D. 1982. Concentration of monoterpenoids in the rumen ingesta of wild mule deer. Journal of Range Management. 35(2): 192-194. [91419]
135. Coates, Peter S.; Prochazka, Brian G.; Ricca, Mark A.; Gustafson, K. Ben; Ziegler, Pilar; Casazza, Michael L. 2016. Pinyon and juniper encroachment into sagebrush ecosystems impacts distribution and survival of greater sage-grouse. Rangeland Ecology and Management. 70(1): 25-38. [91613]
136. Cogan, Dan; Taylor, Jeanne E.; Schulz, Keith. 2012. Vegetation Inventory Project: Great Basin National Park. Natural Resource Rep. NPS/MOJN/NRR-2012/568. Fort Collins, CO: U.S. Department of the Interior, National Park Service. 79 p. [+ appendices]. [89265]
137. Colorado Parks and Wildlife. 2005. Sagebrush vole (Lemmiscus curtatus), [Online]. Colorado sagebrush: A conservation assessment and strategy. A82-A90. Colorado Parks & Wildlife (Producer). Available: https://cpw.state.co.us/Documents/WildlifeSpecies/Sagebrush/SagebrushVole.pdf [2017, January 12]. [91439]
138. Condon, Lea A.; Weisberg, Peter J. 2016. Topographic context of the burn edge influences postfire recruitment of arid land shrubs. Rangeland Ecology & Management. 69(2): 129-133. [91203]
139. Condon, Lea; Weisberg, Peter J.; Chambers, Jeanne C. 2011. Abiotic and biotic influences on Bromus tectorum invasion and Artemisia tridentata recovery after fire. International Journal of Wildland Fire. 20(4): 597-604. [83125]
140. Connelly, John W.; Reese, Kerry P.; Fischer, Richard A.; Wakkinen, Wayne L. 2000. Response of a sage grouse breeding population to fire in southeastern Idaho. Wildlife Society Bulletin. 28(1): 90-96. [35803]
141. Connelly, John W.; Rinkes, E. Thomas; Braun, Clait E. 2011. Characteristics of greater sage-grouse habitats: A landscape species at micro- and macroscales. In: Knick, Steven T.; Connelly, John W., eds. Greater sage-grouse: Ecology and conservation of a landscape species and its habitats. Studies in Avian Biology, No. 38. Berkeley, CA: University of California Press: 69-83. [89612]
142. Connelly, John W.; Schroeder, Michael A.; Sands, Alan R.; Braun, Clait E. 2000. Guidelines to manage sage grouse populations and their habitats. Wildlife Society Bulletin. 28(4): 967-985. [89610]
143. Cook, John G. 2002. Nutrition and food. In: Toweill, Dale E.; Thomas, Jack Ward, eds. North American elk: Ecology and management. Washington, DC: Smithsonian Institution Press: 259-350. [81796]
144. Cook, John G.; Hershey, Terry J.; Irwin, Larry L. 1994. Vegetative response to burning on Wyoming mountain-shrub big game ranges. Journal of Range Management. 47(4): 296-302. [23449]
145. Cottam, W. P.; Stewart, George. 1940. Plant succession as a result of grazing and of meadow desiccation by erosion since settlement in 1862. Journal of Forestry. 38(8): 613-626. [696]
146. Cottrell, Thomas R.; Bonham, Charles D. 1992. Characteristics of sites occupied by subspecies of Artemisia tridentata in the Piceance Basin, Colorado. The Great Basin Naturalist. 52(2): 174-178. [19688]
147. Cotts, N. R.; Redente, E. F.; Schiller, R. 1991. Restoration methods for abandoned roads at lower elevations in Grand Teton National Park, Wyoming. Arid Soil Research and Rehabilitation. 5(4): 235-249. [16995]
148. Cox, Robert D.; Anderson, Val Jo. 2004. Increasing native diversity of cheatgrass dominated rangeland through assisted succession. Journal of Range Management. 57(2): 203-210. [91652]
149. Crawford, John A.; Olson, Rich A.; West, Neil E.; Mosley, Jeffrey C.; Schroeder, Michael A.; Whitson, Tom D.; Miller, Richard F.; Gregg, Michael A.; Boyd, Chad S. 2004. Ecology and management of sage-grouse and sage-grouse habitat. Journal of Range Management. 57(1): 2-19. [47019]
150. Crawford, John Earl, Jr. 1960. The movements, productivity, and management of sage grouse in Clark and Fremont Counties, Idaho. Moscow, ID: University of Idaho. 85 p. Thesis. [5845]
151. Crawford, Rex C.; Chappell, Christopher B. 2001. Subalpine parkland. In: Chappell, Christopher B.; Crawford, Rex C.; Barrett, Charley; Kagan, Jimmy; Johnson, David H.; O'Mealy, Mikell; Green, Greg A.; Ferguson, Howard L.; Edge, W. Daniel; Greda, Eva L.; O'Neil, Thomas A. Wildlife habitats: Descriptions, status, trends, and system dynamics. In: Johnson, David H.; O'Neil, Thomas A., managing directors. Wildlife-habitat relationships in Oregon and Washington. Corvallis, OR: Oregon State University Press: 38-40. [68098]
152. Crawford, Rex C.; Kagan, Jimmy. 2001. 13. Western juniper and mountain mahogany woodlands. In: Chappell, Christopher B.; Crawford, Rex C.; Barrett, Charley; Kagan, Jimmy; Johnson, David H.; O'Mealy, Mikell; Green, Greg A.; Ferguson, Howard L.; Edge, W. Daniel; Greda, Eva L.; O'Neil, Thomas A. Wildlife habitats: Descriptions, status, trends, and system dynamics. In: Johnson, David H.; O'Neil, Thomas A., managing directors. Wildlife-habitat relationships in Oregon and Washington. Corvallis, OR: Oregon State University Press: 45-46. [67917]
153. Crawford, Rex C.; Kagan, Jimmy. 2001. 16. Shrub-steppe. In: Chappell, Christopher B.; Crawford, Rex C.; Barrett, Charley; Kagan, Jimmy; Johnson, David H.; O'Mealy, Mikell; Green, Greg A.; Ferguson, Howard L.; Edge, W. Daniel; Greda, Eva L.; O'Neil, Thomas A. Wildlife habitats: Descriptions, status, trends, and system dynamics. In: Johnson, David H.; O'Neil, Thomas A., managing directors. Wildlife-habitat relationships in Oregon and Washington. Corvallis, OR: Oregon State University Press: 50-51. [68105]
154. Creutzburg, Megan K.; Halofsky, Jessica E.; Halofsky, Joshua S.; Christopher, Treg A. 2014. Climate change and land management in the rangelands of central Oregon. Environmental Management. 55(1): 43-55. [90145]
155. Cronquist, Arthur; Holmgren, Arthur H.; Holmgren, Noel H.; Reveal, James L.; Holmgren, Patricia K. 1994. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Vol. 5: Asterales. New York: The New York Botanical Garden. 496 p. [28653]
156. D'Antonio, Carla M.; Vitousek, Peter M. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics. 23: 63-87. [20148]
157. Dalhgren, David K.; Larsen, Randy T.; Danvir, Rick; Wilson, George; Thacker, Eric T.; Black, Todd A.; Naugle, David E.; Connelly, John W.; Messmer, Terry A. 2015. Greater sage-grouse and range management: Insights from a 25-year case study in Utah and Wyoming. Rangeland Ecology and Management. 68(5): 375-382. [90435]
158. Dalke, Paul D.; Pyrah, Duane B.; Stanton, Don C.; Crawford, John E.; Schlatterer, Edward F. 1963. Ecology, productivity, and management of sage grouse in Idaho. The Journal of Wildlife Management. 27(4): 810-841. [5975]
159. Daubenmire, Rexford F. 1975. Ecology of Artemisia tridentata subsp. tridentata in the state of Washington. Northwest Science. 49(1): 24-35. [744]
160. Daubenmire, Rexford. 1981. Subalpine parks associated with snow transfer in the mountains of northern Idaho and eastern Washington. Northwest Science. 55(2): 124-135. [8273]
161. Davies, K. W.; Bates, J. D.; Madsen, M. D.; Nafus, A. M. 2014. Restoration of mountain big sagebrush steppe following prescribed burning to control western juniper. Environmental Management. 53(5): 1015-1022. [87988]
162. Davies, K. W.; Bates, J. D.; Nafus, A. M. 2012. Comparing burned and mowed treatments in mountain big sagebrush steppe. Environmental Management. 50(3): 541-461. [87396]
163. Davies, Kirk W.; Bates, Jon D. 2017. Restoring big sagebrush after controlling encroaching western juniper with fire: Aspect and subspecies effects. Restoration Ecology. 25(1): 33-41. [91446]
164. Davies, Kirk W.; Bates, Jon D.; Boyd, Chad S.; Nafus, Aleta M. 2014. Is fire exclusion in mountain big sagebrush communities prudent? Soil nutrient, plant diversity and arthropod response to burning. International Journal of Wildland Fire. 48(3): 417-424. [89706]
165. Davies, Kirk W.; Bates, Jonathan D.; Johnson, Dustin D.; Nafus, Aleta M. 2009. Influence of mowing Artemisia tridentata spp. wyomingensis on winter habitat for wildlife. Environmental Management. 44(1): 84-92. [91448]
166. Davies, Kirk W.; Bates, Jonathan D.; Miller, Richard F. 2008. Effects of burning Wyoming big sagebrush-bunchgrass communities. In: Davies, Kirk W.; Nafus, Aleta M., comps. Sagebrush steppe--Research Progress Report 2007. ARS Publication EOARC. Burns, OR: U.S. Department of Agriculture, Agricultural Research Service: 4-9. [86222]
167. Davies, Kirk W.; Bates, Jonathan D.; Svejcar, Tony J.; Boyd, Chad S. 2010. Effects of long-term livestock grazing on fuel characteristics in rangelands: An example from the sagebrush steppe. Rangeland Ecology & Management. 63(6): 662-669. [82380]
168. Davies, Kirk W.; Boyd, Chad S.; Beck, Jeffrey L.; Bates, Jon D.; Svejcar, Tony J.; Gregg, Michael A. 2011. Saving the sagebrush sea: An ecosystem conservation plan for big sagebrush plant communities. Biological Conservation. 144(11): 2573-2584. [83899]
169. Davis, Dawn M.; Crawford, John A. 2014. Case study: Short-term response of greater sage-grouse habitats to wildfire in mountain big sagebrush communities. Wildlife Society Bulletin. 39(1): 1-9. [90066]
170. Davison, Jason. 1996. Livestock grazing in wildland fuel management programs. Rangelands. 18(6): 242-245. [28895]
171. Dealy, J. Edward; Leckenby, Donavin A.; Concannon, Diane M. 1981. Wildlife habitats on managed rangelands--the Great Basin of southeastern Oregon: Plant communities and their importance to wildlife. Gen. Tech. Rep. PNW-120. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest and Range Experiment Station. 66 p. [786]
172. Dean, Sheila; Burkhardt, J. Wayne; Meeuwig, Richard O. 1981. Estimating twig and foliage biomass of sagebrush, bitterbrush, and rabbitbrush in the Great Basin. Journal of Range Management. 34(3): 224-227. [787]
173. DeByle, Norbert V. 1985. Managing wildlife habitat with fire in the aspen ecosystem. In: Lotan, James E.; Brown, James K., compilers. Fire's effects on wildlife habitat--symposium proceedings; 1984 March 21; Missoula, MT. Gen. Tech. Rep. INT-186. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 73-82. [8336]
174. Deming, O. V. 1963. Antelope and sagebrush. In: Yoakum, Jim, compiler. Transactions, Interstate antelope conference; 1963 December 4-5; Alturas, CA. Reno, NV: Interstate Antelope Conference: 55-60. [25687]
175. Diamond, Joel M.; Call, Christopher A.; Devoe, Nora. 2009. Effects of targeted cattle grazing on fire behavior of cheatgrass-dominated rangeland in the northern Great Basin, USA. International Journal of Wildland Fire. 18(8): 944-950. [81834]
176. DiCristina, Katherine; Germino, Matthew. 2006. Correlation of neighborhood relationships, carbon assimilation, and water status of sagebrush seedlings establishing after fire. Western North American Naturalist. 66(4): 441-449. [66439]
177. DiCristina, Katherine; Germino, Matthew; Seefeldt, Steven. 2006. Spatial and temporal patterns of sagebrush (Artemisia tridentata ssp. vaseyana) establishment following fire. In: Weber, Keith T., ed. Final Report: Detection, prediction, impact, and management of invasive plants using GIS. Pocatello, ID: Idaho State University: 99-110. [90892]
178. Dicus, Christopher, A.; Delfino, Kenneth; Weise, David R. 2009. Predicted fire behavior and societal benefits in three eastern Sierra Nevada vegetation types. Fire Ecology. 5(1): 67-78. [81039]
179. Dittberner, Phillip L.; Olson, Michael R. 1983. The Plant Information Network (PIN) data base: Colorado, Montana, North Dakota, Utah, and Wyoming. FWS/OBS-83/86. Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service. 786 p. [806]
180. Dorn, Robert D. 1988. Vascular plants of Wyoming. Cheyenne, WY: Mountain West Publishing. 340 p. [6129]
181. Dorrance, Michael J. 1967. A literature review on behavior of mule deer. Special Report Number 7. [Denver, CO]: Colorado Department of Game, Fish, and Parks, Game Research Division; Colorado Cooperative Wildlife Research Unit. 26 p. [85019]
182. Duncan, Corinne M. 2008. Seed bank response to juniper expansion in the semi-arid lands of Oregon, USA. Corvallis, OR: Oregon State University. 81 p. Thesis. [88013]
183. Earnst, Susan L.; Newsome, Heidi L.; LaFromboise, William L.; LaFromboise, Nancy. 2009. Avian response to wildfire in Interior Columbia Basin shrubsteppe. The Condor. 111(2): 370-376. [91431]
184. Eberhardt, L. L.; White, P. J.; Garrott, R. A.; Houston, D. B. 2007. A seventy-year history of trends in Yellowstone's northern elk herd. The Journal of Wildlife Management. 71(2): 594-602. [83765]
185. Eddleman, Lee E.; Miller, Patricia M.; Miller, Richard F.; Dysart, Patricia L. 1994. Western juniper woodlands (of the Pacific Northwest): Science assessment. Walla Walla, WA: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, Interior Columbia Basin Ecosystem Management Project. 131 p. [27969]
186. Ellison, Lincoln. 1960. Influence of grazing on plant succession of rangelands. Botanical Review. 26(1): 1-78. [862]
187. Ellison, Lincoln; Woolfolk, E. J. 1937. Effects of drought on vegetation near Miles City, Montana. Ecology. 18(3): 329-336. [6264]
188. Ellsworth, Lisa M.; Kauffman, J. Boone. 2010. Native bunchgrass response to prescribed fire in ungrazed mountain big sagebrush ecosystems. Fire Ecology. 6(3): 86-86. [81890]
189. Elmore, Wayne. 1984. Juniper manipulation and the implications on wildlife habitat. In: Proceedings--western juniper management short course; 1984 October 15-16; Bend, OR. Corvallis, OR: Oregon State University, Extension Service and Department of Rangeland Resources: 48-52. [863]
190. Emmerich, F. L.; Tipton, F. H.; Young, J. A. 1993. Cheatgrass: Changing perspectives and management strategies. Rangelands. 15(1): 37-40. [20463]
191. Eng, Robert L.; Schladweiler, P. 1972. Sage grouse winter movements and habitat use in central Montana. The Journal of Wildlife Management. 36: 141-146. [7529]
192. Erikson, Heidi Jo. 2011. Herbaceous and avifauna responses to prescribed fire and grazing timing in a high-elevation sagebrush ecosystem. Fort Collins, CO: Colorado State University. 165 p. Thesis. [90063]
193. Evans, Raymond A.; Young, James A. 1986. Germination profiles for five populations of big sagebrush. In: McArthur, E. Durant; Welch, Bruce L., compilers. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 366-369. [883]
194. Everett, Richard L. 1980. Use of containerized shrubs for revegetating arid roadcuts. Reclamation Review. 3: 33-40. [11003]
195. Evers, Louisa B.; Miller, Richard F.; Doescher, Paul S.; Hemstrom, Miles; Neilson, Ronald P. 2013. Stimulating current successional trajectories in sagebrush ecosystems with multiple disturbances using a state-and-transition modeling framework. Rangeland Ecology & Management. 66(3): 313-329. [87377]
196. Evers, Louisa. 2010. Modeling sage-grouse habitat using a state-and-transition model. Corvallis, OR: Oregon State University. 162 p. [+ appendices]. Dissertation. [90037]
197. Ewers, B. E.; Pendall, E. 2008. Spatial patterns in leaf area and plant functional type cover across chronosequences of sagebrush ecosystems. Plant Ecology. 194(1): 67-83. [91476]
198. Fagerstone, Kathleen A.; Lavoie, G. Keith; Griffith, Richard E., Jr. 1980. Black-tailed jackrabbit diet and density on rangeland and near agricultural crops. Journal of Range Management. 33(3): 229-233. [21756]
199. Fairchild, John A. 1999. Pinyon-juniper chaining design guidelines for big game winter range enhancement projects. In: Monsen, Stephen B.; Stevens, Richard, compilers. Proceedings: Ecology and management of pinyon-juniper communities within the Interior West: Sustaining and restoring a diverse ecosystem; 1997 September 15-18; Provo, UT. Proceedings RMRS-P-9. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 278-280. [30565]
200. Fatehi, Mohammad; Pieper, Rex D.; Beck, Reldon E. 1988. Seasonal food habits of blacktailed jackrabbits (Lepus californicus) in southern New Mexico. The Southwestern Naturalist. 33(3): 367-370. [6156]
201. Fechner, Gilbert H.; Barrows, Jack S. 1976. Aspen stands as wildfire fuel breaks. Eisenhower Consortium Bulletin 4. Fort Collins, CO: Colorado State University, College of Forestry. In cooperation with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station; Eisenhower Consortium for Western Environmental Forestry Research. 26 p. [7611]
202. Finch, Deborah M.; Smith, D. Max; LeDee, Olivia; Cartron, Jean-Luc E.; Rumble, Mark A. 2012. Climate change, animal species, and habitats: Adaptation and issues. In: Finch, Deborah M., ed. Climate change in grasslands, shrublands, and deserts of the interior American West: A review and needs assessment. Gen. Tech. Rep. RMRS-GTR-285. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 1-20. [91462]
203. Fischer, Richard A.; Reese, Kerry P.; Connelly, John W. 1996. An investigation on fire effects within xeric sage grouse brood habitat. Journal of Range Management. 49(3): 194-198. [26598]
204. Flora of North America Editorial Committee, eds. 2018. Flora of North America north of Mexico, [Online]. Flora of North America Association (Producer). Available: http://www.efloras.org/flora_page.aspx?flora_id=1. [36990]
205. Forbis, Tara A.; Provencher, Louis; Turner, Lee; Medlyn, Gary; Thompson, Julie; Jones, Gina. 2007. A method for landscape-scale vegetation assessment: Application to Great Basin rangeland ecosystems. Rangeland Ecology & Management. 60(3): 209-217. [91220]
206. Fraas, W. Wyatt; Wambolt, Carl L.; Frisina, Michael R. 1992. Prescribed fire effects on a bitterbrush--mountain big sagebrush--bluebunch wheatgrass community. In: Clary, Warren P.; McArthur, E. Durant; Bedunah, Don; Wambolt, Carl L., comps. Proceedings--symposium on ecology and management of riparian shrub communities; 1991 May 29-31; Sun Valley, ID. Gen. Tech. Rep. INT-289. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 212-216. [19124]
207. Frank, Carolyn T.; Smith, Bruce N.; Welch, Bruce L. 1986. Photosynthesis, growth, transpiration, and 13C relationships among three subspecies of big sagebrush (Artemisia tridentata Nutt.). In: McArthur, E. Durant; Welch, Bruce L., compilers. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT: U.S. Dep. Agric., For. Serv., Intrmtn. Res. Stn.: 332-335. [960]
208. Franklin, Jerry F.; Dyrness, C. T. 1973. Natural vegetation of Oregon and Washington. Corvallis, OR: Oregon State University Press. 452 p. [961]
209. Freeman, D. C.; Turner, W. A.; McArthur, E. D.; Graham, J. H. 1991. Characterization of a narrow hybrid zone between two subspecies of big sagebrush (Artemisia tridentata: Asteraceae). American Journal of Botany. 78(6): 805-815. [15470]
210. Freeman, D. Carl; Graham, John H.; Byrd, David W.; McArthur, E. Durant; Turner, William A. 1995. Narrow hybrid zone between two subspecies of big sagebrush, Artemisia tridentata (Asteraceae). III. Developmental instability. American Journal of Botany. 82(9): 1144-1152. [26180]
211. Freeman, D. Carl; Wang, Han; Sanderson, Stewart; McArthur, E. Durant. 1999. Characterization of a narrow hybrid zone between two subspecies of big sagebrush (Artemisia tridentata, Asteraceae): VII. Community and demographic analyses. Evolutionary Ecology Research. 1: 487-502. [91018]
212. Friggens, Megan M.; Warwell, Marcus V.; Chambers, Jeanne C.; Kitchen, Stanley G. 2012. Modeling and predicting vegetation response of western USA grasslands, shrublands, and deserts to climate change. In: Finch, Deborah M., ed. Climate change in grasslands, shrublands, and deserts of the interior American West: A review and needs assessment. Gen. Tech. Rep. RMRS-GTR-285. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 1-20. [90151]
213. Frischknecht, Neil C. 1962. Factors influencing brush invasion of crested wheatgrass range. Bulletin of the Ecological Society of America. 43(3): 53. [3832]
214. Frischknecht, Neil C.; Baker, Maurice F. 1972. Voles can improve sagebrush rangelands. Journal of Range Management. 25(6): 466-468. [91453]
215. Frischknecht, Neil C.; Harris, Lorin E. 1973. Sheep can control sagebrush on seeded range if... Utah Science. 34(1): 27-30. [91610]
216. Frischknecht, Niel C.; Harris, Lorin E.; Woodward, Harry K. 1953. Cattle gains and vegetal changes as influences by grazing treatments in crested wheatgrass. Journal of Range Management. 6(3): 151-158. [91231]
217. Frisina, Michael R.; Wambolt, Carl L. 2004. Keying in on big sagebrush. Rangelands. 26(1): 12-16. [47232]
218. Gallant, Alisa L.; Hansen, Andrew J.; Councilman, John S.; Monte, Duane K.; Betz, David W. 2003. Vegetation dynamics under fire exclusion and logging in a Rocky Mountain watershed, 1856-1996. Ecological Applications. 13(2): 385-403. [44584]
219. Ganskopp, Dave; Bohnert, Dave; Johnson, Dustin; Munday, Kristen. 2009. Pre- and post-burn cattle distribution patterns in sagebrush steppe. In: Bohnert, Dave; Boyd, Chad; Clark, Pat; Cooke, Reinaldo; [and others]. Range field day 2009 progress report. Special report 1092. Corvalis, OR: Department of Oregon State University Department of Rangeland Ecology and Management; Burns, OR: USDA, Agricultural Research Service, Eastern Oregon Agricultural Research Center, U.S. Department of Agriculture: 25-38. [88677]
220. Garrison, Heather D.; Shultz, Leila M.; McArthur, E. Durant. 2013. Studies of a new hybrid taxon in the Artemisia tridentata (Asteraceae: Anthemideae) complex. Western North American Naturalist. 73(1): 1-19. [91014]
221. Gates, Dillard H. 1964. Sagebrush infested by leaf defoliating moth. Journal of Range Management. 17: 209-210. [91452]
222. Gates, Robert J.; Eng, Robert L. 1984. Sage grouse, pronghorn, and lagomorph use of a sagebrush-grassland burn site on the Idaho National Engineering Laboratory. In: Markham, O. Doyle, ed. Idaho National Engineering Laboratory radio ecology and ecology programs: 1983 progress reports. Idaho Falls, ID: U.S. Department of Energy, Radiological and Environmental Sciences Laboratory: 220-235. [1005]
223. Gates, Robert John. 1983. Sage grouse, lagomorph, and pronghorn use of a sagebrush grassland burn site on the Idaho National Engineering Laboratory. Bozeman, MT: Montana State University. 125 p. [+ appendices]. Thesis. [90177]
224. Geist, Valerius. 1981. Behavior: Adaptive strategies in mule deer. In: Wallmo, Olof C., ed. 1981. Mule and black-tailed deer of North America. Lincoln, NE: University of Nebraska Press: 157-224. [84942]
225. Gentilcore, Dominic M. 2015. Response of pinyon-juniper woodlands to fire, chaining, and hand thinning. Reno, NV: University of Nevada. 109 p. Thesis. [90223]
226. Gifford, Gerald F. 1981. Impact of burning pinyon-juniper debris on select soil properties. Journal of Range Management. 34(5): 357-359. [1016]
227. Goodrich, Sherel. 2005. Classification and capabilities of woody sagebrush communities of western North America with emphasis on sage-grouse habitat. In: Shaw, Nancy L.; Pellant, Mike; Monsen, Stephen B., eds. Sage-grouse habitat restoration symposium proceedings; 2001 June 4-7; Boise, ID. Proc. RMRS-P-38. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 17-37. [63179]
228. Goodrich, Sherel. 2008. Trout Creek 1999 burn. In: Kitchen, Stanley G.; Pendleton, Rosemary L.; Monaco, Thomas A.; Vernon, Jason, comps. Proceedings--shrublands under fire: Disturbance and recovery in a changing world; 2006 June 6-8; Cedar City, UT. Proc. RMRS-P-52. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 147-150. [73289]
229. Goodrich, Sherel; Huber, Allen. 2001. Mountain big sagebrush communities on the Bishop Conglomerate in the eastern Uinta Mountains. In: McArthur, E. Durant; Fairbanks, Daniel J., compilers. Shrubland ecosystem genetics and biodiversity: Proceedings; 2000 June 13-15; Provo, UT. Proc. RMRS-P-21. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 336-343. [41998]
230. Goodrich, Sherel; Huber, Allen; Monroe, Brian. 2008. Trend of mountain big sagebrush crown cover and ground cover on burned sites, Uinta Mountains and West Tavaputs Plateau, Utah. In: Kitchen, Stanley G.; Pendleton, Rosemary L.; Monaco, Thomas A.; Vernon, Jason, comps. Proceedings--shrublands under fire: Disturbance and recovery in a changing world; 2006 June 6-8; Cedar City, UT. Proc. RMRS-P-52. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 151-160. [73290]
231. Goodrich, Sherel; McArthur, E. Durant; Winward, Alma H. 1985. A new combination and a new variety in Artemisia tridentata. The Great Basin Naturalist. 45(1): 99-104. [1034]
232. Goodrich, Sherel; McArthur, E. Durant; Winward, Alma H. 1999. Sagebrush ecotones and average annual precipitation. In: McArthur, E. Durant; Ostler, W. Kent; Wambolt, Carl L., comps. Proceedings: Shrubland ecotones; 1998 August 12-14; Ephraim, UT. Proceedings RMRS-P-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 88-94. [36068]
233. Goodwin, Duwayne Leroy. 1956. Autecological studies of Artemisia tridentata, Nutt. Pullman, WA: State College of Washington. 79 p. Dissertation. [1035]
234. Goodwin, Kim; Sheley, Roger; Clark, Janet. 2002. Integrated noxious weed management after wildfires. EB-160. Bozeman, MT: Montana State University, Extension Service (Producer). 46 p. Available online: http://www.msuextension.org/store/Products/Integrated-Noxious-Weed-Management-After-Wildfires__EB0160.aspx [2011, January 20]. [45303]
235. Graham, John H.; Freeman, D. Carl; McArthur, E. Durant. 1995. Narrow hybrid zone between two subspecies of big sagebrush (Artemisia tridentata: Asteraceae). II. Selection gradients and hybrid fitness. American Journal of Botany. 82(6): 709-716. [26072]
236. Great Plains Flora Association. 1986. Flora of the Great Plains. Lawrence, KS: University Press of Kansas. 1392 p. [1603]
237. Green, Jeffery, S.; Flinders, Jerran T. 1980. Habitat and dietary relationships of the pygmy rabbit. Journal of Range Management. 33(2): 136-142. [6257]
238. Green, Jeffrey S.; Flinders, Jerran T. 1980. Brachylagus idahoensis. Mammalian Species. 125: 1-4. [23631]
239. Griffin, Duane. 2002. Prehistoric human impacts on fire regimes and vegetation in the northern Intermountain West. In: Vale, Thomas R., ed. Fire, native peoples, and the natural landscape. Washington: Island Press: 77-100. [88509]
240. Grove, Adam J. 1998. Effects of Douglas fir establishment in southwestern Montana mountain big sagebrush communities. Bozeman, MT: Montana State University, Department of Animal and Range Science. 150 p. Thesis. [46897]
241. Grove, Adam J.; Wambolt, Carl L.; Frisina, Michael R. 2005. Douglas-fir's effect on mountain big sagebrush wildlife habitats. Wildlife Society Bulletin. 33(1): 74-80. [91221]
242. Gruell, George E. 1983. Fire and vegetative trends in the northern Rockies: Interpretations from 1871-1982 photographs. Gen. Tech. Rep. INT-158. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 117 p. [5280]
243. Gruell, George E. 1985. Fire on the early western landscape: An annotated record of wildland fire. Northwest Science. 59(2): 97-107. [15660]
244. Hall, Harvey M.; Clements, Frederic E. 1923. The phylogenetic method in taxonomy: The North American species of Artemisia, Chrysothamnus, and Atriplex. Publication No. 326. Washington, DC: The Carnegie Institute of Washington. 355 p. [43183]
245. Hann, Wendel J.; Jones, Jeffrey L.; Karl, Michael G. "Sherm"; Hessburg, Paul F.; Keane, Robert E.; Long, Donald G.; Menakis, James P.; McNicoll, Cecilia H.; Leonard, Stephen G.; Gravenmier, Rebecca A.; Smith, Bradley G. 1997. Landscape dynamics of the Basin. In: Quigley, Thomas M.; Arbelbide, Sylvia, tech. eds. An assessment of ecosystem components in the Interior Columbia Basin and portions of the Klamath and Great Basins. Gen. Tech. Rep. PNW-GTR-405. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 337-968 [+ appendices]. [89980]
246. Hanna, Sara K.; Fulgham, Kenneth O. 2015. Post-fire vegetation dynamics of a sagebrush steppe community change significantly over time. California Agriculture. 69(1): 36-42. [90764]
247. Hanson, Clayton L.; Johnson, Clifton W.; Wight, J. Ross. 1982. Foliage mortality of mountain big sagebrush (Artemisia tridentata subsp. vaseyana) in southwestern Idaho during the winter of 1976-77. Journal of Range Management. 35(2): 142-145. [1080]
248. Hardegree, Stuart P. 1994. Matric priming increases germination rate of Great Basin native perennial grasses. Agronomy Journal. 86(2): 289-293. [41619]
249. Hargis, Christina; McCarthy, Clinton. 1986. Vegetation changes following a prescribed burn on a Great Basin meadow. Transactions, Western Section of the Wildlife Society. 2(22): 47-51. [15955]
250. Harniss Roy O.; McDonough, W. T. 1976. Yearly variation in germination in three subspecies of big sagebrush. Journal of Range Management. 29(2): 167-168. [1084]
251. Harniss, R. O.; McDonough, W. T. 1975. Seedling growth of three big sagebrush subspecies under controlled temperature regimens. Journal of Range Management. 28(3): 243-244. [91467]
252. Harrell, Destin. 2016. Personal communication [Phone call with Robin Innes]. 19 May. Regarding use of prescribed fire in greater sage-grouse habitats within the Cody field office. Cody, WY: U.S. Department of the Interior, Bureau of Land Management, Wind River/Bighorn Basin District, Cody Field Office. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [90480]
253. Harte, John; Shaw, Rebecca. 1995. Shifting dominance within a montane vegetation community: Results of a climate-warming experiment. Science. 267(5199): 876-880. [91460]
254. Harvey, S.; Weaver, T. 1979. Vegetative reproduction in twenty-one native shrubs in Montana. Proceedings, Montana Academy of Sciences. 38: 73-77. [47071]
255. Harvey, Stephen John. 1981. Life history and reproductive strategies in Artemisia. Bozeman, MT: Montana State University. 132 p. Thesis. [1102]
256. Hassan, M. A.; West, N. E. 1986. Dynamics of soil seed pools in burned and unburned sagebrush semi-deserts. Ecology. 67(1): 269-272. [1103]
257. Hedlund, J. D.; Rickard, W. H. 1981. Wildfire and the short-term response of small mammals inhabiting a sagebrush-bunchgrass community. Murrelet. 62: 10-14. [1114]
258. Hemstrom, Miles A.; Wisdom, Michael J.; Hann, Wendel J.; Rowland, Mary M.; Wales, Barbara C.; Gravenmier, Rebecca A. 2002. Sagebrush-steppe vegetation dynamics and restoration potential in the interior Columbia Basin, U.S.A. Conservation Biology. 16(5): 1243-1255. [45072]
259. Hess, Jennifer E. 2011. Greater sage-grouse (Centrocercus urophasianus) habitat response to mowing and prescribed burning Wyoming big sagebrush and the influence of disturbance factors on lek persistence in the Bighorn Basin, Wyoming. Laramie, WY: University of Wyoming. 141 p. [+ tables & figures]. Thesis. [90178]
260. Heyerdahl, Emily K.; Miller, Richard F.; Parsons, Russell A. 2006. History of fire and Douglas-fir establishment in a savanna and sagebrush-grassland mosaic, southwestern Montana, USA. Forest Ecology and Management. 230: 107-118. [62651]
261. Hickman, James C., ed. 1993. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 1400 p. [21992]
262. Higgins, Kenneth F.; Kruse, Arnold D.; Piehl, James L. 1989. Effects of fire in the Northern Great Plains. Ext. Circ. EC-761. Brookings, SD: South Dakota State University, Cooperative Extension Service; South Dakota Cooperative Fish and Wildlife Research Unit. 47 p. [14749]
263. Hironaka, M.; Fosberg, M. A.; Winward, A. H. 1983. Sagebrush-grass habitat types of southern Idaho. Bulletin Number 35. Moscow, ID: University of Idaho, Forest, Wildlife and Range Experiment Station. 44 p. [1152]
264. Hironaka, Minoru. 1979. Basic synecological relationships of the Columbia River sagebrush type. In: The sagebrush ecosystem: A symposium: Proceedings; 1978 April; Logan, UT. Logan, UT: Utah State University, College of Natural Resources: 27-32. [1155]
265. Hobbs, N. Thompson; Schimel, David S. 1984. Fire effects on nitrogen mineralization and fixation in mountain shrub and grassland communities. Journal of Range Management. 37(5): 402-405. [91177]
266. Hodgkinson, Harmon S. 1989. Big sagebrush subspecies and management implications. Rangelands. 11(1): 20-22. [6265]
267. Hoffman, Trista Lynn. 1996. An ecological investigation of mountain big sagebrush in the Gardiner Basin. Bozeman, MT: Montana State University. 84 p. Thesis. [46911]
268. Holechek, Jerry L. 1981. Brush control impacts on rangeland wildlife. Journal of Soil and Water Conservation. 36(5): 265-269. [1182]
269. Holmes, Aaron L. 2007. Short-term effects of a prescribed burn on songbirds and vegetation in mountain big sagebrush. Western North American Naturalist. 67(2): 292-298. [67950]
270. Holmes, Aaron L. 2010. Small mammal and bird abundance in relation to post-fire habitat succession in mountain big sagebrush (Artemisia tridentata spp. vaseyana) communities. Corvallis, OR: Oregon State University. 128 p. [+ appendices]. Dissertation. [91212]
271. Holmes, Aaron L.; Maestas, Jeremy D.; Naugle, David E. 2017. Bird response to removal of western juniper in sagebrush-steppe. Rangeland Ecology and Management. 70(1): 87-94. [91612]
272. Holmes, Aaron L.; Robinson, W. Douglas. 2013. Fire mediated patterns of population densities in mountain big sagebrush bird communities. The Journal of Wildlife Management. 77(4): 737-748. [86949]
273. Holmes, Aaron L.; Robinson, W. Douglas. 2016. Small mammal abundance in mountain big sagebrush communities after fire and vegetation recovery. Western North American Naturalist. 76(3): 326-338. [91596]
274. Holmes, Jennifer A.; Johnson, Matthew J. 2005. Brewer's sparrow (Spizella breweri): A technical conservation assessment, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Region (Producer). 51 p. [+ appendices]. Available: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5182053.pdf [2017, February 10]. [91428]
275. Holmes, Jennifer A.; Johnson, Matthew J. 2005. Sage sparrow (Amphispiza belli): A technical conservation assessment, [Online]. Durango, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Region (Producer). Available: http://www.fs.usda.gov/r2/projects/scp/assessments/sagesparrow.pdf [2017, February 1]. [91429]
276. Hooven, Edward F. 1973. Effects of vegetational changes on small forest mammals. In: Hermann, Richard K.; Lavender, Denis P., eds. Even-age management: Proceedings of a symposium; 1972 August 1; [Corvallis, OR]. Paper 848. Corvallis, OR: Oregon State University, School of Forestry: 75-97. [16241]
277. Hopkins, William E. 1979. Plant associations of South Chiloquin and Klamath Ranger Districts--Winema National Forest. R6-Ecol-79-005. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 96 p. [7339]
278. Horner, Michael A. 2001. Vascular flora of the Glass Mountain region, Mono County, California. Aliso. 20(2): 75-105. [53374]
279. Horton, Jerome S.; Campbell, C. J. 1974. Management of phreatophyte and riparian vegetation for maximum multiple use values. Res. Pap. RM-117. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 23 p. [6318]
280. Houston, Douglas B. 1982. The vegetation: Statistics and dynamics. In: The Northern Yellowstone elk: Ecology and management. New York, NY: Macmillan Publishing Co., Inc: 85-136. [91943]
281. Huber, Allen; Goodrich, Sherel; Anderson, Kim. 1999. Diversity with successional status in the pinyon-juniper/mountain mahogany/bluebunch wheatgrass community type near Dutch John, Utah. In: Monsen, Stephen B.; Stevens, Richard, compilers. Proceedings: Ecology and management of pinyon-juniper communities within the Interior West: Sustaining and restoring a diverse ecosystem; 1997 September 15-18; Provo, UT. Proceedings RMRS-P-9. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 114-117. [30544]
282. Huerta, Mariana A.; Whitlock, Cathy; Yale, Jason. 2009. Holocene vegetation-fire-climate linkages in northern Yellowstone National Park, USA. Palaeogeography, Palaeoclimatology, Palaeoecology. 271(1-2): 170-181. [91552]
283. Hulbert, Lloyd C. 1955. Ecological studies of Bromus tectorum and other annual bromegrasses. Ecological Monographs. 25(2): 181-213. [1205]
284. Hull, A. C., Jr.; Hansen, W. Theron, Jr. 1974. Delayed germination of cheatgrass seed. Journal of Range Management. 27(5): 366-368. [43176]
285. Hull, A. C., Jr.; Hull, Mary Kay. 1974. Presettlement vegetation in Cache Valley, Utah and Idaho. Journal of Range Management. 27(1): 27-29. [91604]
286. Hull, A. C., Jr.; Pechanec, Joseph F. 1947. Cheatgrass--a challenge to range research. Journal of Forestry. 45(8): 555-564. [9930]
287. Humphrey, L. David. 1984. Patterns and mechanisms of plant succession after fire on Artemisia-grass sites in southeastern Idaho. Vegetatio. 57: 91-101. [1214]
288. Hungerford, R. D.; Harrington, M. G.; Frandsen, W. H.; Ryan, K. C.; Niehoff, G. J.. 1991. Influence of fire on factors that affect site productivity. In: Harvey, Alan E.; Neuenschwander, Leon F., compilers. Proceedings--management and productivity of western-montane forest soils; 1990 April 10-April 12; Boise. Gen. Tech. Rep. INT-280. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 32-50. [17438]
289. Jacobs, Jim; Scianna, Joseph D.; Winslow, Susan R. 2011. Big sagebrush establishment. Plant Materials Technical Note, No. MT-68. Bozeman, MT: U.S. Department of Agriculture, Natural Resources Conservation Service. 9 p. [91211]
290. Jacobson, Tracy L. C.; Welch, Bruce L. 1987. Planting depth of Hobble Creek Mountain big sagebrush seed. The Great Basin Naturalist. 47(3): 497-499. [3027]
291. Jain, Theresa B.; Battaglia, Mike A.; Han, Han-Sup; Graham, Russell T.; Keyes, Christopher R.; Fried, Jeremy S.; Sandquist, Jonathan E. 2012. A comprehensive guide to fuel management practices for dry mixed conifer forests in the northwestern United States. Gen. Tech. Rep. RMRS-GTR-292. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 331 p. [87811]
292. Jankovsky-Jones, Mabel; Rust, Steven K.; Moseley, Robert K. 1999. Riparian reference areas in Idaho: A catalog of plant associations and conservation sites. Gen. Tech. Rep. RMRS-GTR-20. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 141 p. [29900]
293. Jensen, M. E.; Simonson, G. H.; Dosskey, M. 1990. Correlation between soils and sagebrush-dominated plant communities of northeastern Nevada. Soil Science Society of America Journal. 54: 902-910. [15502]
294. Jensen, Mark E. 1989. Soil characteristics of mountainous northeastern Nevada sagebrush community types. The Great Basin Naturalist. 49(4): 469-481. [9903]
295. Jensen, Mark E. 1989. Soil climate and plant community relationships on some rangelands of northeastern Nevada. Journal of Range Management. 42(4): 275-280. [7976]
296. Jensen, Mark E. 1990. Interpretation of environmental gradients which influence sagebrush community distribution in northeastern Nevada. Journal of Range Management. 43(2): 161-167. [38660]
297. Jensen, Mark E.; Simonson, G. H.; Keane, R. E. 1989. Soil temperature and moisture regime relationships within some rangelands of the Great Basin. Soil Science. 147(2): 134-139. [38661]
298. Johnson, Charles G., Jr.; Clausnitzer, Roderick R.; Mehringer, Peter J.; Oliver, Chadwick D. 1994. Biotic and abiotic processes of eastside ecosystems: The effects of management on plant and community ecology, and on stand and landscape vegetation dynamics. Gen. Tech. Rep. PNW-GTR-322. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 66 p. [23002]
299. Johnson, Charles Grier, Jr. 1998. Vegetation response after wildfires in national forests of northeastern Oregon. R6-NR-ECOL-TP-06-98. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 128 p. [+ appendices]. [30061]
300. Johnson, Charles Grier, Jr.; Swanson, David K. 2005. Bunchgrass plant communities of the Blue and Ochoco Mountains: A guide for managers. Gen. Tech. Rep. PNW-GTR-641. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 119 p. [63805]
301. Johnson, Donald E. 1951. Biology of the elk calf, Cervus canadensis Nelsoni. The Journal of Wildlife Managment. 15(4): 396-410. [83301]
302. Johnson, Douglas H.; Holloran, Matthew J.; Connelly, John W.; Hanser, Steven E.; Amundson, Coutney L.; Knick, Steven T. 2011. Influences of environmental and anthropogenic features on greater sage-grouse populations, 1997-2007. Chapter 17. In: Knick, Steven T.; Connelly, John W., eds. Greater sage-grouse: Ecology and conservation of a landscape species and its habitats. Studies in Avian Biology, No. 38. Berkeley, CA: University of California Press: 407-450. [89723]
303. Johnson, Dustin D.; Miller, Richard F. 2006. Structure and development of expanding western juniper woodlands as influenced by two topographic variables. Forest Ecology and Management. 229(1-3): 7-15. [62648]
304. Johnson, Kendall L. 1979. Basic synecological relationships of the sagebrush types on the high plains of Montana, Wyoming and the Dakotas. In: The sagebrush ecosystem: A symposium: Proceedings; 1978 April; Logan, UT. Logan, UT: Utah State University, College of Natural Resources: 42-49. [1281]
305. Johnson, Kendall L. 1986. Sagebrush over time: A photographic study of rangeland change. In: McArthur, E. Durant; Welch, Bruce L., comps. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 223-252. [1282]
306. Johnston, Barry C. 1987. Plant associations of Region 2: Potential plant communities of Wyoming, South Dakota, Nebraska, Colorado, and Kansas. 4th ed. R2-ECOL-87-2. Lakewood, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Region. 429 p. [54304]
307. Kachergis, Emily; Fernandez-Gimenez, Maria E.; Rocca, Monique E. 2012. Differences in plant species composition as evidence of alternate states in the sagebrush steppe. Rangeland Ecology & Management. 65(5): 486-497. [91027]
308. Kaltenecker, J. H.; Wicklow-Howard, M. C.; Rosentreter, R. 1999. Biological soil crusts in three sagebrush communities recovering from a century of livestock trampling. In: McArthur, E. Durant; Ostler, W. Kent; Wambolt, Carl L., compilers. Proceedings: Shrubland ecotones; 1998 August 12-14; Ephraim, UT. Proceedings RMRS-P-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 222-226. [36091]
309. Karban, Richard; Ishizaki, Satomi; Shiojiri, Kaori. 2012. Long-term demographic consequences of eavesdropping for sagebrush. Journal of Ecology. 100(4): 932-938. [85645]
310. Kartesz, J. T. The Biota of North America Program (BONAP). 2015. Taxonomic Data Center, [Online]. Chapel Hill, NC: The Biota of North America Program (Producer). Available: http://bonap.net/tdc [Maps generated from Kartesz, J. T. 2010. Floristic synthesis of North America, Version 1.0. Biota of North America Program (BONAP). [in press]. [84789]
311. Kartesz, John T. 1994. A synonymized checklist of the vascular flora of the United States, Canada, and Greenland. Volume I--checklist. 2nd ed. Portland, OR: Timber Press. 622 p. [23877]
312. Kasworm, Wayne F.; Irby, Lynn R.; Ihsle Pac, Helga B. 1984. Diets of ungulates using winter ranges in northcentral Montana. Journal of Range Management. 37(1): 67-71. [63610]
313. Keane, Robert E.; Agee, James K.; Fule, Peter; Keeley, Jon E.; Key, Carl; Kitchen, Stanley G.; Miller, Richard; Schulte, Lisa A. 2008. Ecological effects of large fires on US landscapes: Benefit or catastrophe? International Journal of Wildland Fire. 17(6): 696-712. [73387]
314. Keane, Robert E.; Gray, Kathy; Bacciu, Valentina. 2012. Spatial variability of wildland fuel characteristics in northern Rocky Mountain ecosystems. Res. Pap. RMRS-RP-98. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 56 p. [88951]
315. Keating, Kimberly A.; Irby, Lynn R.; Kasworm, Wayne F. 1985. Mountain sheep winter food habits in the upper Yellowstone Valley. The Journal of Wildlife Management. 49(1): 156-161. [15521]
316. Kelsey, Rick G.; Wright, William E.; Sneva, Forrest; Winward, Al; Britton, Carlton. 1983. The concentration and composition of big sagebrush essential oils from Oregon. Biochemical Systematics and Ecology. 11(4): 353-360. [91447]
317. Kerley, Linda L.; Anderson, Stanley H. 1995. Songbird responses to sagebrush removal in a high elevation sagebrush steppe ecosystem. Prairie Naturalist. 27(3): 129-146. [27097]
318. Kindschy, Robert R.; Sundstrom, Charles; Yoakum, James D. 1982. Wildlife habitats in managed rangelands--the Great Basin of southeastern Oregon: Pronghorns. Gen. Tech. Rep. PNW-145. Portland, OR: U.S. Department of Agriculture, Forest Service. 18 p. [9496]
319. Kitchen, Stanley G.; McArthur, E. Durant. 2007. Big and black sagebrush landscapes. In: Hood, Sharon M.; Miller, Melanie, eds. Fire ecology and management of the major ecosystems of southern Utah. Gen. Tech. Rep. RMRS-GTR-202. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 73-95. [71081]
320. Kitchen, Stanley G.; Weisberg, Peter J. 2013. Historic fire frequency in mountain big sagebrush communities of the eastern Great Basin and Colorado Plateau: A comparison of estimates based upon proxy fire records and predictions derived from post-fire succession rates, [Online]. JFSP Research Project Reports, Paper 92, Project Number 06-3-1-17. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Joint Fire Science Program (Producer). 39 p. Available: http://digitalcommons.unl.edu/jfspresearch/92. [90808]
321. Klebenow, D.; Beall, R; Bruner, A.; Mason, R.; Roundy, B.; Stager, W; Ward, K. 1977. Response of wildlife populations to prescribed burning in pinyon-juniper woodlands. In: Controlled fire as a management tool in the pinyon-juniper woodland, Nevada. Summary Progress Report FY 1977. Reno, NV: University of Nevada, Nevada Agricultural Experiment Station: 86-103. [25710]
322. Klebenow, Donald A. 1973. The habitat requirements of sage grouse and the role of fire in management. In: Proceedings, annual Tall Timbers fire ecology conference; 1972 June 8-9; Lubbock, TX. No. 12. Tallahassee, FL: Tall Timbers Research Station: 305-315. [1345]
323. Klebenow, Donald A. 1985. Big game response to fire in sagebrush-grass rangelands. In: Saunders, Ken; Durham, Jack; [and others], eds. Rangeland fire effects: Proceedings of the symposium; 1984 November 27-29; Boise, ID. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office: 53-57. [1347]
324. Klott, James H.; Ketchum, Chris. 1991. The results of using "Hobble Creek" sagebrush on two fire rehabilitations. Idaho BLM Technical Bulletin 91-1. Boise, ID: U.S. Department of the Interior, Bureau of Land Management. 12 p. [19869]
325. Knapp, Paul A. 1998. Spatio-temporal patterns of large grassland fires in the Intermountain West, U.S.A. Global Ecology and Biogeography Letters. 7(4): 259-273. [30109]
326. Knick, Steven T.; Dobkin, David S.; Rotenberry, John T.; Schroeder, Michael A.; Vander Haegen, W. Matthew; van Riper, Charles, III. 2003. Teetering on the edge or too late? Conservation issues for avifauna of sagebrush habitats. The Condor. 105(4): 611-634. [75524]
327. Knick, Steven T.; Holmes, Aaron L.; Miller, Richard F. 2005. The role of fire in structuring sagebrush habitats and bird communities. In: Saab, Victoria A.; Powell, Hugh D. W., eds. Fire and avian ecology in North America. Studies in Avian Biology No. 30. Ephrata, PA: Cooper Ornithological Society: 63-75. [65140]
328. Knick, Steven T.; Rotenberry, John T. 1997. Landscape characteristics of disturbed shrubsteppe habitats in southwestern Idaho (U.S.A.). Landscape Ecology. 12(5): 287-297. [43168]
329. Koniak, Susan. 1985. Succession in pinyon-juniper woodlands following wildfire in the Great Basin. The Great Basin Naturalist. 45(3): 556-566. [1371]
330. Korfmacher, John L.; Chambers, Jeanne C.; Tausch, Robin J.; Roundy, Bruce A.; Meyer, Susan E.; Kitchen, Stanley. 2003. Technical Note: A technique for conducting small-plot burn treatments. Journal of Range Management. 56(3): 251-254. [47097]
331. Kufeld, Roland C. 1973. Foods eaten by the Rocky Mountain elk. Journal of Range Management. 26(2): 106-113. [1385]
332. Kufeld, Roland C.; Wallmo, O. C.; Feddema, Charles. 1973. Foods of the Rocky Mountain mule deer. Res. Pap. RM-111. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 31 p. [1387]
333. Kuntz, David Edward. 1982. Plant response following spring burning in an Artemisia tridentata subsp. vaseyana/ Festuca idahoensis habitat type. Moscow, ID: University of Idaho. 73 p. Thesis. [1388]
334. La Tourrette, Jospeh E.; Young, James A.; Evans, Raymond A. 1971. Seed dispersal in relation to rodent activities in seral big sagebrush communities. Journal of Range Management. 24(2): 118-120. [90620]
335. Lackschewitz, Klaus. 1991. Vascular plants of west-central Montana--identification guidebook. Gen. Tech. Rep. INT-227. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 648 p. [13798]
336. Lamanna, Christine A. 2012. The structure and function of subalpine ecosystems in the face of climate change. Tucson, AZ: The University of Arizona. 171 p. [+ tables & figures]. Dissertation. [90829]
337. Lambert, Scott M. 2005. Seeding considerations in restoring big sagebrush habitat. In: Shaw, Nancy L.; Pellant, Mike; Monsen, Stephen B., eds. Sage-grouse habitat restoration symposium proceedings; 2001 June 4-7; Boise, ID. Proc. RMRS-P-38. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 75-80. [63186]
338. Lambrecht, Susan C.; Loik, Michael E.; Inouye, David W.; Harte, John. 2007. Reproductive and physiological responses to simulated climate warming for four subalpine species. New Phytologist. 173(1): 121-134. [65271]
339. Landeen, Melissa L. 2015. Mountain big sagebrush (Artemisia tridentata ssp vaseyana) seed production. Provo, UT: Brigham Young University. 72 p. Thesis. [91499]
340. LANDFIRE. 2008. Alaska refresh (LANDFIRE 1.1.0). Biophysical settings layer. In: LANDFIRE data distribution site, [Online]. In: LANDFIRE. U.S. Department of the Interior, Geological Survey (Producer). Available: https://landfire.cr.usgs.gov/viewer/ [2017, January 10]. [86808]
341. LANDFIRE. 2008. CONUS refresh (LANDFIRE 1.1.0). Biophysical settings layer, LANDFIRE data distribution site, [Online]. In: LANDFIRE. U.S. Department of the Interior, Geological Survey (Producer). Available: https://landfire.cr.usgs.gov/viewer/ [2017, January 10]. [89416]
342. Langenheim, Jean H. 1956. Plant succession on a subalpine earthflow in Colorado. Ecology. 37(2): 301-317. [50443]
343. Larrucea, E. S.; Brussard, P. F. 2008. Shift in location of pygmy rabbit (Brachylagus idahoensis) habitat in response to changing environments. Journal of Arid Environments. 72(9): 1636-1643. [91522]
344. Larrucea, Eveline; S.; Brussard, P. F. 2008. Habitat selection and current distribution of the pygmy rabbit in Nevada and California, USA. Journal of Mammalogy. 89(3): 691-699. [91443]
345. Laycock, W. A. 1991. Stable states and thresholds of range condition on North American rangelands: A viewpoint. Journal of Range Management. 44(5): 427-433. [43349]
346. Laycock, William A. 1979. Management of sagebrush. Rangelands. 1(5): 207-210. [1423]
347. Lesica, Peter. 2017. Personal communication [Email to Robin Innes]. 6 November. Unpublished data and information regarding mountain big sagebrush postfire recovery. Missoula, MT. Documents on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92325]
348. Lesica, Peter; Cooper, Stephen V.; Kudray, Greg. 2005. Big sagebrush shrub-steppe postfire succession in southwest Montana. Report to Bureau of Land Management, Dillon Field Office. Helena, MT: Montana Natural Heritage Program, Natural Resource Information System. 29 p. [+ appendices]. [91595]
349. Lesica, Peter; Cooper, Stephen V.; Kudray, Greg. 2007. Recovery of big sagebrush following fire in southwest Montana. Rangeland Ecology & Management. 60(3): 261-269. [68272]
350. Levitt, Jacob. 1980. 2nd ed. Responses of plants to environmental stresses. Volume I: Chilling, freezing, and high temperature stresses. Physiological Ecology Series. New York: Academic Press. 497 p. [79587]
351. Lewis, Mont E. 1971. Flora and major plant communities of the Ruby-East Humboldt Mountains with special emphasis on Lamoille Canyon. Elko, NV: U.S. Department of Agriculture, Forest Service, Region 4, Humboldt National Forest. 62 p. [1450]
352. Link, Steven O.; Keeler, Carson W.; Hill, Randal W.; Hagan, Eric. 2006. Bromus tectorum cover mapping and fire risk. International Journal of Wildland Fire. 15: 113-119. [91583]
353. Littell, Jeremy S.; McKenzie, Donald; Peterson, David L.; Westerling, Anthony L. 2009. Climate and wildfire area burned in western U.S. ecoprovinces, 1916-2003. Ecological Applications. 19(4): 1003-1021. [81459]
354. Loik, Michael E.; Harte, John. 1996. High-temperature tolerance of Artemisia tridentata and Potentilla gracilis under a climate change manipulation. Oecologia. 108(2): 224-231. [28508]
355. Lommasson, T. 1948. Succession in sagebrush. Journal of Range Management. 1: 19-21. [1468]
356. Long, Lynn E. 1986. Container nursery production of Artemisia and Chrysothamnus species. In: McArthur, E. Durant; Welch, Bruce L., compilers. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 395-396. [1469]
357. Long, Ryan A.; Rachlow, Janet L.; Kie, John G. 2008. Effects of season and scale on response of elk and mule deer to habitat manipulation. The Journal of Wildlife Management. 72(5): 1133-1142. [82412]
358. Long, Ryan A.; Rachlow, Janet L.; Kie, John G.; Vavra, Martin. 2008. Fuels reduction in a western coniferous forest: Effects on quantity and quality of forage for elk. Rangeland Ecology and Management. 61: 302-313. [70956]
359. Longland, William S. 1994. Seed use by desert granivores. In: Monsen, Stephen B.; Kitchen, Stanley G., compilers. Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 233-237. [24288]
360. Lyon, L. Jack; Crawford, Hewlette S.; Czuhai, Eugene; Fredriksen, Richard L.; Harlow, Richard F.; Metz, Louis J.; Pearson, Henry A. 1978. Effects of fire on fauna: A state-of-knowledge review--National fire effects workshop; 1978 April 10-14; Denver, CO. Gen. Tech. Rep. WO-6. Washington, DC: U.S. Department of Agriculture, Forest Service. 41 p. [25066]
361. Lyon, L. Jack; Telfer, Edmund S.; Schreiner, David Scott. 2000. Direct effects of fire and animal responses. In: Smith, Jane Kapler, ed. Wildland fire in ecosystems: Effects of fire on fauna. Gen. Tech. Rep. RMRS-GTR-42-Vol. 1. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 17-23. [44435]
362. MacDonald, Linda. 1999. Wildfire rehabilitation in Utah. In: Monsen, Stephen B.; Stevens, Richard, compilers. Proceedings: Ecology and management of pinyon-juniper communities within the Interior West: Sustaining and restoring a diverse ecosystem; 1997 September 15-18; Provo, UT. Proceedings RMRS-P-9. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 410-411. [30590]
363. Mackie, Richard J.; Kie, John G.; Pac, David F.; Hamlin, Kenneth L. 2003. Mule deer (Odocoileus hemionus). In: Feldhamer, George A.; Thompson, Bruce C.; Chapman, Joseph A., eds. Wild mammals of North America: Biology, management, and conservation. 2nd ed. Baltimore, MD: Johns Hopkins University Press: 889-905. [82121]
364. Mahalovich, Mary F.; McArthur, E. Durant. 2004 (revised 2016). Sagebrush (Artemisia spp.): Seed and plant transfer guidelines. Native Plants Journal. Fall 2004: 141-148. [90903]
365. Maier, Aaron M.; Perryman, Barry L.; Olson, Richard A.; Hild, Ann L. 2001. Climatic influences on recruitment of 3 subspecies of Artemisia tridentata. Journal of Range Management. 54(6): 699-703. [40189]
366. Mangan, Larry; Autenrieth, R. 1985. Vegetation changes following 2,4-D application and fire in a mountain big sagebrush habitat type. In: Saunders, Ken; Durham, Jack; [and others], eds. Rangeland fire effects: Proceedings of the symposium; 1984 November 27-29; Boise, ID. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office: 61-65. [1519]
367. Manier, D. J.; Wood, D. J. A.; Bowen, Z. H.; Donovan, R. M.; Holloran, M. J.; Juliusson, L. M.; Mayne, K. S.; Oyler-McCance, S. J.; Quamen, F. R.; Saher, D. J.; Titolo, A. J. 2013. Summary of science, activities, programs, and policies that influence the rangewide conservation of greater sage-grouse (Centrocercus urophasianus), [Online]. Open-File Report 2013-1098. Reston, VA: U.S. Department of the Interior, U.S. Geological Survey (Producer). 151 p. [+ supplementary information] Available: http://pubs.usgs.gov/of/2013/1098/ [2016, March 21]. [90148]
368. Manier, Daniel J.; Hobbs, N. Thompson; Theobald, David M.; Reich, Robin M.; Kalkhan, Mohammed A.; Campbell, Mark R. 2005. Canopy dynamics and human caused disturbance on a semi-arid landscape in the Rocky Mountains, USA. Landscape Ecology. 20(1): 1-17. [91631]
369. Manning, Mary E.; Padgett, Wayne G. 1995. Riparian community type classification for Humboldt and Toiyabe National Forests, Nevada and eastern California. R4-Ecol-95-01. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Region. 306 p. [42196]
370. Marlette, Guy M.; Anderson, Jay E. 1986. Seed banks and propagule dispersal in crested-wheatgrass stands. Journal of Applied Ecology. 23: 161-175. [1526]
371. Marlow, Clayton B; Pogacnik, Thomas M; Quinsey, Shannon D. 1987. Streambank stability and cattle grazing in southwestern Montana. Journal of Soil and Water Conservation. 42(4): 291-296. [2888]
372. Martin, Alexander C.; Zim, Herbert S.; Nelson, Arnold L. 1951. American wildlife and plants. New York: McGraw-Hill. 500 p. [4021]
373. Martin, Robert C. 1990. Sage grouse responses to wildfire in spring and summer habitats. Moscow, ID: University of Idaho. 36 p. Thesis. [24907]
374. Maser, Chris; Gashwiler, Jay S. 1978. Interrelationships of wildlife and western juniper. In: Martin, Robert E.; Dealy, J. Edward; Caraher, David L., eds. Proceedings of the western juniper ecology and management workshop; 1977 January; Bend, OR. Gen. Tech. Rep. PNW-74. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: 37-82. [1541]
375. Maser, Chris; Strickler, Gerald S. 1978. The sage vole, Lagurus curtatus, as an inhabitant of subalpine sheep fescue, Festuca ovina, communities on Steens Mountain--an observation and interpretation. Northwest Science. 52(3): 276-284. [15507]
376. Mattise, Samuel N. 1995. Sage grouse in Idaho: Forum '94. Tech. Bull. No. 95-15. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office. 10 p. [26119]
377. McAdoo, J. Kent; Barrington, Mack R.; Ports, Mark A. 2006. Habitat affinities of rodents in northeastern Nevada rangeland communities. Western North American Naturalist. 66(3): 321-331. [65155]
378. McAdoo, J. Kent; Longland, William S.; Cluff, Greg J.; Klebenow, Donald A. 1987. Use of new rangeland seedings by black-tailed jackrabbits. Journal of Range Management. 40(6): 520-524. [135]
379. McArthur, E. D.; Welch, B. L.; Sanderson, S. C. 1988. Natural and artificial hybridization between big sagebrush (Artemisia tridentata) subspecies. Journal of Heredity. 79: 268-276. [76195]
380. McArthur, E. Durant. 1994. Ecology, distribution, and values of sagebrush within the Intermountain region. In: Monsen, Stephen B.; Kitchen, Stanley G., compilers. Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 347-351. [24308]
381. McArthur, E. Durant. 2000. Sagebrush systematics and distribution. In: Entwistle, P. G.; DeBolt, A. M.; Kaltenecker, J. H.; Steenhof, K., compilers. Sagebrush steppe ecosystems symposium: Proceedings; 1999 June 21-23; Boise, ID. Publ. No. BLM/ID/PT-001001+1150. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Boise State Office: 9-14. [41811]
382. McArthur, E. Durant. 2005. Sagebrush, common and uncommon, palatable and unpalatable. Rangelands. 27(4): 47-51. [60405]
383. McArthur, E. Durant; Blauer, A. Clyde; Plummer, A. Perry; Stevens, Richard. 1979. Characteristics and hybridization of important Intermountain shrubs. III. Sunflower family. Res. Pap. INT-220. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 82 p. [1571]
384. McArthur, E. Durant; Blauer, A. Clyde; Sanderson, Stewart C. 1988. Mule deer-induced mortality of mountain big sagebrush. Journal of Range Management. 41(2): 114-116. [3068]
385. McArthur, E. Durant; Mudge, Joann; Van Buren, Renee; Andersen, W. Ralph; Sanderson, Stewart C.; Babbel, David G. 1998. Randomly amplified polymorphic DNA analysis (RAPD) of Artemisia subgenus Tridentatae species and hybrids. The Great Basin Naturalist. 58(1): 12-27. [28609]
386. McArthur, E. Durant; Sanderson, Stewart C. 1999. Cytogeography and chromosome evolution of subgenus Tridentatae of Artemisia (Asteraceae). American Journal of Botany. 86(12): 1754-1775. [34931]
387. McArthur, E. Durant; Sanderson, Stewart C. 1999. Ecotones: Introduction, scale, and big sagebrush example. In: McArthur, E. Durant; Ostler, W. Kent; Wambolt, Carl L., compilers. Proceedings: shrubland ecotones; 1998 August 12-14; Ephraim, UT. Proceedings RMRS-P-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 3-8. [36053]
388. McArthur, E. Durant; Welch, Bruce L. 1982. Growth rate differences among big sagebrush [Artemisia tridentata] accessions and subspecies. Journal of Range Management. 35(3): 396-401. [91233]
389. McDonough, W. T.; Harniss, R. O. 1974. Effects of temperature on germination in three subspecies of big sagebrush. Journal of Range Management. 27(3): 204-205. [1597]
390. McDonough, W. T.; Harniss, R. O. 1974. Seed dormancy in Artemisia tridentata Nutt. subspecies vaseyana Rydb. Northwest Science. 48(1): 17-20. [1598]
391. McDowell, Michelle K. D. 2000. The effects of burning in mountain big sagebrush on key sage grouse habitat characteristics in southeastern Oregon. Corvallis, OR: Oregon State University. 62 p. Thesis. [90041]
392. McGee, John M. 1977. Effects of prescribed burning on a sagebrush ecosystem in northwestern Wyoming. Final report: Cooperative Agreement No. 16-675-CA. Laramie, WY: University of Wyoming. 134 p. [49191]
393. McGee, John M. 1982. Small mammal populations in an unburned and early fire successional sagebrush community. Journal of Range Management. 35(2): 177-180. [1601]
394. McIntyre, Kristian K. 2003. Species composition and beta diversity of avian communities along a burned/unburned gradient in sagebrush steppe habitat at Sheldon National Wildlife Refuge, Nevada. Alpine, TX: Sul Ross State University. 86 p. [+ appendices]. Thesis. [91471]
395. McIver, J. D.; Brunson, M.; Bunting, S.; Chambers, J.; Devoe, N.; Doescher, P.; Grace, J.; Johnson, D.; Knick, S.; Miller, R.; Pellant, M.; Pierson, F.; Pyke, D.; Rollins, K.; Roundy, B.; Schupp, E.; Tausch, R.; Turner, D. 2010. The Sagebrush Steppe Treatment Evaluation Project (SageSTEP): A test of state-and-transition theory. RMRS-GTR-237. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 16 p. [91518]
396. McLean, Alastair. 1970. Plant communities of the Similkameen Valley, British Columbia. Ecological Monographs. 40(4): 403-424. [1620]
397. Medin, Dean E.; Welch, Bruce L.; Clary, Warren P. 2000. Bird habitat relationships along a Great Basin elevational gradient. Res. Pap. RMRS-RP-23. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 22 p. [38470]
398. Mehus, Chris Allen. 1995. Influences of browsing and fire on sagebrush taxa of the northern Yellowstone winter range. Bozeman, MT: Montana State University. 70 p. Thesis. [46920]
399. Menakis, James P.; Osborne, Dianne; Miller, Melanie. 2003. Mapping the cheatgrass-caused departure from historical natural fire regimes in the Great Basin, USA. In: Omi, Philip N.; Joyce, Linda A., tech. eds. Fire, fuel treatments, and ecological restoration: Conference proceedings; 2002 April 16-18; Fort Collins, CO. Proceedings RMRS-P-29. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 281-287. [45333]
400. Messina, Frank J.; Durham, Susan L.; Richards, James H.; McArthur, E. Durant. 2002. Trade-off between plant growth and defense? A comparison of sagebrush populations. Oecologia. 131(1): 43-51. [91234]
401. Meyer, S. E. 1990. Seed source differences in germination under snowpack in northern Utah. In: Proceedings, 5th Billings symposium on disturbed land rehabilitation: Volume I; 1990 March 25-30; Billings, MT. Reclamation Research Unit Publication No. 9003. 184-191. [19829]
402. Meyer, S. E. 2012. Restoring and managing cold desert shrublands for climate change mitigation, Chapter 2. In: Finch, Deborah M., ed. Climate change in grasslands, shrublands, and deserts of the interior American West: A review and needs assessment. Gen. Tech. Rep. RMRS-GTR-285. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 21-34. [91463]
403. Meyer, Susan E. 1994. Germination and establishment ecology of big sagebrush: Implications for community restoration. In: Monsen, Stephen B.; Kitchen, Stanley G., compilers. Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 244-251. [24290]
404. Meyer, Susan E. 2008. Artemisia L., sagebrush. In: Bonner, Franklin T.; Karrfalt, Robert P., eds. Woody plant seed manual. Agric. Handbook No. 727. Washington, DC: U.S. Department of Agriculture, Forest Service: 274-282. [45115]
405. Meyer, Susan E.; Monsen, Stephen B. 1990. Seed-source differences in initial establishment for big sagebrush and rubber rabbitbrush. In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., compilers. Proceedings--symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 200-208. [12852]
406. Meyer, Susan E.; Monsen, Stephen B. 1991. Habitat-correlated variation in mountain big sagebrush (Artemisia tridentata ssp. vaseyana) seed germination patterns. Ecology. 72(2): 739-742. [15484]
407. Meyer, Susan E.; Monsen, Stephen B. 1992. Big sagebrush germination patterns: Subspecies and population differences. Journal of Range Management. 45(1): 87-93. [17776]
408. Meyer, Susan E.; Monsen, Stephen B.; McArthur, E. Durant. 1990. Germination response of Artemisia tridentata (Asteraceae) to light and chill: Patterns of between-population variation. Botanical Gazette. 151(2): 176-183. [15525]
409. Miglia, Kathleen J. 2004. Importance of genotype, soil type, and location on the performance of parental and hybrid big sagebrush reciprocal transplants in the gardens of Salt Creek Canyon. In: Hild, Ann L.; Shaw, Nancy L.; Meyer, Susan E.; Booth, D. Terrance; McArthur, E. Durant, compilers. Seed and soil dynamics in shrubland ecosystems: Proceedings; 2002 August 12-16; Laramie, WY. Proceedings RMRS-P-31. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 30-36. [49082]
410. Miller, R. F.; Bates, J. D.; Svejcar, T. J.; Pierson, F. B.; Eddleman, L. E. 2007. Western juniper field guide: Asking the right questions to select appropriate management actions. U.S. Geological Survey Circular 1321. Reston, VA: U.S. Department of the Interior, U.S. Geological Survey. 61 p. [88757]
411. Miller, R. F.; Doescher P. S.; Swanson, S. R.; Svejcar, T. J.; Buckhouse, J. C.; Sneva, F. A.; Winward, A. H. 1982. Soil-plant relationships among three big sagebrush subspecies. In: Special Report 663. 1982 Progress report--research in rangeland management. Corvallis, OR: Oregon State University, Agricultural Experiment Station: 5-9. In cooperation with: U.S. Department of Agriculture, Agricultural Research Service. [3643]
412. Miller, Richard F. 1989. Plant competition in Oregon's high desert. In: Oregon's high desert: the last 100 years. Special Report 841. Corvallis, OR: Oregon State University, Agricultural Experiment Station: 7-14. In cooperation with: U.S. Department of Agriculture, Agricultural Research Service. [15513]
413. Miller, Richard F.; Bates, Jon D.; Svejcar, Tony J.; Pierson, Fred B.; Eddleman, Lee E. 2005. Biology, ecology, and management of western juniper, [Online]. Tech. Bull. 152. Corvallis, OR: Oregon State University, Agricultural Experiment Station (Producer). 77 p. Available: http://juniper.oregonstate.edu/bibliography/documents/phpQ65pOk_tb152.pdf [2016, April 6]. [64176]
414. Miller, Richard F.; Chambers, Jeanne C.; Pellant, Mike. 2014. A field guide for selecting the most appropriate treatment in sagebrush and pinon-juniper ecosystems in the Great Basin: Evaluating resilience to disturbance and resistance to invasive annual grasses, and predicting vegetation response. RMRS-GTR-322-rev. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 68 p. [90440]
415. Miller, Richard F.; Chambers, Jeanne C.; Pellant, Mike. 2015. A field guide for rapid assessment of post-wildfire recovery potential in sagebrush and pinon-juniper ecosystems in the Great Basin: Evaluating resilience to disturbance and resistance to invasive annual grasses and predicting vegetation response. Gen. Tech. Rep. RMRS-GTR-338. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 70 p. [90443]
416. Miller, Richard F.; Chambers, Jeanne C.; Pyke, David A.; Pierson, Fred B.; Williams, C. Jason. 2013. A review of fire effects on vegetation and soils in the Great Basin Region: Response and ecological site characteristics. Gen. Tech. Rep. RMRS-GTR-308. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 126 p. [87889]
417. Miller, Richard F.; Eddleman, Lee L. 2000. Spatial and temporal changes of sage grouse habitat in the sagebrush biome. Tech. Bull. 151. Corvallis, OR: Oregon State University, Agricultural Experiment Station. 35 p. [40586]
418. Miller, Richard F.; Heyerdahl, Emily K. 2008. Fine-scale variation of historical fire regimes in sagebrush-steppe and juniper woodland: An example from California, USA. International Journal of Wildland Fire. 17: 245-254. [70528]
419. Miller, Richard F.; Knick, Steven T.; Pyke, David A.; Meinke, Cara W.; Hanser, Steven E.; Wisdom, Michael J.; Hild, Ann L. 2011. Characteristics of sagebrush habitats and limitations to long-term conservation. In: Knick, Steven T.; Connelly, John W., eds. Greater sage-grouse: Ecology and conservation of a landscape species and its habitats. Studies in Avian Biology, No. 38. Berkeley, CA: University of California Press: 145-184. [89659]
420. Miller, Richard F.; Ratchford, Jaime; Roundy, Bruce A.; Tausch, Robin J.; Hulet, April; Chambers, Jeanne. 2014. Response of conifer-encroached shrublands in the Great Basin to prescribed fire and mechanical treatments. Rangeland Ecology and Management. 67(5): 468-481. [89200]
421. Miller, Richard F.; Rose, Jeffery A. 1995. Historic expansion of Juniperus occidentalis (western juniper) in southeastern Oregon. The Great Basin Naturalist. 55(1): 37-45. [25666]
422. Miller, Richard F.; Rose, Jeffrey A. 1999. Fire history and western juniper encroachment in sagebrush steppe. Journal of Range Management. 52(6): 550-559. [28671]
423. Miller, Richard F.; Svejcar, Tony J.; Rose, Jeffrey A. 2000. Impacts of western juniper on plant community composition and structure. Journal of Range Management. 53(6): 574-585. [36578]
424. Miller, Richard F.; Svejcar, Tony J.; West, Neil E. 1994. Implications of livestock grazing in the Intermountain sagebrush region: Plant composition. In: Vavra, Martin; Laycock, William A.; Pieper, Rex D., eds. Ecological implications of livestock herbivory in the West. Denver, CO: Society for Range Management: 101-146. [91614]
425. Miller, Richard F.; Tausch, Robin J. 2001. The role of fire in juniper and pinyon woodlands: A descriptive analysis. In: Galley, Krista E. M.; Wilson, Tyrone P., eds. Proceedings of the invasive species workshop: The role of fire in the control and spread of invasive species; Fire conference 2000: The first national congress on fire ecology, prevention, and management; 2000 November 27 - December 1; San Diego, CA. Misc. Publ. No. 11. Tallahassee, FL: Tall Timbers Research Station: 15-30. [40675]
426. Miller, Richard F.; Tausch, Robin J.; McArthur, E. Durant; Johnson, Dustin D.; Sanderson, Stewart C. 2008. Age structure and expansion of pinyon-juniper woodlands: A regional perspective in the Intermountain West. Research Paper RMRS-RP-69. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 15 p. [91332]
427. Miller, Richard F.; Wigand, Peter E. 1994. Holocene changes in semiarid pinyon-juniper woodlands. Bioscience. 44(7): 465-474. [23563]
428. Miller, Rick; Rose, Jeff. 1998. Pre- and post-settlement fire return intervals on Intermountain sagebrush steppe. In: Annual report: Eastern Oregon Agricultural Research Center. Corvallis, OR: Oregon State University, Agricultural Experiment Station: 16-17. [29194]
429. Miller, Rick; Rose, Jeffrey; Svejcar, Tony; Bates, Jon; Paintner, Kara. 1995. Western juniper woodlands: 100 years of plant succession. In: Shaw, Douglas W.; Aldon, Earl F.; LoSapio, Carol, tech. coords. Desired future conditions for pinon-juniper ecosystems: Proceedings of the symposium; 1994 August 8-12; Flagstaff, AZ. Gen. Tech. Rep. RM-258. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 5-8. [24790]
430. Miller, Rick; Svejcar, Tony; Rose, Jeff. 1999. Conversion of shrub steppe to juniper woodland. In: Monsen, Stephen B.; Stevens, Richard, compilers. Proceedings: ecology and management of pinyon-juniper communities within the Interior West: Sustaining and restoring a diverse ecosystem; 1997 September 15-18; Provo, UT. Proceedings RMRS-P-9. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 385-390. [30583]
431. Millspaugh, Sarah H.; Whitlock, Cathy; Bartlein, Patrick J. 2000. Variations in fire frequency and climate over the past 17000 yr in central Yellowstone National Park. Geology. 28(3): 211-214. [91551]
432. Moffet; Corey A; Taylor, J. Bret; Booth, D. Terrance. 2015. Postfire shrub cover dynamics: A 70-year fire chronosequence in mountain big sagebrush communities. Journal of Arid Environments. 114: 116-123. [89591]
433. Moir, W. H.; Hendzel, Leonard, tech. coords. 1983. Proceedings of the workshop on Southwestern habitat types; 1983 April 6-8; Albuquerque, NM. Albuquerque, NM: U.S. Department of Agriculture, Forest Service, Southwestern Region. 110 p. [1673]
434. Monsen, S. B.; Meyer, S. E. 1990. Planning, rehabilitation, and treatment of disturbed lands: Seeding equipment effects on establishment of big sagebrush on mine disturbances. In: Munshower, Frank F.; Fisher, Scott E., Jr., co-chairmen. Proceedings: Fifth Billings Symposium on Disturbed Land Rehabilitation: 1990 March 25-30; Billings, MT. Volume 1: Hardrock waste, analytical and revegetation. Helena, MT: Montana State University: 192-199. [91229]
435. Monsen, S. B.; Shaw, N. L. 1997. Persistence of cheatgrass (Bromus tectorum) amid bunchgrass/shrubsteppe communities. In: Grasslands 2000: Proceedings, 18th international grassland congress; 1997 June 8-18; Winnipeg, MB. Saskatoon, SK: International Grasslands Congress. Session 21.2: 27-28. [33480]
436. Monsen, Stephen B. 2000. Establishment of big sagebrush (Artemisia tridentata) in semiarid environments. In: Entwistle, P. G.; DeBolt, J. H.; Kaltenecker, J. H.; Steenhof, K., compilers. Sagebrush steppe ecosystems symposium: Proceedings; 1999 June 21-23; Boise, ID. Publication No. PLM/ID/PT-001001+1150. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office: 81-86. [41931]
437. Monsen, Stephen B.; McArthur, E. Durant. 1985. Factors influencing establishment of seeded broadleaf herbs and shrubs following fire. In: Saunders, Ken; Durham, Jack; [and others], eds. Rangeland fire effects: Proceedings of the symposium; 1984 November 27-29; Boise, ID. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office: 112-124. [1682]
438. Monsen, Stephen B.; Meyer, Susan E.; Carlson, Stephanie L. 1992. Sagebrush establishment enhanced by snowfencing. In: Rangeland Technology Equipment Council: 1992 annual report. 2200-Range, 922-2842-MTDC. Washington, DC: U.S. Department of Agriculture, Forest Service, Technology and Development Program: 6-8. [91046]
439. Monsen, Stephen B.; Shaw, Nancy L. 2000. Big sagebrush (Artemisia tridentata) communities--ecology, importance and restoration potential. In: Schuman, Gerald E.; Richmond, Timothy C.; Neuman, Dennis R., eds. Sagebrush establishment on mined lands: Ecology and research: Proceedings of the symposium; 2000 March 20-24; Billings, MT. Bozeman, MT: Montana State University, Reclamation Research Unit: 1-15. [42417]
440. Morgan, Penelope; Heyerdahl, Emily K.; Gibson, Carly E. 2008. Multi-season climate synchronized forest fires throughout the 20th century, Northern Rockies, USA. Ecology. 89(3): 717-728 [+ supplementary information]. [90451]
441. Morris, Christo; Badik, Kevin J.; Morris, Lesley R.; Weltz, Mark A. 2016. Integrating precipitation, grazing, past effects and interactions in long-term vegetation change. Journal of Arid Environments. 124: 111-117. [91050]
442. Morris, Melvin S.; Kelsey, Rick G.; Griggs, Dave. 1976. The geographic and ecological distribution of big sagebrush and other woody Artemisias in Montana. Proceedings of the Montana Academy of Sciences. 36: 56-79. [1695]
443. Mosley, Jeffrey C. 1996. Prescribed sheep grazing to suppress cheatgrass: A review. Sheep & Goat Research Journal. 12(2): 74-81. [30475]
444. Mueggler, W. F. 1967. Voles damage big sagebrush in southwestern Montana. Journal of Range Management. 20(2): 88-91. [91417]
445. Mueggler, W. F. 1985. Vegetation associations. In: DeByle, Norbert V.; Winokur, Robert P., eds. Aspen: Ecology and management in the western United States. Gen. Tech. Rep. RM-119. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 45-55. [11907]
446. Mueggler, W. F.; Stewart, W. L. 1980. Grassland and shrubland habitat types of western Montana. Gen. Tech. Rep. INT-66. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 154 p. [1717]
447. Mueggler, Walter F. 1956. Is sagebrush seed residual in the soil of burns or is it wind-borne? Research Note No. 35. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 10 p. [1704]
448. Mueggler, Walter F. 1976. Ecological role of fire in western woodland and range ecosystems. In: Use of prescribed burning in western woodland and range ecosystems: Proceedings of the symposium; 1976 March 18-19; Logan, UT. Logan, UT: Utah State University, Utah Agricultural Experiment Station: 1-9. [1709]
449. Mueggler, Walter F.; Campbell, Robert B., Jr. 1982. Aspen community types on the Caribou and Targhee National Forests in southeastern Idaho. Res. Pap. INT-294. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 32 p. [1713]
450. Mumma, John W. 1990. High-mountain resources on National Forest lands. In: Schmidt, Wyman C.; McDonald, Kathy J., compilers. Proceedings--symposium on whitebark pine ecosystems: Ecology and management of a high-mountain resource; 1989 March 29-31; Bozeman, MT. Gen Tech. Rep. INT-270. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 49-52. [11674]
451. Nagy, Julius G. 1979. Wildlife nutrition and the sagebrush ecosystem. In: The sagebrush ecosystem: A symposium: Proceedings; 1978 April; Logan, UT. Logan, UT: Utah State University, College of Natural Resources: 164-168. [1729]
452. NatureServe. 2013. International Ecological Classification Standard: Terrestrial Ecological Classifications of the United States and Canada. In: NatureServe Central Databases. Arlington, VA, (Producer). 1530 p. [89169]
453. NatureServe. 2016. International ecological classification standard: Terrestrial ecological classifications, [Online]. In: NatureServe Central Databases. Arlington, VA: NatureServe (Producer). Available: http://explorer.natureserve.org/servlet/NatureServe?init=Ecol [2015, May 1]. [87721]
454. Neilson, Ronald P.; Lenihan, James M.; Bachelet, Dominique; Drapek, Raymond J. 2005. Climate change implications for sagebrush ecosystems. In: Sparrowe, Rollin D., chair; Carpenter, Len H., cochair. The sage-grouse dilemma: A case study of long-term landscape use and abuse; 2005 March 16-19; Arlington, VA. Transactions of the 70th North American Wildlife and Natural Resources Conference. Washington, D.C.: Wildlife Management Institute: 145-159. [91458]
455. Nelle, Pamela J.; Reese, Kerry P.; Connelly, John W. 2000. Long-term effects of fire on sage grouse habitat. Journal of Range Management. 53(6): 586-591. [37079]
456. Nelson, David L.; Harper, Kimball T.; Boyer, Kenneth C.; Weber, Darrell J.; Haws, B. Austin; Marble, James R. 1989. Wildland shrub dieoffs in Utah: An approach to understanding the cause. In: Wallace, Arthur; McArthur, E. Durant; Haferkamp, Marshall R., compilers. Proceedings--symposium on shrub ecophysiology and biotechnology; 1987 June 30 - July 2; Logan, UT. Gen. Tech. Rep. INT-256. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 119-135. [5942]
457. Nelson, David L.; Tiernan, Charles F. 1983. Winter injury of sagebrush and other wildland shrubs in the western United States. Research Paper INT-314. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 17 p. [1741]
458. Nelson, N. A.; Pierce, J. 2010. Late Holocene relationships among fire, climate, and vegetation in a forest-sagebrush ecotone of southwestern Idaho, USA. The Holocene. 20(8): 1179-1194. [90220]
459. Nelson, Zachary J. 2017. Personal communication [Email to Robin Innes]. 6 November. Unpublished data and information regarding mountain big sagebrush postfire recovery. Reno, NV: University of Nevada, Department of Natural Resources and Environmental Science, Great Basin Landscape Ecology Lab. Documents on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92324]
460. Nelson, Zachary J.; Weisberg, Peter J.; Kitchen, Stanley G. 2014. Influence of climate and environment on post-fire recovery of mountain big sagebrush. International Journal of Wildland Fire. 23(1): 131-142. [87792]
461. Neuenschwander, L. F. 1978. The fire induced autecology of selected shrubs of the cold desert and surrounding forests: A-state-of-the-art review. Unpublished manuscript on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 31 p. [1747]
462. Neuenschwander, L. F. 1980. Broadcast burning of sagebrush in the winter. Journal of Range Management. (33)3: 233-236. [1746]
463. Nevada Department of Conservation and Natural Resources, Nevada Natural Heritage Program. 2003. National vegetation classification for Nevada, [Online]. Carson City, NV: Nevada Department of Conservation and Natural Resources, Nevada Natural Heritage Program (Producer). 15 p. Available: http://heritage.nv.gov/ecology/nv_nvc.htm [2005, November 3]. [55021]
464. Newsome, Heidi. 2016. Personal communication [Email to Robin Innes]. 1 December. Regarding use of Rx fire in big sagebrush communities within the Mid-Columbia River National Wildlife Refuge Complex. Burbank, WA: U.S. Department of the Interior, US Fish and Wildlife Service, Mid-Columbia River National Wildlife Refuge Complex. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [91634]
465. Ngugi, Kinuthia R.; Powell, Jeff; Hinds, Frank C.; Olson, Richard A. 1992. Range animal diet composition in southcentral Wyoming. Journal of Range Management. 45(6): 542-545. [19781]
466. Nichols, R.; Menke, J. 1984. Effects of chaparral shrubland fire on terrestrial wildlife. In: DeVries, Johannes J., ed. Shrublands in California: Literature review and research needed for management. Contribution No. 191. Davis, CA: University of California, Water Resources Center: 74-97. [5706]
467. Noson, Anna C. 2002. Avian communities in relation to habitat influenced by fire in a sagebrush steppe landscape. Corvallis, OR: Oregon State University. 77 p. [+ appendices]. Thesis. [91597]
468. Noson, Anna C.; Schmitz, Richard A.; Miller, Richard F. 2006. Influence of fire and juniper encroachment on birds in high-elevation sagebrush steppe. Western North American Naturalist. 66(3): 343-353. [65137]
469. Noste, Nonan V.; Bushey, Charles L. 1987. Fire response of shrubs of dry forest habitat types in Montana and Idaho. Gen. Tech. Rep. INT-239. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 22 p. [255]
470. Nydegger, Nicholas C.; Smith, Graham W. 1986. Prey populations in relation to Artemisia vegetation types in southwestern Idaho. In: McArthur, E. Durant; Welch, Bruce L., compilers. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 152-156. [1787]
471. O'Connor, Casey; Miller, Rick; Bates, Jonathan D. 2013. Vegetation response to western juniper slash treatments. Environmental Management. 52(3): 553-566. [87565]
472. O'Meara, Timothy E.; Haufler, Jonathan B.; Stelter, Lavern H.; Nagy, Julius G. 1981. Nongame wildlife responses to chaining of pinyon-juniper woodlands. The Journal of Wildlife Management. 45(2): 381-389. [4502]
473. Oldemeyer, John L.; Martin, Stephen J.; Woodis, Steven G. 1983. A preliminary report on the effects of a deferred-rotation grazing system on wildlife at the Sheldon National Wildlife Refuge. Cal-Neva Wildlife Transactions 1983: 26-42. [91437]
474. Olson, Rich. 1992. Mule deer habitat requirements and management in Wyoming. B-965. Laramie, WY: University of Wyoming, Cooperative Extension Service. 15 p. [20679]
475. Omernik, James M.; Griffith, Glenn E. 2014. Ecoregions of the conterminous United States: Evolution of a hierarchical spatial framework. Environmental Management. 54(6): 1249-1266. [92352]
476. Ostoja, Steven M.; Schupp, Eugene W. 2009. Conversion of sagebrush shrublands to exotic annual grasslands negatively impacts small mammal communities. Diversity and Distributions. 15(5): 863-870. [91523]
477. Ottmar, Roger D.; Vihnanek, Robert E.; Wright, Clinton S. 2007. Stereo photo series for quantifying natural fuels. Volume X: Sagebrush with grass and ponderosa pine-juniper types in central Montana. Gen. Tech. Rep. PNW-GTR-719. Seattle, WA: U.S. Department of Agriculture, Pacific Northwest Research Station. 59 p. [91260]
478. Paige, Christine; Ritter, Sharon A. 1999. Birds in a sagebrush sea: Managing sagebrush habitats for bird communities. Boise, ID: Partners in Flight Western Working Group. 47 p. [65948]
479. Palmquist, Kyle A.; Schlaepfer, Daniel R.; Bradford, John B.; Lauenroth, William K. 2016. Spatial and ecological variation in dryland ecohydrological responses to climate change: Implications for management. Ecosphere. 7(11): 1-20. [91505]
480. Parish, Roberta; Coupe, Ray; Lloyd, Dennis, eds. 1996. Plants of southern interior British Columbia. Vancouver, BC: Lone Pine Publishing. 450 p [+ supplementary information]. [35949]
481. Passey, H. B.; Hugie, Vern K.; Williams, E. W.; Ball, D. E. 1982. Relationships between soil, plant community, and climate on rangelands of the Intermountain West. Tech. Bull. 1669. Washington, DC: U.S. Department of Agriculture, Soil Conservation Service. 123 p. [1834]
482. Patterson, Robert L. 1952. The sage grouse in Wyoming. Federal Aid to Wildlife Restoration Project 28-R. Denver, CO: Sage Books, Inc. 341 p. [7513]
483. Pearson, Scott M.; Turner, Monica G.; Wallace, Linda L.; Romme, William H. 1995. Winter habitat use by large ungulates following fire in northern Yellowstone National Park. Ecological Applications. 5(3): 744-755. [26077]
484. Pechanec, J. F.; Pickford, G. D.; Stewart, George. 1937. Effects of the 1934 drought on native vegetation of the Upper Snake River Plains, Idaho. Ecology. 18(4): 490-505. [91047]
485. Pechanec, Joseph F.; Plummer, A. Perry; Robertson, Joseph H.; Hull, A. C., Jr. 1965. Sagebrush control on rangelands. Agriculture Handbook No. 277. Washington, DC: U.S. Department of Agriculture. 40 p. [1858]
486. Pechanec, Joseph F.; Stewart, George; Blaisdell, James P. 1954. Sagebrush burning--good and bad. Farmers' Bulletin No. 1948. Washington, DC: U.S. Department of Agriculture. 34 p. [1859]
487. Pedersen, E. K.; Connelly, J. W.; Hendrickson, J. R.; Grant, W. E. 2003. Effect of sheep grazing and fire on sage grouse populations in southeastern Idaho. Ecological Modelling. 165(1): 23-47. [90036]
488. Peek, James M. 2003. Wapiti (Cervus elaphus). In: Feldhamer, George A.; Thompson, Bruce C.; Chapman, Joseph A., eds. Wild mammals of North America: Biology, management, and conservation. 2nd ed. Baltimore, MD: The Johns Hopkins University Press: 877-888. [82099]
489. Peek, James M.; Riggs, Robert A.; Lauer, Jerry L. 1979. Evaluation of fall burning on bighorn sheep winter range. Journal of Range Management. 32(6): 430-432. [1863]
490. Peek, James, M.; Demarchi, Dennis A.; Demarchi, Raymond A.; Stucker, Donald E. 1985. Bighorn sheep and fire: Seven case histories. In: Lotan, James E.; Brown, James K., compilers. Fire's effect on wildlife habitat--symposium proceedings; 1984 March 21; Missoula, MT. Gen. Tech. Rep. INT-186. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 36-43. [1864]
491. Pehrson, Krystle A.; Sowell, Bok F. 2011. Converting crested wheatgrass stands to enhance big sagebrush: A literature review. In: Wambolt, Carl L.; Kitchen, Stanley G.; Frisina, Michael R.; Sowell, Bok; Keigley, Richard B.; Palacios, Patsy; Robinson, Jill, comps. Proceedings--shrublands: Wildlands and wildlife habitats; 15th wildland shrub symposium; 2008 June 17-19; Bozeman, MT. Natural Resources and Environmental Issues, Volume XVI. Logan, UT: Utah State University, College of Natural Resources, S. J. and Jessie E. Quinney Natural Resources Research Library: 121-130. [83476]
492. Peirce, E. S. 1915. The regeneration of denuded areas in the Bighorn Mountains by Douglas-fir. Forestry Quarterly. 13(3): 303-307. [39038]
493. Pellant, Mike. 1990. The cheatgrass-wildfire cycle--are there any solutions? In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., compilers. Proceedings--symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 11-18. [12730]
494. Pellant, Mike. 1994. History and applications of the Intermountain greenstripping program. In: Monsen, Stephen B.; Kitchen, Stanley G., compilers. Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 63-68. [24254]
495. Pendleton, Rosemary L.; Pendleton, Burton K.; Harper, Kimball T. 1989. Breeding systems of woody plant species in Utah. In: Wallace, Arthur; McArthur, E. Durant; Haferkamp, Marshall R., comps. Proceedings--symposium on shrub ecophysiology and biotechnology; 1987 June 30 - July 2; Logan, UT. Gen. Tech. Rep. INT-256. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 5-22. [5918]
496. Perfors, Tracy; Harte, John; Alter, S. Elizabeth. 2003. Enhanced growth of sagebrush (Artemisia tridentata) in response to manipulated ecosystem warming. Global Change Biology. 9(5): 736-742. [91459]
497. Perryman, B. L.; Maier, A. M.; Hild, A. L.; Olson, R. A. 2000. Demographic characteristics of three Artemisia tridentata subspecies. In: Schuman, Gerald E.; Richmond, Timothy C.; Neuman, Dennis R., eds. Sagebrush establishment on mined lands: Ecology and research: Proceedings of symposium; 2000 March 20; Billings, MT. [Part of: 2000 Billings land reclamation symposium; 2000 March 20-24; Billings, MT]. Bozeman, MT: Montana State University, Reclamation Research Unit: 56-64. [42422]
498. Perryman, Barry L.; Maier, Aaron M. ; Hild, Ann L.; Olson, Richard A. 2001. Demographic characteristics of 3 Artemisia tridentata Nutt. subspecies. Journal of Range Management. 54(2): 166-170. [39049]
499. Perryman, Barry L.; Olson, Richard A. 2000. Age-stem diameter relationships of big sagebrush and their management implications. Journal of Range Management. 53(3): 342-346. [35851]
500. Personius, Timothy L.; Wambolt, Carl L.; Stephens, Jeffrey R.; Kelsey, Rick G. 1987. Crude terpenoid influence on mule deer preference for sagebrush. Journal of Range Management. 40(1): 84-88. [1872]
501. Peters, Erin F.; Bunting, Stephen C. 1994. Fire conditions pre-and post-occurrence of annual grasses on the Snake River Plain. In: Monsen, Stephen B.; Kitchen, Stanley G., comps. Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 31-36. [24249]
502. Petersen, Kenneth L.; Best, Louis B. 1987. Effects of prescribed burning on nongame birds in a sagebrush community. Wildlife Society Bulletin. 15: 317-329. [173]
503. Petersen, Steven L.; Stringham, Tamzen K.; Roundy, Bruce A. 2009. A process-based application of state-and-transition models: A case study of western juniper (Juniperus occidentalis) encroachment. Rangeland Ecology & Management. 62(2): 186-192. [75456]
504. Peterson, Eric B. 2008. International vegetation classification alliances and associations occurring in Nevada with proposed additions. Carson City, NV: Nevada Natural Heritage Program. 347 p. Available online: http://heritage.nv.gov/sites/default/files/library/ivclist.pdf [2018, January 10]. [77864]
505. Peterson, Joel G. 1995. Ecological implications of sagebrush manipulation: A literature review. Project W-101-R-2. Helena, MT: Montana Department of Fish, Wildlife and Parks. 48 p. [+ appendix]. [91444]
506. Pfister, Robert D.; Kovalchik, Bernard L.; Arno, Stephen F.; Presby, Richard C. 1977. Forest habitat types of Montana. Gen. Tech. Rep. INT-34. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 174 p. [1878]
507. Pickett, Brent M.; Isackson, Carl; Wunder, Rebecca; Fletcher, Thomas H.; Butler, Bret W.; Weise, David R. 2010. Experimental measurements during combustion of moist individual foliage samples. International Journal of Wildland Fire. 19(2): 153-162. [81950]
508. Pietruszka, Brad. 2016. Personal communication [Email to Robin Innes]. 18 May. Regarding use of Rx fire in Gunnison sage-grouse habitats in the Tres Rios field office. Montrose, CA: U.S. Department of the Interior, Bureau of Land Management, Southwest District. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [90477]
509. Plummer, A. Perry. 1976. Shrubs for the subalpine zone of the Wasatch Plateau. In: Zuck, R. H.; Brown, L. F., eds. High altitude revegetation workshop--No. 2: Proceedings of a workshop; 1976 April 5-6; Fort Collins, CO. Information Series No. 21. Fort Collins, CO: Colorado State University; Colorado Water Resources Research Institute: 33-40. [1899]
510. Plummer, A. Perry; Christensen, Donald R.; Monsen, Stephen B. 1968. Restoring big-game range in Utah. Publ. No. 68-3. Ephraim, UT: Utah Division of Fish and Game. 183 p. [4554]
511. Poore, Rebecca E.; Lamanna, Christine A.; Ebersole, James J.; Enquist, Brian J. 2009. Controls on radial growth of mountain big sagebrush and implications for climate change. Western North American Naturalist. 69(4): 556-562. [90865]
512. Powell, David C.; Johnson, Charles G., Jr.; Crowe, Elizabeth A.; Wells, Aaron; Swanson, David K. 2007. Potential vegetation hierarchy for the Blue Mountains section of northeastern Oregon, southeastern Washington, and west-central Idaho. Gen. Tech. Rep. PNW-GTR-709. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 87 p. [69013]
513. Prenzlow, E. J.; Gilbert, D. L.; Glover, F. A. 1968. Some behavior patterns of the pronghorn. Special Report No. 17/GFP-R-S 17. Denver, CO: Colorado Department of Game, Fish and Parks. 16 p. [25691]
514. Provencher, Louis; Blankenship, Kori. 2016. Personal communication [Email to Robin Innes]. 12 October. Regarding big sagebrush. Reno, NV: The Nature Conservancy, Nevada Field Office. Email on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [91913]
515. Pyke, David A.; Wirth, Troy A.; Beyers, Jan L. 2013. Does seeding after wildfires in rangelands reduce erosion or invasive species? Restoration Ecology. 21(4): 415-421. [87983]
516. Pyle, William H.; Crawford, John A. 1996. Availability of foods of sage grouse chicks following prescribed fire in sagebrush-bitterbrush. Journal of Range Management. 49(4): 320-324. [26885]
517. Qi, Yi; Jolly, W. Matt; Dennison, Philip E.; Kropp, Rachael C. 2016. Seasonal relationships between foliar moisture content, heat content and biochemistry of lodgepole pine and big sagebrush foliage. International Journal of Wildland Fire. 25(5): 574-578. [90925]
518. Raper, Bob; Clark, Bob; Matthews, Marion; Aldrich, Ann. 1985. Early effects of a fall burn on a western Wyoming mountain big sagebrush-grass community. In: Saunders, Ken; Durham, Jack; [and others], eds. Rangeland fire effects: Proceedings of the symposium; 1984 November 27-29; Boise, ID. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office: 88-92. [1938]
519. Rau, B. M.; Blank, R. R.; Chambers, J. C.; Johnson, D. W. 2007. Prescribed fire in a Great Basin sagebrush ecosystem: Dynamics of soil extractable nitrogen and phosphorus. Journal of Arid Environments. 71(4): 362-375. [68653]
520. Rau, Benjamin M.; Chambers, Jeanne C.; Blank, Robert R.; Johnson, Dale W. 2008. Prescribed fire, soil, and plants: Burn effects and interactions in the central Great Basin. Rangeland Ecology and Management. 61(2): 169-181. [71125]
521. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford, England: Clarendon Press. 632 p. [2843]
522. Reichenberger, Gunther; Pyke, David A. 1990. Impact of early root competition on fitness components of four semiarid species. Oecologia. 85(2): 159-166. [91230]
523. Reiner, Alicia L. 2004. Fuel load and understory community changes associated with varying elevation and pinyon-juniper dominance. Reno, NV: University of Nevada. 63 p. Thesis. [91569]
524. Reiner, Alicia L.; Tausch, Robin J.; Walker, Roger F. 2010. Estimation procedures for understory biomass and fuel loads in sagebrush steppe invaded by woodlands. Western North American Naturalist. 70(3): 312-322. [82208]
525. Reinkensmeyer, Daniel P.; Miller, Richard F.; Anthony, Robert G.; Marr, Vern E. 2000. Avian community structure along a mountain big sagebrush successional gradient. Journal of Wildlife Management. 71(4): 1057-1066. [91436]
526. Reisner, Michael D.; Grace, James B.; Pyke, David A.; Doescher, Paul S. 2013. Conditions favouring Bromus tectorum dominance of endangered sagebrush steppe ecosystems. Journal of Applied Ecology. 50(4): 1039-1049. [91615]
527. Rens, Reyer Jan. 2001. Elk effects on sagebrush-grassland after fire on Yellowstone's Northern Range. Bozeman, MT: Montana State University. 61 p. Thesis. [46927]
528. Richards, J. H.; Caldwell, M. M. 1987. Hydraulic lift: Substantial nocturnal water transport between soil layers by Artemisia tridentata. Oecologia. 73(4): 486-489. [16213]
529. Richardson, Bland Z.; Monsen, Stephen B.; Bowers, Diane M. 1986. Interseeding selected shrubs and herbs on mine disturbances in southeastern Idaho. In: McArthur, E. Durant; Welch, Bruce L., compilers. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 134-139. [1973]
530. Richardson, Bryce A.; Chaney, Lindsay; Shaw, Nancy L.; Still, Shannon M. 2016. Will phenotypic plasticity affecting flowering phenology keep pace with climate change? Global Change Biology: 1-10. doi: 10.1111/gcb.13532. [91682]
531. Richardson, Bryce A.; Page, Justin T.; Bajgain, Prabin; Sanderson, Stewart C.; Udall, Joshua A. 2012. Deep sequencing of amplicons reveals widespread intraspecific hybridization and multiple origins of polyploidy in big sagebrush (Artemisia tridentata; Asteraceae). American Journal of Botany. 99(12): 1962-1975. [91016]
532. Rickard, W. H.; Keough, R. F. 1968. Soil-plant relationships of two steppe desert shrubs. Plant and Soil. 29(2): 205-211. [1983]
533. Riegel, Gregg M.; Miller, Richard F.; Skinner, Carl N.; Smith, Sydney E. 2006. Northeastern Plateaus bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 225-263. [65541]
534. Ripplinger, Julie; Franklin, Janet; Edwards, Thomas C., Jr. 2015. Legacy effects of no-analogue disturbances alter plant community diversity and composition in semi-arid sagebrush steppe. Journal of Vegetation Science. 26(5): 923-933. [91611]
535. Roberson, Jay A. 1986. Sage grouse-sagebrush relationships: A review. In: McArthur, E. Durant; Welch, Bruce L., compilers. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 157-167. [2006]
536. Roberts, Thomas C., Jr. 1999. The budgetary, ecological, and managerial impacts of pinyon-juniper and cheatgrass fires. In: Monsen, Stephen B.; Stevens, Richard, compilers. Proceedings: Ecology and management of pinyon-juniper communities within the Interior West: Sustaining and restoring a diverse ecosystem; 1997 September 15-18; Provo, UT. Proceedings RMRS-P-9. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 400-402. [30587]
537. Robertson, Mark D. 1991. Winter ecology of migratory sage grouse and associated effects of prescribed fire in southeastern Idaho. Moscow, ID: University of Idaho. 88 p. Thesis. [35404]
538. Rodriguez, Ronald L.; Welch, Bruce L. 1989. Effects of heavy grazing by mule deer on 'Hobble Creek' mountain big sagebrush seed stalk production. In: Wallace, Arthur; McArthur, E. Durant; Haferkamp, Marshall R., compilers. Proceedings--symposium on shrub ecophysiology and biotechnology; 1987 June 30 - July 2; Logan, UT. Gen. Tech. Rep. INT-256. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 141-143. [5943]
539. Romme, William H.; Allen, Craig D.; Bailey, John D.; Baker, William L.; Bestelmeyer, Brandon T.; Brown, Peter M.; Eisenhart, Karen S.; Floyd, M. Lisa; Huffman, David W.; Jacobs, Brian F.; Miller, Richard F.; Muldavin, Esteban H.; [and others]. 2009. Historical and modern disturbance regimes, stand structures, and landscape dynamics in Pinon-juniper vegetation of the western United States. Rangeland Ecology & Management. 62(3): 203-222. [75419]
540. Rose, Jeffrey A.; Eddleman, Lee E. 1994. Ponderosa pine and understory growth following western juniper removal. Northwest Science. 68(2): 79-85. [23145]
541. Rosentreter, Roger. 2005. Sagebrush identification, ecology, and palatability relative to sage-grouse. In: Shaw, Nancy L.; Pellant, Mike; Monsen, Stephen B., eds. Sage-grouse habitat restoration symposium proceedings; 2001 June 4-7; Boise, ID. Proc. RMRS-P-38. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 3-16. [63178]
542. Rosentreter, Roger; Kelsey, Rick G. 1991. Xeric big sagebrush, a new subspecies in the Artemisia tridentata complex. Journal of Range Management. 44(4): 330-335. [15479]
543. Rothleutner, Andy. 2016. Personal communication [Email with Robin Innes]. 20 May. Regarding use of Rx fire in greater sage-grouse habitats within the Worland field office. Worland, WY: U.S. Department of the Interior, Bureau of Land Management, Wind River/Bighorn Basin District, Worland Field Office. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [90499]
544. Rottler, Caitlin M.; Noseworthy, Cara E.; Fowers, Beth; Beck, Jeffrey L. 2015. Effects of conversion from sagebrush to non-native grasslands on sagebrush-associated species. Rangelands. 37(1): 1-6. [90402]
545. Rowland, Mary M.; Suring, Lowell H.; Tausch, Robin J.; Geer, Susan; Wisdom, Michael J. 2011. Dynamics of western juniper woodland expansion into sagebrush communities in central Oregon. In: Wambolt, Carl L.; Kitchen, Stanley G.; Frisina, Michael R.; Sowell, Bok; Keigley, Richard B.; Palacios, Patsy; Robinson, Jill, comps. Proceedings--shrublands: Wildlands and wildlife habitats; 15th wildland shrub symposium; 2008 June 17-19; Bozeman, MT. Natural Resources and Environmental Issues, Volume XVI, Article 13. Logan, UT: Utah State University, College of Natural Resources, S. J. and Jessie E. Quinney Natural Resources Research Library: 89-99. [83473]
546. Sankey, Joel B.; Wallace, Cynthia S. A.; Ravi, Sujith. 2013. Phenology-based, remote sensing of post-burn disturbance windows in rangelands. Ecological Indicators. 30: 35-44. [88824]
547. Sawyer, Hall; Nielson, Ryan M.; Lindzey, Fred G.; Keith, Lorraine; Powell, Jake H.; Abraham, Anu A. 2007. Habitat selection of Rocky Mountain elk in a nonforested environment. The Journal of Wildlife Management. 71(3): 868-874. [83401]
548. Sawyer, John O.; Keeler-Wolf, Todd. 2007. Alpine vegetation. In: Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. Terrestrial vegetation of California. Berkeley, CA: University of California Press: 539-573. [82712]
549. Schell, Andrew. 2017. Personal communication [Email to Robin Innes]. 24 January. Regarding use of Rx fire in big sagebrush communities within the Charles M. Russell NWR. Spokane, WA: U.S. Department of the Interior, U.S. Department of the Interior, US Fish and Wildlife Service, Charles M. Russell National Wildlife Refuge. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [91637]
550. Schlaepfer, Daniel R., Lauenroth, William K.; Bradford, John B. 2012. Ecohydrological niche of sagebrush ecosystems. Ecohydrology. 5(4): 453-466. [91550]
551. Schlaepfer, Daniel R.; Lauenroth, William K.; Bradford, John B. 2012. Effects of ecohydrological variables on current and future ranges, local suitability patterns, and model accuracy in big sagebrush. Ecography. 35(4): 374-384. [90149]
552. Schlaepfer, Daniel R.; Taylor, Kyle A.; Pennington, Victoria E.; Nelson, Kellen N.; Martyn, Trace E.; Rottler, Caitlin M.; Lauenroth, William K.; Bradford, John B. 2015. Simulated big sagebrush regeneration supports predicted changes at the trailing and leading edges of distribution shifts. Ecosphere. 6(1): 1-31. [91461]
553. Schlatterer, Edward F. 1972. A preliminary description of plant communities found on the Sawtooth, White Cloud, Boulder and Pioneer Mountains. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Region. Unpublished paper on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 111 p. [2076]
554. Schneegas, Edward R. 1967. Sage grouse and sagebrush control. Transactions, North American Wildlife Conference. 32: 270-274. [4933]
555. Scholl, Jackson P.; Kelsey, Rick G.; Shafizadeh, Fred. 1977. Involvement of volatile compounds of Artemisia in browse preference by mule deer. Biochemical Systematics and Ecology. 5: 291-295. [2086]
556. Schroeder, Michael A.; Connelly, John W.; Wambolt, Carl L.; Braun, Calit E.; Hagen, Christian A.; Frisina, Michael R. 2006. View points: Society for range management issue paper: Ecology and management of sage-grouse and sage-grouse habitat: A reply. Rangelands. 28(3): 3-7. [89733]
557. Schupp, Eugene W.; Gomez, Jose M.; Jimenez, Jaime E.; Fuentes, Marcelino. 1997. Dispersal of Juniperus occidentalis (western juniper) seeds by frugivorous mammals on Juniper Mountain, southeastern Oregon. The Great Basin Naturalist. 57(1): 74-78. [27379]
558. Scott, M. Douglas; Geisser, Hannes. 1996. Pronghorn migration and habitat use following the 1988 Yellowstone fires. In: Greenlee, Jason, ed. The ecological implications of fire in Greater Yellowstone: Proceedings, 2nd biennial conference on the Greater Yellowstone Ecosystem; 1993 September 19-21; Yellowstone National Park, WY. Fairfield, WA: International Association of Wildland Fire: 123-132. [27837]
559. Severson, Kieth E. 1984. Small mammals in modified pinyon-juniper woodlands, New Mexico. Journal of Range Management. 39(1): 31-34. [91598]
560. Severson, Kieth E.; Medina, Alvin L. 1983. Deer and elk habitat management in the Southwest. Journal of Range Management. Monograph No. 2. Denver, CO: Society for Range Management. 64 p. [2110]
561. Shafer, Sarah L.; Bartlein, Patrick J.; Thompson, Robert S. 2001. Potential changes in the distribution of western North America tree and shrub taxa under future climate scenarios. Ecosystems. 4(3): 200-215. [91465]
562. Shantz, H. L. 1947. The use of fire as a tool in the management of the brush ranges of California. Sacramento, CA: State of California, Department of Natural Resources, Division of Forestry. 156 p. [36305]
563. Shaw, M. Rebecca; Loik, Michael E.; Harte, John. 2000. Gas exchange and water relations of two Rocky Mountain shrub species exposed to climate change manipulation. Plant Ecology. 146(2): 197-206. [90978]
564. Shaw, Nancy L.; Monsen, Stephen B. 1990. Use of sagebrush for improvement of wildlife habitat. In: Fisser, Herbert G., ed. Wyoming shrublands: Aspen, sagebrush and wildlife management: Proceedings, 17th Wyoming shrub ecology workshop; 1988 June 21-22; Jackson, WY. Laramie, WY: University of Wyoming, Department of Range Management; Shrub Ecology Workshop: 19-35. [22929]
565. Sheehy, Dennis P.; Winward, A. H. 1981. Relative palatability of seven Artemisia taxa to mule deer and sheep. Journal of Range Management. 34(5): 397-399. [2128]
566. Sheehy, Dennis Patrick. 1975. Relative palatability of seven Artemisia taxa to mule deer and sheep. Corvalis, OR: Oregon State University. 132 p. [+ appendices]. Thesis. [91427]
567. Shepperd, Wayne D.; Battaglia, Michael A. 2002. Ecology, silviculture, and management of Black Hills ponderosa pine. Gen. Tech. Rep. RMRS-GTR-97. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 112 p. [44794]
568. Sheridan, Chris. 2016. Personal communication [Email to Robin Innes]. 2 December. Regarding use of Rx fire in big sagebrush communities within the Spokane District. Spokane, WA: U.S. Department of the Interior, U.S. Department of the Interior, Bureau of Land Management, Spokane District. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [91636]
569. Shinn, Dean A. 1980. Historical perspectives on range burning in the inland Pacific Northwest. Journal of Range Management. 33(6): 415-423. [2136]
570. Shive, Kristen L.; Sieg, Carolyn H.; Fule, Peter Z. 2013. Pre-wildfire management treatments interact with fire severity to have lasting effects on post-wildfire vegetation response. Forest Ecology and Management. 297: 75-83. [86959]
571. Shriver, Robert K.; Minckley, Thomas A. 2012. Late-Holocene response of limber pine (Pinus flexilis) to fire disturbance in the pine forest range, Nevada, USA. Quaternary Research. 78(3): 465-473. [87529]
572. Shriver, Robert K.; Minckley, Thomas A. 2013. Corrigendum to "Late-Holocene response of limber pine (Pinus flexilis) to fire disturbance in the pine forest range, Nevada, USA" [Quaternary Research 78 (2012) 465-473]. Quaternary Research. 79(3): 309. [90815]
573. Shultz, Leila M. 2009. Monograph of Artemisia subgenus Tridentatae (Asteraceae-Anthemideae). Systematic Botany Monographs. Laramie, WY: American Society of Plant Taxonomists. 89: 1-131. [90844]
574. Shumar, Mark L.; Anderson, Jay E. 1986. Gradient analysis of vegetation dominated by two subspecies of big sagebrush. Journal of Range Management. 39(2): 156-159. [2142]
575. Shumar, Mark L.; Anderson, Jay E.; Reynolds, Timothy D. 1982. Identification of subspecies of big sagebrush by ultraviolet spectrophotometry. Journal of Range Management. 35(1): 60-62. [30181]
576. Sime, Carolyn Anne. 1991. Sage grouse use of burned, non-burned, and seeded vegetation communities on the Idaho National Engineering Laboratory, Idaho. Bozeman, MT: Montana State University. 72 p. Thesis. [24908]
577. Singer, Francis J.; Coughenour, Michael B.; Norland, Jack E. 2004. Elk biology and ecology before and after the Yellowstone fires of 1988. In: Wallace, Linda L., ed. After the fires: The ecology of change in Yellowstone National Park. New Haven, CT: Yale University Press: 117-139. [81963]
578. Singer, Francis J.; Mack, John A. 1999. Predicting the effects of wildfire and carnivore predation on ungulates. In: Clark, Tim W.; Curlee, A. Peyton; Minta, Steven C.; Karieva, Peter M., eds. Carnivores in ecosystems: The Yellowstone experience. New Haven, CT: Yale University Press: 188-237. [91470]
579. Singer, Francis J.; Renkin, Roy A. 1995. Effects of browsing by native ungulates on the shrubs in big sagebrush communities in Yellowstone National Park. Great Basin Naturalist. 55(3): 201-212. [35476]
580. Skousen, J. G.; Davis, J. N.; Brotherson, J. D. 1989. Pinyon-juniper chaining and seeding for big game in central Utah. Journal of Range Management. 42(2): 98-104. [1297]
581. Skovlin, Jon M.; Zager, Peter; Johnson, Bruce K. 2002. Elk habitat selection and evaluation. In: Toweill, Dale E.; Thomas, Jack Ward, eds. North American elk: Ecology and management. Washington, DC: Smithsonian Institution Press: 531-556. [81803]
582. Slater, Steven J. 2003. Sage-grouse (Centrocercus urophasianus) use of different-aged burns and the effects of coyote control in southwestern Wyoming. Laramie, WY: University of Wyoming. 148 p. [+ supplementary information]. Thesis. [90154]
583. Smith, Bruce N.; Eldredge, Susan; Moulton, David L.; Monaco, Thomas A.; Jones, Angela R.; Hansen, Lee D.; McArthur, E. Durant; Freeman, D. Carl. 1999. Differences in temperature dependence of respiration distinguish subspecies and hybrid populations of big sagebrush: Nature versus nurture. In: McArthur, E. Durant; Ostler, W. Kent; Wambolt, Carl L., compilers. Proceedings: Shrubland ecotones; 1998 August 12-14; Ephraim, UT. Proceedings RMRS-P-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 25-28. [36056]
584. Smith, Michael A. 1985. Prescribed burning of big sagebrush in Wyoming. In: Fisser, Herbert G., ed. Wyoming shrublands: Proceedings, 14th Wyoming shrub ecology workshop; 1985 May 29-30; Rock Springs, WY. Laramie, WY: University of Wyoming, Department of Range Management, Wyoming Shrub Ecology Workshop: 41-45. [13910]
585. Smith, Tom S.; Hardin, Perry J.; Flinders, Jerran T. 1999. Response of bighorn sheep to clear-cut logging and prescribed burning. Wildlife Society Bulletin. 27(3): 840-845. [36492]
586. Soehn, George. 2016. Personal communication [Email to Robin Innes]. 19 May. Regarding use of Rx fire in greater sage-grouse habitats within the Casper field office. Casper, WY: U.S. Department of the Interior, Bureau of Land Management, Wind River/Bighorn Basin District, Casper Field Office. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [90482]
587. Spomer, George G.; Henderson, Douglass M. 1988. Use of UV absorption for identifying subspecies of Artemisia tridentata. Journal of Range Management. 41(5): 395-398. [6260]
588. Stebleton, Andrea; Bunting, Stephen. 2009. Guide for quantifying fuels in the sagebrush steppe and juniper woodlands of the Great Basin. Tech. Note 430. Denver, CO: U.S. Department of the interior, Bureau of Land Management. 81 p. [88764]
589. Steele, Robert; Cooper, Stephen V.; Ondov, David M.; Roberts, David W.; Pfister, Robert D. 1983. Forest habitat types of eastern Idaho-western Wyoming. Gen. Tech. Rep. INT-144. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 122 p. [2230]
590. Steele, Robert; Geier-Hayes, Kathleen. 1993. The Douglas-fir/pinegrass habitat type in central Idaho: Succession and management. Gen. Tech. Rep. INT-298. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 83 p. [21512]
591. Steele, Robert; Pfister, Robert D.; Ryker, Russell A.; Kittams, Jay A. 1981. Forest habitat types of central Idaho. Gen. Tech. Rep. INT-114. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 138 p. [2231]
592. Stefanic, Todd. 2017. Personal communication [Email to Robin Innes]. 23 February. Regarding use of Rx fire in big sagebrush communities within the Craters of the Moon National Monument and Preserve. Arco, ID: U.S. Department of the Interior, National Park Service, Craters of the Moon National Monument and Preserve. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [91635]
593. Stevens, Richard. 1994. Interseeding and transplanting to enhance species composition. In: Monsen, Stephen B.; Kitchen, Stanley G., compilers. Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 300-306. [24301]
594. Stevens, Richard. 2004. Incorporating wildlife habitat needs into restoration and rehabilitation projects. In: Monsen, Stephen B.; Stevens, Richard; Shaw, Nancy L., comps. Restoring western ranges and wildlands. Gen. Tech. Rep. RMRS-GTR-136-vol. 1. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 155-174. [52826]
595. Stevens, Richard; McArthur, E. Durant. 1974. A simple field technique for identification of some sagebrush taxa. Journal of Range Management. 27(4): 325-326. [2245]
596. Stevens, Richard; Monsen, Stephen B. 2004. Guidelines for restoration and rehabilitation of principal plant communities. In: Monsen, Stephen B.; Stevens, Richard; Shaw, Nancy L., comps. Restoring western ranges and wildlands. Gen. Tech. Rep. RMRS-GTR-136-vol. 1. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 199-294. [52829]
597. Stewart, George. 1941. Historic records bearing on agricultural and grazing ecology in Utah. Journal of Forestry. 39: 363-375. [91603]
598. Stickney, Peter F. 1989. Seral origin of species comprising secondary plant succession in northern Rocky Mountain forests. FEIS workshop: Postfire regeneration. Unpublished draft on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 10 p. [20090]
599. Still, Shannon M.; Richardson, Bryce A. 2015. Projections of contemporary and future climate niche for Wyoming big sagebrush (Artemisia tridentata subsp. wyomingensis): A guide for restoration. Natural Areas Journal. 35(1): 30-43. [89167]
600. Strand, Eva K. 2007. Landscape dynamics in aspen and western juniper woodlands on the Owyhee Plateau, Idaho. Moscow, ID: University of Idaho. 158 p. [+ appendices]. Dissertation. [91319]
601. Strand, Eva K.; Bunting, Stephen C.; Keefe, Robert F. 2013. Influence of wildland fire along a successional gradient in sagebrush steppe and western juniper woodlands. Rangeland Ecology and Management. 66(6): 667-679. [87802]
602. Stranton, David J.; McArthur, E. Durant; Freeman, D. Carl; Golenberg, Edward M. 2002. No genetic substructuring in Artemisia subgenus Tridentatae despite strong ecotypic subspecies selection. Biochemical Systematics and Ecology. 30(6): 579-593. [43067]
603. Striby, Karl D.; Wambolt, Carl L.; Kelsey, Rick G.; Havstad, Kris M. 1987. Crude terpenoid influence on in vitro digestibility of sagebrush. Journal of Range Management. 40(3): 244-248. [2265]
604. Stucker, Donald E.; Peek, James M. 1984. Response of bighorn sheep to the Ship Island Burn. Report submitted to the Northern Forest Fire Laboratory: Supplement No. INT-80-108CA. 33 p. Unpublished paper on file at: U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [17070]
605. Sturges, David L. 1977. Soil water withdrawal and root characteristics of big sagebrush. The American Midland Naturalist. 98(2): 257-274. [30167]
606. Sturges, David L. 1989. Response of mountain big sagebrush to induced snow accumulation. Journal of Applied Ecology. 26: 1035-1041. [11841]
607. Sturges, David L.; Nelson, David L. 1986. Snow depth and incidence of a snowmold disease on mountain big sagebrush. In: McArthur, E. Durant; Welch, Bruce L., compilers. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 215-221. [2277]
608. Suminski, Rita R. 1993. Management implications for mule deer winter range in northern pinon-juniper. In: Aldon, Earl F.; Shaw, Douglas W., technical coordinators. Managing pinon-juniper ecosystems for sustainability and social needs: Proceedings; 1993 April 26-30; Santa Fe, NM. Gen. Tech. Rep. RM-236. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 133-139. [22864]
609. Suring, Lowell H.; Rowland, Mary M.; Wisdom, Michael J.; Schueck, Linda; Meinke, Cara W. 2005. Vegetation communities. In: Wisdom, Michael J.; Rowland, Mary M.; Suring, Lowell H., eds. Habitat threats in the sagebrush ecosystem: Methods of regional assessment and applications in the Great Basin. Lawrence, KS: Alliance Communications Group: 94-113. [67402]
610. Suring, Lowell H.; Wisdom, Michael J.; Tausch, Robin J.; Miller, Richard F.; Rowland, Mary M.; Schueck, Linda; Meinke, Cara W. 2005. Modeling threats to sagebrush and other shrubland communities. In: Wisdom, Michael J.; Rowland, Mary M.; Suring, Lowell H., eds. Habitat threats in the sagebrush ecosystem: Methods of regional assessment and applications in the Great Basin. Lawrence, KS: Alliance Communications Group: 114-149. [67403]
611. Swanson, Donald Oscar. 1970. Roosevelt elk-forest relationships in the Douglas-fir region of the southern Oregon Coast Range. Ann Arbor, MI: University of Michigan. 173 p. Dissertation. [83259]
612. Swanson, Sherman R.; Simonson, Gerald H.; Buckhouse, John C. 1986. Physical and chemical soil properties of three big sagebrush subspecies. Soil Science Society of America Journal. 50: 783-787. [3037]
613. Synergy Resource Solutions, Inc. 2002. Data collection and fire modeling determine potential for the use of Plateau to establish fuel breaks in Bromus tectorum dominated rangelands. Sparks, NV: Synergy Resource Solutions, Inc. Unpublished report available on a CD-ROM. On file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. [43417]
614. Tabler, Ronald D. 1964. The root system of Artemisia tridentata at 9,500 feet in Wyoming. Ecology. 45(3): 633-636. [2302]
615. Taper, Mark L.; Gogan, Peter J. 2002. The northern Yellowstone elk: Density dependence and climatic conditions. The Journal of Wildlife Management. 66(1): 106-122. [82436]
616. Tausch, R. J.; Miller, R. F.; Boundy, B. A.; Chambers, J. C., eds. 2009. Pinon and juniper field guide: Asking the right questions to select appropriate management actions. Circular 1335. Reston, VA: U.S. Department of the Interior, Geological Survey. 96 p. [76595]
617. Tausch, R. J.; Tueller, P. T. 1990. Foliage biomass and cover relationships between tree- and shrub- dominated communities in pinyon-juniper woodlands. The Great Basin Naturalist. 50(2): 121-134. [15528]
618. Tausch, Robin J.; Nowak, Robert S. 1999. Fifty years of ecotone change between shrub and tree dominance in the Jack Springs Pinyon Research Natural Area. In: McArthur, E. Durant; Ostler, W. Kent; Wambolt, Carl L., comps. Proceedings: Shrubland ecotones; 1998 August 12-14; Ephraim, UT. Proceedings RMRS-P-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 71-77. [36065]
619. Tausch, Robin J.; Tueller, Paul T. 1977. Plant succession following chaining of pinyon-juniper woodlands in eastern Nevada. Journal of Range Management. 30(1): 44-49. [2305]
620. Tausch, Robin J.; West, Neil E. 1988. Differential establishment of pinyon and juniper following fire. The American Midland Naturalist. 119(1): 174-184. [3671]
621. Taylor, Kyle L. 2015. Exploring ecological risks facing regional sagebrush ecosystems in the 21st century. Laramie, WY: University of Wyoming. 141 p. Thesis. [90565]
622. Taylor, Michael H.; Rollins, Kimberly; Kobayashi, Mimako; Tausch, Robin J. 2013. The economics of fuel management: Wildfire, invasive plants, and the dynamics of sagebrush rangelands in the western United States. Journal of Environmental Management. 126: 157-173. [88818]
623. Terwilliger, Charles, Jr.; Tiedeman, James A. 1978. Habitat types of the mule deer critical winter range and adjacent steppe region of Middle Park, Colorado. Final report: Cooperative Agreement No. 16-739-CA. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 108 p. [5611]
624. The Wildlife Society, Nevada Chapter. 1998. Influence of fire on wildlife habitat in the Great Basin: A position statement - August 16, 1998. Transactions, Western Section of the Wildlife Society. 34: 42-57. [35093]
625. Thorne, Robert F.; Schoenherr, Allan A.; Clements, Charlie D.; Young, James A. 2007. Transmontane coniferous vegetation. In: Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. Terrestrial vegetation of California. Berkeley, CA: University of California Press: 574-586. [82713]
626. Tiedeman, James A.; Francis, Richard E.; Terwilliger, Charles, Jr.; Carpenter, Len H. 1987. Shrub-steppe habitat types of Middle Park, Colorado. Res. Pap. RM-273. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 20 p. [2329]
627. Tipper, Gary K. 1988. Prescribed burning for wildlife in the southern East Kootenay. In: Feller, M. C.; Thomson, S. M., eds. Wildlife and range prescribed burning workshop proceedings; 1987 October 27-28; Richmond, BC. Vancouver, BC: The University of British Columbia, Faculty of Forestry: 61-69. [3100]
628. Tisdale, E. W.; Hironaka, M. 1981. The sagebrush-grass region: A review of the ecological literature. Bull. 33. Moscow, ID: University of Idaho, Forest, Wildlife and Range Experiment Station. 31 p. [2344]
629. Toney, Jaime L.; Anderson, R. Scott. 2006. A postglacial palaeoecological record from the San Juan Mountains of Colorado USA: Fire, climate and vegetation history. The Holocene. 16(4): 505-517. [67025]
630. Toweill, Dale E.; Thomas, Jack Ward. 2002. The future of elk and elk management. In: Toweill, Dale E.; Thomas, Jack Ward, eds. North American elk: Ecology and management. Washington, DC: Smithsonian Institution Press: 793-841. [81807]
631. Tracy, Benjamin F. 2004. Fire effects, elk, and ecosystem resilience in Yellowstone's sagebrush grasslands. In: Wallace, Linda L., ed. After the fires: The ecology of change in Yellowstone National Park. New Haven, CT: Yale University Press: 102-116. [82007]
632. Tracy, Benjamin F.; McNaughton, Samuel J. 1996. Comparative ecosystem properties in summer and winter ungulate ranges following the 1988 fires in Yellowstone National Park. In: Greenlee, Jason, ed. The ecological implications of fire in Greater Yellowstone: Proceedings, 2nd biennial conference on the Greater Yellowstone Ecosystem; 1993 September 19-21; Yellowstone National Park, WY. Fairfield, WA: International Association of Wildland Fire: 181-191. [27847]
633. Tracy, Benjamin F.; McNaughton, Samuel J. 1997. Elk grazing and vegetation responses following a late season fire in Yellowstone National Park. Plant Ecology. 130(2): 111-119. [28963]
634. Trainer, Charles E.; Willis, Mitchell J.; Keister, George P., Jr.; Sheehy, Dennis P. 1983. Fawn mortality and habitat use among pronghorn during spring and summer in southeastern Oregon, 1981-1982. Wildlife Research Report No. 12. Portland, OR: Oregon Department of Fish and Wildlife, Wildlife Research and Development Division. 117 p. [25692]
635. Trent, James D.; Svejcar, Tony J.; Blank, Robert R. 1994. Mycorrhizal colonization, hyphal lengths, and soil moisture associated with two Artemisia tridentata subspecies. The Great Basin Naturalist. 54(4): 291-300. [25507]
636. TRW Environmental Safety Systems Inc. 1999. Appendix C: Descriptions of land cover types found within or near Yucca Mountain and the potential transportation corridors and facilities. In: Environmental baseline file for biological resources: B00000000-01717-5700-00009 REV 00. Civilian Radioactive Waste Management System: Management and Operating Contractor--Contract Number DE-AC08-91RW00134, [Online]. North Las Vegas, NV: U.S. Department of Energy, Yucca Mountain Site Characterization Office (Producer). Available: http://www.ymp.gov/documents/biology/appendixc.htm [2000, November 6]. [35852]
637. Tueller, Paul T.; Beeson, C. Dwight; Tausch, Robin J.; West, Neil E.; Rea, Kenneth H. 1979. Pinyon-juniper woodlands of the Great Basin: Distribution, flora, vegetal cover. Res. Pap. INT-229. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 22 p. [2367]
638. Tueller, Paul T.; Eckert, Richard E., Jr. 1987. Big sagebrush (Artemisia tridentata vaseyana) and longleaf snowberry (Symphoricarpos oreophilus) plant associations in northeastern Nevada. The Great Basin Naturalist. 47(1): 117-131. [3015]
639. Tuhy, Joel S.; Jensen, Sherman. 1982. Riparian classification for the Upper Salmon/Middle Fork Salmon River drainages, Idaho. Final report. Smithfield, UT: White Horse Associates. Unpublished report on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station Fire Sciences Laboratory, Missoula, MT. 153 p. [8380]
640. Tweit, Susan J.; Houston, Kent E. 1980. Grassland and shrubland habitat types of the Shoshone National Forest. Cody, WY: U.S. Department of Agriculture, Forest Service, Region 2, Shoshone National Forest. 143 p. [2377]
641. U.S. Department of Agriculture, Forest Service, Northern Region. 1973. U.S.D.A. Forest Service environmental statement: Burning for control of big sagebrush. Unpublished draft supplied by Steve Yurich, Regional Forester, U.S. Department of Agriculture, Forest Service, Region 1. 70 p. [2379]
642. Updike, Douglas R.; Loft, Eric R.; Hall, Frank A. 1990. Wildfires on big sagebrush/antelope bitterbrush range in northeastern California: Implications for deer populations. In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., compilers. Proceedings--symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 41-46. [12734]
643. USDA, NRCS. 2018. PLANTS Database, [Online]. U.S. Department of Agriculture, Natural Resources Conservation Service (Producer). Available: https://plants.usda.gov/. [34262]
644. USDA. 2001. Guide to noxious weed prevention practices, [Online]. Washington, DC: U.S. Department of Agriculture, Forest Service (Producer). 25 p. Available online: https://www.fs.usda.gov/invasivespecies/documents/FS_WeedBMP_2001.pdf [2017, January 10] [2009, November 19]. [37889]
645. USDA. 2016. Digital photo series, [Online]. Seattle, WA: Pacific Northwest Research Station, Pacific Wildlife Fire Sciences Laboratory, Fire and Environmental Research Applications Team. Available at: https://depts.washington.edu/nwfire/dps/. [2017, January 10]. [91298]
646. USDA. 2016. Sagebrush natural range of variation, [Online]. In: Pacific Region, Ecology program documents, reports and publications, Natural range of variation of Sierra Nevada habitats. Vallejo, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Region (Producer). Available: https://www.fs.usda.gov/detail/r5/plants-animals/?cid=stelprdb5434436 [2017, January 10]. 36 p. [90944]
647. USDI, Bureau of Land Management. 1993. Fire effects in sagebrush/grass and pinyon-juniper plant communities. In: Fire effects in plant communities on the public lands. EA #MT-930-93-01. Billings, MT: U.S. Department of the Interior, Bureau of Land Management, Montana State Office: I-1 to I-42. [55086]
648. USDI, Fish and Wildlife Service. 2013. Greater sage-grouse (Centrocercus urophasianus) conservation objectives: Final report. Denver, CO: U.S. Department of the Interior, Fish and Wildlife Service. 91 p. [+ appendices]. [90064]
649. Vaitkus, Milda R.; Eddleman, Lee E. 1991. Tree size and understory phytomass production in a western juniper woodland. The Great Basin Naturalist. 51(3): 236-243. [16869]
650. Vale, Thomas R. 1974. Sagebrush conversion projects: An element of contemporary environmental change in the western United States. Biological Conservation. 6(4): 274-284. [91588]
651. Vale, Thomas R. 1975. Presettlement vegetation in the sagebrush-grass area of the Intermountain West. Journal of Range Management. 28(1): 32-36. [2413]
652. Vale, Thomas R. 2002. The pre-European landscape of the United States: Pristine or humanized? In: Vale, Thomas R., ed. Fire, native peoples, and the natural landscape. Washington: Island Press: 1-40. [88507]
653. Van de Water, Kip M.; Safford, Hugh D. 2011. A summary of fire frequency estimates for California vegetation before Euro-American settlement. Fire Ecology. 7(3): 26-58. [85190]
654. Van Dyke, Fred; Darragh, Jeffrey A. 2006. Short- and longer-term effects of fire and herbivory on sagebrush communities in south-central Montana. Environmental Management. 38(3): 365-376. [63225]
655. Van Dyke, Fred; Deboer, Michael J.; Van Beek, Grant M. 1996. Winter range plant production and elk use following prescribed burning. In: Greenlee, Jason, ed. The ecological implications of fire in Greater Yellowstone: Proceedings, 2nd biennial conference on the Greater Yellowstone Ecosystem; 1993 September 19-21; Yellowstone National Park, WY. Fairfield, WA: International Association of Wildland Fire: 193-200. [27855]
656. Van Dyke, Walter A.; Sands, Alan; Yoakum, Jim; Polenz, Allan; Blaisdell, James. 1983. Wildlife habitats in managed rangelands--the Great Basin of southeastern Oregon: bighorn sheep. Gen. Tech. Rep. PNW-159. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest and Range Experiment Station. 37 p. [2417]
657. Veblen, Kari E.; Newingham, Beth A.; Bates, Jon; LaMalfa, Eric; Gicklhorn, Jeff. 2015. Post-fire grazing management in the Great Basin. Great Basin Factsheet series: 7(858), [Online]. Corvallis, OR: Oregon State University (Producer). Available: http://oregonstate.edu/dept/EOARC/sites/default/files/858_gbfs7_post-fire_grazing_2015.pdf [2016, November 9]. 4 p. [91198]
658. Volland, Leonard A.; Dell, John D. 1981. Fire effects on Pacific Northwest forest and range vegetation. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region, Range Management and Aviation and Fire Management. 23 p. [2434]
659. Vosburgh, Timothy; Kramer, Timothy. 2016. Personal communication [Email to Robin Innes]. 19 May. Regarding use of Rx fire in greater sage-grouse habitats within the Lander field office. Lander, WY: U.S. Department of the Interior, Bureau of Land Management, Wind River/Bighorn Basin District, Lander Field Office. Unpublished information on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [90481]
660. Wagle, R. F. 1981. Fire: Its effects on plant succession and wildlife in the Southwest: Some effects of fire on plant succession and variability in the Southwest from a wildlife management viewpoint. Tucson, AZ: University of Arizona. 82 p. [4031]
661. Wagstaff, Fred J.; Welch, Bruce L. 1990. Rejuvenation of mountain big sagebrush on mule deer winter ranges using onsite plants as a seed source. In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., comps. Proceedings--symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 171-174. [12848]
662. Wagstaff, Fred J.; Welch, Bruce L. 1991. Seedstalk production of mountain big sagebrush enhanced through short-term protection from heavy browsing. Journal of Range Management. 44(1): 72-74. [15529]
663. Wakelyn, Leslie A. 1987. Changing habitat conditions on bighorn sheep ranges in Colorado. The Journal of Wildlife Management. 51(4): 904-912. [3001]
664. Walhof, Kendal Scott. 1997. A comparison of burned and unburned big sagebrush communities in southwest Montana. Bozeman, MT: Montana State University. 74 p. Thesis. [46902]
665. Wallestad, Richard. 1975. Life history and habitat requirements of sage grouse in central Montana. Helena, MT: Montana Department of Fish and Game; U.S. Department of the Interior, Bureau of Land Management. 65 p. [25890]
666. Wallestad, Richard; Peterson, Joel G.; Eng, Robert L. 1975. Foods of adult sage grouse in central Montana. The Journal of Wildlife Management. 39(3): 628-630. [2444]
667. Wallmo, Olof C. 1981. Mule and black-tailed deer distribution and habitats. In: Wallmo, Olof C., ed. 1981. Mule and black-tailed deer of North America. Lincoln, NE: University of Nebraska Press: 1-26. [14391]
668. Wallmo, Olof C.; Regelin, Wayne L. 1981. Rocky Mountain and Intermountain habitats: Part 1. Food habits and nutrition. In: Wallmo, Olof C., ed. 1981. Mule and black-tailed deer of North America. Lincoln, NE: University of Nebraska Press: 387-398. [14387]
669. Walser, R. H.; Weber, D. J.; McArthur, E. D.; Sanderson, S. C. 1990. Winter cold hardiness of seven wildland shrubs. In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., compilers. Proceedings--symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 115-118. [12843]
670. Wambolt, C. L.; Creamer, W. H.; Rossi, R. J. 1994. Predicting big sagebrush winter forage by subspecies and browse form class. Journal of Range Management. 47(3): 231-234. [23240]
671. Wambolt, C. L.; Walhof, K. S.; Frisina, M. R. 2001. Recovery of big sagebrush communities after burning in south-western Montana. Journal of Environmental Management. 61(3): 243-252. [40788]
672. Wambolt, Carl L. 1995. Elk and mule deer use of sagebrush for winter forage. Montana Agresearch. Bozeman, MT: Montana Agricultural Experiment Station. 12(2): 35-40. [27101]
673. Wambolt, Carl L. 1996. Mule deer and elk foraging preference for 4 sagebrush taxa. Journal of Range Management. 49(6): 499-503. [27222]
674. Wambolt, Carl L. 1998. Sagebrush and ungulate relationships on Yellowstone's Northern Range. Wildlife Society Bulletin. 26(3): 429-437. [75521]
675. Wambolt, Carl L. 2001. Mule deer foraging preference among five sagebrush (Artemisia L.) taxa. Western North American Naturalist. 61(4): 490-494. [91345]
676. Wambolt, Carl L. 2004. Browsing and plant age relationships to winter protein and fiber of big sagebrush subspecies. Journal of Range Management. 57(6): 620-623. [37475]
677. Wambolt, Carl L. 2005. Sagebrush-ungulate relationships on the Northern Yellowstone Winter Range. In: Shaw, Nancy L.; Pellant, Mike; Monsen, Stephen B., eds. Sage-grouse habitat restoration symposium proceedings; 2001 June 4-7; Boise, ID. Proc. RMRS-P-38. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 109-115. [63190]
678. Wambolt, Carl L.; Frisina, Michael R. 2002. Montana sagebrush: A taxonomic key and habitat descriptions. Intermountain Journal of Sciences. 8(2): 46-59. [47358]
679. Wambolt, Carl L.; Harp, Aaron J.; Welch, Bruce L.; Shaw, Nancy; Connelly, John W.; Reese, Kerry P.; Braun, Clait E.; Klebenow, Donald A.; McArthur, E. Durant; Thompson, James G.; Torell, L. Allen; Tanaka, John A. 2002. Conservation of greater sage-grouse on public lands in the western U.S.: Implications of recovery and management policies. PACWPL Policy Paper SG-02-02. Caldwell, ID: Policy Analysis Center for Western Public Lands. 41 p. [47283]
680. Wambolt, Carl L.; Hoffman, Trista L. 2001. Mountain big sagebrush age distribution and relationships on the northern Yellowstone winter range. In: McArthur, E. Durant; Fairbanks, Daniel J., comps. Shrubland ecosystem genetics and biodiversity: Proceedings; 2000 June 13-15; Provo, UT. Proc. RMRS-P-21. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 330-335. [41992]
681. Wambolt, Carl L.; Hoffman, Trista L.; Mehus, Chris A. 1999. Response of shrubs in big sagebrush habitats to fire on the northern Yellowstone winter range. In: McArthur, E. Durant; Ostler, W. Kent; Wambolt, Carl L., comps. Proceedings: Shrubland ecotones; 1998 August 12-14; Ephraim, UT. Proceedings RMRS-P-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 238-242. [36093]
682. Wambolt, Carl L.; Rens, Reyer J. 2011. Elk and fire impacts on mountain big sagebrush range in Yellowstone. In: Wambolt, Carl L.; Kitchen, Stanley G.; Frisina, Michael R.; Sowell, Bok; Keigley, Richard B.; Palacios, Patsy; Robinson, Jill, comps. Proceedings--shrublands: Wildlands and wildlife habitats; 15th wildland shrub symposium; 2008 June 17-19; Bozeman, MT. Natural Resources and Environmental Issues, Volume XVI. Logan, UT: Utah State University, College of Natural Resources, S. J. and Jessie E. Quinney Natural Resources Research Library: 73-78. [83471]
683. Wambolt, Carl L.; Sherwood, Harrie W. 1999. Sagebrush response to ungulate browsing in Yellowstone. Journal of Range Management. 52(4): 363-369. [91455]
684. Wang, Han; McArthur, E. Durant; Sanderson, Stewart C.; Graham, John H.; Freeman, D. Carl. 1997. Narrow hybrid zone between two subspecies of big sagebrush (Artemisia tridentata: Asteraceae). IV. Reciprocal transplant experiments. Evolution. 51(1): 95-102. [29409]
685. Wansi, Tchouassi; Pieper, Rex D.; Beck, Reldon F.; Murray, Leigh W. 1992. Botanical content of black-tailed jackrabbit diets on semidesert rangeland. The Great Basin Naturalist. 52(4): 300-308. [20177]
686. Warren, Andy. 2018. Personal communication [Email to Robin Innes]. 3 March. Unpublished data and information regarding mountain big sagebrush recovery after prescribed burns. Cheyenne, WY: U.S. Department of the Interior, Bureau of Land Management, National Interagency Fire Center. Documents on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92577]
687. Weber, William A.; Wittmann, Ronald C. 1996. Colorado flora: Eastern slope. 2nd ed. Niwot, CO: University Press of Colorado. 524 p. [27572]
688. Weiner, Nathan I. 2014. The influence of duff distribution in post-fire vegetation recovery patterns in western juniper (Juniperus occidentalis) woodlands in southwestern Idaho. Moscow, ID: University of Idaho. 61 p. Thesis. [88533]
689. Weiss, Nondor T.; Verts, B. J. 1984. Habitat and distribution of pygmy rabbits (Sylvilagus idahoensis) in Oregon. The Great Basin Naturalist. 44(4): 563-571. [23635]
690. Welch, Bruce L. 1989. Nutritive value of shrubs. In: McKell, Cyrus M., ed. The biology and utilization of shrubs. San Diego, CA: Academic Press, Inc.: 405-424. [8041]
691. Welch, Bruce L. 1996. Effects of humidity on storing big sagebrush seed. Res. Pap. INT-RP-493. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 5 p. [30477]
692. Welch, Bruce L. 1997. Seeded versus containerized big sagebrush plants for seed-increase gardens. Journal of Range Management. 50(6): 611-614. [90608]
693. Welch, Bruce L. 2002. Bird counts of burned versus unburned big sagebrush sites. Res. Note RMRS-RN-16. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 6 p. [35562]
694. Welch, Bruce L. 2005. Big sagebrush chemistry and water relations. In: Welch, Bruce L., ed. Big sagebrush: A sea fragmented into lakes, ponds, and puddles. Gen. Tech. Rep. RMRS-GTR-144. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 107-148. [55362]
695. Welch, Bruce L. 2005. Big sagebrush genetics. In: Welch, Bruce L., ed. Big sagebrush: A sea fragmented into lakes, ponds, and puddles. Gen. Tech. Rep. RMRS-GTR-144. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 149-170. [55358]
696. Welch, Bruce L. 2005. Big sagebrush: A sea fragmented into lakes, ponds, and puddles. Gen. Tech. Rep. RMRS-GTR-144. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 210 p. [55356]
697. Welch, Bruce L. 2005. Birds, mammals, and reptiles associated with big sagebrush. In: Welch, Bruce L., ed. Big sagebrush: A sea fragmented into lakes, ponds, and puddles. Gen. Tech. Rep. RMRS-GTR-144. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 47-80. [55399]
698. Welch, Bruce L. 2005. Consequences of controlling big sagebrush. In: Welch, Bruce L., ed. Big sagebrush: A sea fragmented into lakes, ponds, and puddles. Gen. Tech. Rep. RMRS-GTR-144. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 187-204. [55360]
699. Welch, Bruce L. 2005. Getting acquainted with big sagebrush. In: Welch, Bruce L., ed. Big sagebrush: A sea fragmented into lakes, ponds, and puddles. Gen. Tech. Rep. RMRS-GTR-144. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 1-46. [55357]
700. Welch, Bruce L. 2005. Other foragers and winter damage on big sagebrush. In: Welch, Bruce L., ed. Big sagebrush: A sea fragmented into lakes, ponds, and puddles. Gen. Tech. Rep. RMRS-GTR-144. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 81-106. [55361]
701. Welch, Bruce L. 2005. Putting big sagebrush back into its ecosystem. In: Welch, Bruce L., ed. Big sagebrush: A sea fragmented into lakes, ponds, and puddles. Gen. Tech. Rep. RMRS-GTR-144. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 171-186. [55359]
702. Welch, Bruce L.; Briggs, Steven F.; Johansen, James H. 1996. Big sagebrush seed storage. Res. Note INT-RN-430. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 3 p. [30478]
703. Welch, Bruce L.; Criddle, Craig. 2003. Countering misinformation concerning big sagebrush. Res. Pap. RMRS-RP-40. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 28 p. [47361]
704. Welch, Bruce L.; Jacobson, Tracy L. C. 1988. Root growth of Artemisia tridentata. Journal of Range Management. 41(4): 332-334. [4559]
705. Welch, Bruce L.; McArthur, E. Durant. 1979. Feasibility of improving big sagebrush (Artemisia tridentata) for use on mule deer winter ranges. In: Goodin, J. R.; Northington, David K., eds. Arid land plant resources: [Date and location of symposium unknown]. Lubbock, TX: International Center for Arid and Semi-Arid Land Studies: 451-473. [2482]
706. Welch, Bruce L.; McArthur, E. Durant. 1981. Variation of monoterpenoid content among subspecies and accessions of Artemisia tridentata grown in a uniform garden. Journal of Range Management. 34(5): 380-384. [91418]
707. Welch, Bruce L.; McArthur, E. Durant. 1986. Growth rate of big sagebrush as influenced by accessions, sites, subspecies, and years. In: McArthur, E. Durant; Welch, Bruce L., compilers. Proceedings--symposium on the biology of Artemisia and Chrysothamnus; 1984 July 9-13; Provo, UT. Gen. Tech. Rep. INT-200. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 342-346. [2483]
708. Welch, Bruce L.; McArthur, E. Durant. 1986. Wintering mule deer preference for 21 accessions of big sagebrush. The Great Basin Naturalist. 46(2): 281-286. [2484]
709. Welch, Bruce L.; McArthur, E. Durant; Davis, James N. 1981. Differential preference of wintering mule deer for accessions of big sagebrush and for black sagebrush. Journal of Range Management. 34(5): 409-411. [7801]
710. Welch, Bruce L.; McArthur, E. Durant; Nelson, David L.; Pederson, Jordan C.; Davis, James N. 1986. `Hobble Creek'--a superior selection of low-elevation mountain big sagebrush. Res. Pap. INT-370. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 10 p. [2485]
711. Welch, Bruce L.; Pederson, Jordan C. 1981. In vitro digestibility among accessions of big sagebrush by wild mule deer and its relationship to monoterpenoid content. Journal of Range Management. 34(6): 497-500. [91350]
712. Welch, Bruce L.; Wagstaff, Fred J. 1992. 'Hobble Creek' big sagebrush vs. antelope bitterbrush as a winter forage. Journal of Range Management. 45(2): 140-142. [18250]
713. Welch, Bruce L.; Wagstaff, Fred J.; Jorgensen, Gary L. 1990. `Hobble Creek' mountain big sagebrush seed production. In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., comps. Proceedings--symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 167-174. [12847]
714. Welch, Bruce L.; Wagstaff, Fred J.; Roberson, Jay A. 1991. Preference of wintering sage grouse for big sagebrush. Journal of Range Management. 44(5): 462-465. [16608]
715. Weldon, L. W.; Bohmont, D. W.; Alley, H. P. 1959. The interrelation of three environmental factors affecting germination of sagebrush seed. Journal of Range Management. 12: 236-238. [2489]
716. Welsh, Stanley L.; Atwood, N. Duane; Goodrich, Sherel; Higgins, Larry C., eds. 1987. A Utah flora. The Great Basin Naturalist Memoir No. 9. Provo, UT: Brigham Young University. 894 p. [2944]
717. Weltz, Mark A.; Spaeth, Ken; Taylor, Michael H.; Rollins, Kimberly; Pierson, Fred; Jolley, Leonard; Nearing, Mark; Goodrich, Dave; Hernandez, Mariano; Nouwakpo, Sayjro K.; Rossi, Colleen. 2014. Cheatgrass invasion and woody species encroachment in the Great Basin: Benefits of conservation. Journal of Soil and Water Conservation. 69(2): 39-44. [88918]
718. Weppner, Kerrie N.; Pierce, Jennifer L.; Betancourt, Julio L. 2013. Holocene fire occurrence and alluvial responses at the leading edge of pinyon-juniper migration in the Northern Great Basin, USA. Quaternary Research. 80(2): 143-157. [88833]
719. West, Neil E. 1983. Great Basin-Colorado Plateau sagebrush semi-desert. In: Goodall, David W., ed. Temperate deserts and semi-deserts. Ecosystems of the world: Vol. 5. Amsterdam; Oxford; New York: Elsevier Scientific Publishing Company: 331-349. [2505]
720. West, Neil E. 1988. Intermountain deserts, shrub steppes, and woodlands. In: Barbour, Michael G.; Billings, William Dwight, eds. North American terrestrial vegetation. New York: Cambridge University Press: 209-230. [19546]
721. West, Neil E. 2000. Synecology and disturbance regimes of sagebrush steppe ecosystems. In: Entwistle, P. G.; DeBolt, A. M.; Kaltenecker, J. H.; Steenhof, K., comps. Sagebrush steppe ecosystems symposium: Proceedings; 1999 June 21-23; Boise, ID. Publ. No. BLM/ID/PT-001001+1150. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Boise State Office: 15-26. [42712]
722. West, Neil E.; Tausch, Robin J.; Rea, Kenneth H.; Tueller, Paul T. 1978. Taxonomic determination, distribution, and ecological indicator values of sagebrush within the pinyon-juniper woodlands of the Great Basin. Journal of Range Management. 31(2): 87-92. [2521]
723. Western Association of Fish and Wildlife Agencies. 2009. Prescribed fire as a management tool in xeric sagebrush ecosystems: Is it worth the risk to sage-grouse? [Online]. Boise, ID: Western Association of Fish and Wildlife Agencies, Sage and Columbian sharp-tailed grouse technical committee (producer). 22 pages. Available: https://wgfd.wyo.gov/WGFD/media/content/PDF/Habitat/Sage%20Grouse/SG_PRESCRIBEDBURNING.pdf [2017, April 19]. [91674]
724. Whisenant, Steven G. 1990. Changing fire frequencies on Idaho's Snake River Plains: Ecological and management implications. In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., comps. Proceedings--symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 4-10. [12729]
725. Whisenant, Steven G. 2004. Vegetative manipulation with prescribed burning. In: Monsen, Stephen B.; Stevens, Richard; Shaw, Nancy L., comps. Restoring western ranges and wildlands. Gen. Tech. Rep. RMRS-GTR-136-vol-1. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 101-120. [52824]
726. White, Susan M.; Flinders, Jerran T.; Welch, Bruce L. 1982. Preference of pygmy rabbits (Brachylagus idahoensis) for various populations of big sagebrush (Artemisia tridentata). Journal of Range Management. 35(6): 724-726. [23636]
727. Whitlock, Cathy; Knox, Margaret A. 2002. Prehistoric burning in the Pacific Northwest: Human versus climatic influences. In: Vale, Thomas R., ed. Fire, native peoples, and the natural landscape. Washington: Island Press: 195-269. [88512]
728. Whitlock, Cathy; Shafer, Sarah L.; Marlon, Jennifer. 2003. The role of climate and vegetation change in shaping past and future fire regimes in the northwestern US and the implications for ecosystem management. In: Young, Michael K.; Gresswell, Robert E.; Luce, Charles H., guest eds. Selected papers from an international symposium on effects of wildland fire on aquatic ecosystems in the western USA; 2002 April 22-24; Boise, ID. In: Forest Ecology and Management. Special Issue: The effects of wildland fire on aquatic ecosystems in the western USA. New York: Elsevier Science; 178(1-2): 5-21. [44921]
729. Wijayratne, Upekala C.; Pyke, David A. 2012. Burial increases seed longevity of two Artemisia tridentata (Asteraceae) subspecies. American Journal of Botany. 99(3): 438-447. [86456]
730. Williams, Gerald W. 2004. American Indian fire use in the arid west. Fire Management Today. 64(3): 10-14. [50486]
731. Williams, Mary I.; Thurow, Thomas L.; Paige, Ginger B.; Hild, Ann L.; Gerow, Kenneth G. 2011. Sagebrush-obligate passerine response to ecological site characteristics. In: Wambolt, Carl L.; Kitchen, Stanley G.; Frisina, Michael R.; Sowell, Bok; Keigley, Richard B.; Palacios, Patsy; Robinson, Jill, comps. Proceedings--shrublands: Wildlands and wildlife habitats; 15th wildland shrub symposium; 2008 June 17-19; Bozeman, MT. Natural Resources and Environmental Issues, Volume XVI. Logan, UT: Utah State University, College of Natural Resources, S. J. and Jessie E. Quinney Natural Resources Research Library: 63-71. [83470]
732. Willis, Mitchell J.; Miller, Richard F. 1999. Importance of western juniper communities to small mammals. In: Monsen, Stephen B.; Stevens, Richard, compilers. Proceedings: Ecology and management of pinyon-juniper communities within the Interior West: Sustaining and restoring a diverse ecosystem; 1997 September 15-18; Provo, UT. Proceedings RMRS-P-9. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 210-214. [30554]
733. Wilt, F. Martin; Geddes, Jason D.; Tamma, Rama V.; Miller, Glenn C.; Everett, Richard L. 1992. Interspecific variation of phenolic concentrations in persistent leaves among six taxa from subgenus Tridentatae of Artemisia (Asteraceae). Biochemical Systematics and Ecology. 20(1): 41-52. [34701]
734. Winward, A. H.; Tisdale, E. W. 1977. Taxonomy of the Artemisia tridentata complex in Idaho. Bulletin Number 19. Moscow, ID: University of Idaho, College of Forestry, Wildlife and Range Sciences, Forest, Wildlife and Range Experiment Station. 15 p. [2590]
735. Winward, Al H. 1991. A renewed commitment to management of sagebrush grasslands. In: Miller, R. F., ed. Management in the sagebrush steppe. Special Report 880. Corvallis, OR: Oregon State University, Agricultural Experiment Station: 2-7. [35554]
736. Winward, Alma H. 1970. Taxonomic and ecological relationships of the big sagebrush complex in Idaho. Moscow, ID: University of Idaho. 79 p. Dissertation. [2583]
737. Winward, Alma H. 1980. Taxonomy and ecology of sagebrush in Oregon. Station Bulletin 642. Corvallis, OR: Oregon State University, Agricultural Experiment Station. 15 p. [2585]
738. Winward, Alma H. 1985. Fire in the sagebrush-grass ecosystem--the ecological setting. In: Saunders, Ken; Durham, Jack; [and others], eds. Rangeland fire effects: Proceedings of the symposium; 1984 November 27-29; Boise, ID. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office: 2-6. [2587]
739. Wisdom, Michael J.; Chambers, Jeanne C. 2009. A landscape approach for ecologically based management of Great Basin shrublands. Restoration Ecology. 17(5): 740-749. [82738]
740. Woodard, Paul M.; Van Nest, Terry. 1990. Winter burning bighorn sheep range--a proposed strategy. Forestry Chronicle. October: 473-477. [14619]
741. Wright, Clinton S. 2013. Models for predicting fuel consumption in sagebrush-dominated ecosystems. Rangeland Ecology and Management. 66(3): 254-266. [87795]
742. Wright, Clinton S.; Prichard, Susan J. 2006. Biomass consumption during prescribed fires in big sagebrush ecosystems. In: Andrews, Patricia L.; Butler, Bret W., comps. Fuels management--how to measure success: Conference proceedings; 2006 March 28-30; Portland, OR. Proceedings RMRS-P-41. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 489-500. [65176]
743. Wright, Clinton S.; Vihnanek, Robert E.; Restaino, Joseph C.; Dvorak, Jon E. 2012. Photo series for quantifying natural fuels. Volume XI: Eastern Oregon sagebrush-steppe and spotted owl nesting habitat in the Pacific Northwest. Gen. Tech. Rep. PNW-GTR-878. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 85 p. [87939]
744. Wright, Henry A.; Bailey, Arthur W. 1982. Fire ecology: United States and southern Canada. New York: John Wiley & Sons. 501 p. [2620]
745. Wright, Henry A.; Neuenschwander, Leon F.; Britton, Carlton M. 1979. The role and use of fire in sagebrush-grass and pinyon-juniper plant communities: A state-of-the-art review. Gen. Tech. Rep. INT-58. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 48 p. [2625]
746. Wright. Clinton Stewart. 2010. Effects of disturbance and fuelbed succession on spatial patterns of fuel, fire hazard, and carbon; and fuel consumption in shrub-dominated ecosystems. Seattle, WA: University of Washington. 152 p. [+ appendices]. Dissertation. [91204]
747. Wrobleski, David W.; Kauffman, J. Boone. 2003. Initial effects of prescribed fire on morphology, abundance, and phenology of forbs in big sagebrush communities in southeastern Oregon. Restoration Ecology. 11(1): 82-90. [47380]
748. Wyoming Game and Fish Department. 2010. Sagebrush shrublands, [Online]. Cheyenne, WY: Wyoming Game and Fish Department (Producer). Available: https://wgfd.wyo.gov/Habitat/Habitat-Plans/Wyoming-State-Wildlife-Action-Plan/Terrestrial-Habitat-Types [2016, December 2]. [91222]
749. Wyoming Interagency Vegetation Committee. 2002. Wyoming guidelines for managing sagebrush communities with emphasis on fire management. Cheyenne, WY: U.S. Department of the Interior, Bureau of Land Management, Wyoming Game and Fish Department. 53 p. [90582]
750. Yanish, Curtis R. 2002. Western juniper succession: Changing fuels and fire behavior. Moscow, ID: University of Idaho. 85 p. Thesis. [47382]
751. Yanish, Curtis R.; Bunting, Stephen C.; Kingery, James L. 2005. Western juniper (Juniperus occidentalis) succession: Changing fuels and fire behavior. In: Sagebrush steppe and pinyon-juniper ecosystems: Effects of changing fire regimes, increased fuel loads, and invasive species. Final Report to the Joint Fire Science Program: Project Number 00-1-1-03. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 16-24. [64234]
752. Yoakum, James D. 1979. Managing rangelands for pronghorns. Rangelands. 1(4): 146-148. [25689]
753. Yoakum, Jim D. 2000. An assessment of wildfires and pioneering pronghorn in Long Valley, California/Nevada. In: Fischer, Jon K., ed. Proceedings of the 17th biennial pronghorn antelope workshop; 1996 June 4-7; Lake Tahoe, CA. Sacramento, CA: California Department of Fish and Game: 86-90. [91498]
754. Yoakum, Jim. 1980. Habitat management guides for the American pronghorn antelope. Tech. Note 347. Denver, CO: U.S. Department of the Interior, Bureau of Land Management, Denver Service Center. 77 p. [23170]
755. Yoakum, Jim. 1983. Managing vegetation for pronghorns in the Great Basin. In: Monsen, Stephen B.; Shaw, Nancy, comps. Managing Intermountain rangelands - Improvement of range and wildlife habitats: Proceedings; 1981 Sept 15-17; Twin Falls, ID; 1982 June 22- 24; Elko, NV. Gen. Tech. Rept. INT-157. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 189-193. [2637]
756. Young, James A.; Allen, Fay L. 1997. Cheatgrass and range science: 1930-1950. Journal of Range Management. 50(5): 530-535. [30455]
757. Young, James A.; Clements, Charlie D.; Jansen, Henricus. 2007. Sagebrush steppe. In: Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. Terrestrial vegetation of California. Berkeley, CA: University of California Press: 587-608. [82714]
758. Young, James A.; Evans, Raymond A. 1978. Population dynamics after wildfires in sagebrush grasslands. Journal of Range Management. 31(4): 283-289. [2657]
759. Young, James A.; Evans, Raymond A. 1981. Demography and fire history of a western juniper stand. Journal of Range Management. 34(6): 501-505. [2659]
760. Young, James A.; Evans, Raymond A. 1989. Dispersal and germination of big sagebrush (Artemisia tridentata) seeds. Weed Science. 37(2): 201-206. [7235]
761. Young, James A.; Evans, Raymond A. 1989. Reciprocal common garden studies of the germination of seeds of Big Sagebrush (Artemisia tridentata). Weed Science. 37: 319-325. [8692]
762. Young, James A.; Evans, Raymond A.; Eckert, Richard E., Jr.; Kay, Burgess L. 1987. Cheatgrass. Rangelands. 9(6): 266-270. [288]
763. Young, James A.; Evans, Raymond A.; Palmquist, Debra E. 1989. Big sagebrush (Artemisia tridentata) seed production. Weed Science. 37: 47-53. [6676]
764. Young, James A.; Palmquist, Debra E.; Evans, Raymond A. 1991. Temperature profiles for germination of big sagebrush seeds from native stands. Journal of Range Management. 44(4): 385-390. [15480]
765. Young, Kert R.; Roundy, Bruce A.; Bunting, Stephen C.; Eggett, Dennis L. 2015. Utah juniper and two-needle pinon reduction alters fuel loads. International Journal of Wildland Fire. 24(2): 236-248. [89970]
766. Youngblood, Andrew P.; Mauk, Ronald L. 1985. Coniferous forest habitat types of central and southern Utah. Gen. Tech. Rep. INT-187. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 89 p. [2684]
767. Ziegenhagen, Lori L. 2003. Shrub reestablishment following fire in the mountain big sagebrush (Artemisia tridentata Nutt. spp. vaseyana (Rydb.) Beetle) alliance. Corvallis, OR: Oregon State University. 102 p. [+ appendices]. Thesis. [90185]
768. Ziegenhagen, Lori L.; Miller, Richard F. 2009. Postfire recovery of two shrubs in the interiors of large burns in the Intermountain West, USA. Western North American Naturalist. 69(2): 195-205. [81840]