| FEIS Home Page |
|
 |
| Big Bend National Park. ®Photo courtesy of James Manhart. |
Revisions:
On 4 December 2015, additional varieties of ponderosa pine were listed in the Taxonomy section, and citations supporting the updated taxonomy [177,295] were added. In keeping with this taxonomy, this Species Review was expanded to include both southwestern (Pinus ponderosa var. brachyptera) and Rocky Mountain (Pinus ponderosa var. scopulorum) ponderosa pines.
Callaham [86] also recognizes a sixth entity, Pinus ponderosa subsp. readiana Callaham, called central high plains ponderosa pine. Arizona pine (P. arizonica Englm.) is closely related to ponderosa pine but is treated as a separate species [66,85]. Ponderosa pine populations that extend northward into western Texas from Coahuila are classified as either P. p. var. scopulorum [116] or P. arizonica var. stormiae Mart. [173,177,253]. Information on ponderosa pine in Texas and east of the Continental Divide in Montana, Wyoming, and South Dakota is included in this review.
Follow the links above to FEIS reviews of other ponderosa pine varieties.
Terms used in this review: Many taxonomists did not distinguish between southwestern and Rocky Mountain ponderosa pine until the
early 2010s. Because most literature does not distinguish between the 2 varieties, this review uses "ponderosa pine" to refer to both Rocky Mountain and southwestern ponderosa pines.
Hybrids: Hybridization and introgression occur between varieties of ponderosa pine and Arizona pine.
Ponderosa × Arizona pine hybrids may bear 3- and 4-needle fascicles; 4- and 5-needle
fascicles; or 3-, 4-, and 5-needle fascicles, sometimes on the same spur branch
[38,97,116,236]. Ponderosa and Arizona pines also hybridize and introgress with Apache
pine (P. engelmannii); 3-taxa hybrids (ponderosa × Arizona × Apache
pines) occur occasionally [236].
SYNONYMS:
For southwestern ponderosa pine:
Pinus ponderosa subsp. brachyptera (Engelm.) Silba (documented in 295])
For Rocky Mountain ponderosa pine:
Pinus ponderosa subsp. scopulorum (Watson) Weber [309]
Pinus arizonica var. stormiae Mart., pino real [173,253]
LIFE FORM:
Tree
FEDERAL LEGAL STATUS:
No special status
OTHER STATUS:
No entry
GENERAL DISTRIBUTION:
Ponderosa pine is the most widely distributed pine species in North America,
ranging north-south from southern British Columbia to central Mexico and east-west
from central Nebraska to the west coast [202]. Ponderosa pine
ecosystems occupy about 38 million acres across 14 states [122]. The U.S. Geological Survey provides a distributional
map of ponderosa pine.
Distributions of Rocky Mountain and southwestern ponderosa pine are:
 |
 |
|
| Southwestern ponderosa pine (left) and Rocky Mountain ponderosa pine (right) distributions. Maps from PLANTS: Maps courtesy of USDA, NRCS. 2015. The PLANTS Database. National Plant Data Team, Greensboro, NC. (3 December 2015) [295]. | ||
Distribution of southwestern ponderosa pine continues south from Texas into Coahuila, Tamaulipas, and San Luis Potos, Mexico [81,82,274]. Varieties of ponderosa pine are not geographically distinct taxa [163]. There is a broad zone of intergradation between Rocky Mountain and Pacific ponderosa pines in southeastern Idaho and on both sides of the Continental Divide in Montana and Wyoming [38,73,97,235,236]. Ponderosa pine × Arizona pine intergrades occur in southeastern Arizona, southeastern New Mexico, and Mexico [38,97,116,236]. On sky islands of southeastern Arizona, ponderosa pine occurs mostly above 7,000 feet (2,100 m) elevation. A transition zone of intergraded ponderosa pine × Arizona pine occurs between 5,000 and 7,000 feet (1,500-2,100 m) [97].
ECOSYSTEMS [122]:| AZ | CO | KS | MT | NE | NV | NM | ND | SD | TX | UT | WY |
| MEXICO |
MT: In central Montana the Rocky Mountain ponderosa pine type merges into plains grassland at lower elevations and limber pine at higher elevations. Limber pine, Rocky Mountain Douglas-fir, and Rocky Mountain juniper (Juniperus scopulorum) are common overstory associates. Plains grassland species include blue grama (Bouteloua gracilis), sideoats grama (B. curtipendula), and prairie Junegrass (Koeleria macrantha). Western snowberry (Symphoricarpos occidentalis) and skunkbush sumac (Rhus trilobata) are common shrub associates. Soapweed yucca (Yucca glauca), pygmy prickly-pear (Opuntia fragilis), and plains prickly-pear (O. polyacantha) occur on the driest sites [16,230]. Rocky Mountain juniper, bluebunch wheatgrass (Pseudoroegneria spicata), and longleaf wormwood (Artemisia longifolia) are dominant species in Rocky Mountain ponderosa pine of the Missouri River Breaks [208].
Rocky Mountain ponderosa pine is the only forest tree in southeastern Montana, where it forms several diverse habitats. On dry sites it supports an understory of plains grassland species such as big bluestem (Andropogon gerardii var. gerardii), little bluestem (Schizachyrium scoparium), and blue grama, and of mountain grassland species such as bluebunch wheatgrass and elk sedge (Carex geyeri). Understories are typically dense on wetter sites, and include species characteristic of Columbia ponderosa pine forests to the west. Russet buffaloberry (Shepherdia canadensis), bearberry (Arctostaphylos uva-ursi), twinflower (Linnaea borealis), and heartleaf arnica (Arnica cordifolia) are common moist-site associates [16,230].
WY: Rocky Mountain ponderosa pine is usually the only tree at lower timberline in the Bighorn Mountains. Shrubs or grasses may dominate the understory. The most common understory dominants are bluebunch wheatgrass, Idaho fescue, white spirea (Spiraea betulifolia), and mountain ninebark (Physocarpus monogynus) [164]. Rocky Mountain ponderosa pine communities are uncommon in southeastern Wyoming. Where they occur, elk sedge most commonly dominates the understory. Ross' sedge (C. rossii) and bearberry are occasional understory dominants. Common graminoids and forbs include prairie Junegrass, spike fescue (Leucopoa kingii), western yarrow (Achillea millefolium), and heartleaf arnica [8].
White spruce (Picea glauca) may cooccur with Rocky Mountain ponderosa pine in the Black Hills of Wyoming and South Dakota. Bur oak (Quercus macrocarpa), quaking aspen (Populus tremuloides), and paper birch (Betula papyrifera) are overstory associates [52].
Dakotas: Rocky Mountain ponderosa pine understories in the Black Hills of South Dakota are usually dominated by shrubs including common juniper (Juniperus communis), russet buffaloberry, common snowberry (Symphoricarpos albus), chokecherry (Prunus virginiana), and bur oak. A few stands have grassy understories of timber oatgrass (Danthonia intermedia), Kentucky bluegrass (Poa pratensis), and/or little bluestem [289]. Rocky Mountain ponderosa pine forests and woodlands are not abundant in North Dakota. Understory composition is similar to that of Rocky Mountain ponderosa pine in the Black Hills [125,138].
CO: On the Colorado Front Range, Rocky Mountain ponderosa pine generally occurs with Rocky Mountain Douglas-fir on north slopes and Rocky Mountain juniper on south slopes [215]. Quaking aspen commonly cooccurs on sites that have experienced past fires or other reoccurring disturbance [84]. Understory species include common juniper, wax currant (Ribes cereum), bearberry, spike fescue, blue grama, and buffalo grass (Buchloe dactyloides). At the forest-plains grassland ecotone, understory species may also include black grama (Bouteloua eriopoda), little bluestem, needlegrasses (Hesperostipa and Achnatherum spp.), western wheatgrass (Pascopyrum smithii), and cheatgrass (Bromus tectorum) [215]. Rocky Mountain ponderosa pine is a minor species in western Colorado, where it occurs in Rocky Mountain Douglas-fir forests [165].
In southern Colorado, interior ponderosa pine lies between lower-elevation grassland or pinyon-juniper (Pinus-Juniperus spp.) and higher-elevation Rocky Mountain Douglas-fir. Grassy understories with blue grama, mountain muhly (Muhlenbergia montana), mutton grass (Poa fendleriana), Arizona fescue, and little bluestem are common on dry sites. On moister sites, shrubs such as Gambel oak (Quercus gambelii), wavyleaf oak (Q. × pauciloba), and bearberry may also be important [91].
UT: Ponderosa pine/shrub communities in central and southern Utah are usually the lowest coniferous forest type, and border shrublands or Colorado pinyon-Utah juniper (Pinus edulis-Juniperus osteosperma) woodland. Dominant understory species include curlleaf mountain-mahogany (Cercocarpus ledifolius), greenleaf manzanita (Arctostaphylos patula), black sagebrush (Artemisia nova), Gambel oak, and mountain snowberry (S. oreophilus). Ponderosa pine/mountain muhly occurs in central and southern Utah [322]. Interior ponderosa pine/grassland series also occur in northeastern Utah: dominant grasses are Idaho fescue, sheep fescue (F. ovina), and/or mutton grass. Rocky Mountain lodgepole pine, Utah juniper, and quaking aspen are seral in these habitat types [216].
Ponderosa pine in southeastern Nevada occurs mostly on northerly aspects of canyons. Associated species include Utah juniper, Gambel oak, rubber rabbitbrush (Chrysothamnus nauseosus), big sagebrush (Artemisia tridentata), bottlebrush squirreltail (Elymus elymoides), western wheatgrass, and cheatgrass [45].
Southwest: Ponderosa pine is the most common forest tree in the Southwest [323]. Vegetation patterns in southern New Mexico and Arizona are mostly determined by aspect and elevation. In southeastern Arizona, ponderosa pine forest lies on elevational and moisture gradients between dry, low-elevation Mexican pinyon-Chihuahua pine (Pinus cembroides-P. leiophylla var. chihuahuana)-Apache pine forest and/or Madrean oak woodland and moister, higher-elevation southwestern white pine (P. strobiformis)-Rocky Mountain Douglas-fir forest [30,35,135]. At 7,500 to 8,500 feet (2,300 to 2,600 m) elevation, ponderosa pine, Arizona pine, and encinal oaks tend to dominate dry, south- and west-facing slopes, while Rocky Mountain Douglas-fir and corkbark fir (Abies lasiocarpa var. arizonica) occupy north- and east-facing slopes. Madrean oak associates include Arizona white oak (Quercus arizonica), Gambel oak, Emory oak (Q. emoryi), and silverleaf oak (Q. hypoleucoides). Understories are dominated by grasses including pine dropseed (Blepharoneuron tricholepis), Arizona wheatgrass (Elymus arizonicus), prairie Junegrass, and screwleaf muhly (M. virescens) [30]. In high-elevation (8,400 to 9,300 feet (2,500-2,800 m)) mixed-conifer forest, Engelmann spruce, blue spruce (P. pungens), corkbark fir, Rocky Mountain Douglas-fir, southwestern white pine, and quaking aspen cooccur with ponderosa pine [127].
AZ: Associates in ponderosa pine/bunchgrass in northern Arizona include mountain muhly, mutton grass, Arizona fescue, bottlebrush squirreltail, blue grama, pine dropseed, deergrass (M. rigens), and prairie Junegrass [25]. Shrubs may be nearly absent to fairly common. If present, they are usually widely and irregularly spaced compared to more northern ponderosa pine communities. Fendler ceanothus (Ceanothus fendleri), wax currant, skunkbush sumac, greenleaf manzanita, Stansbury cliffrose (Purshia mexicana var. stansburiana), Apache plume (Fallugia paradoxa), and mountain-mahogany (Cercocarpus spp.) are among the most common shrubs in ponderosa pine/bunchgrass. The ponderosa pine type usually merges into Arizona chaparral at lower elevations of northern and central Arizona. It may also finger into riparian zones [60]. At higher elevations, the type merges into corkbark fir-spruce (Picea spp.) forest. Associates in the upper ponderosa pine zone include those shrubs also found in fir-spruce forest: common juniper, Oregon boxwood (Paxistima myrsinites), red raspberry (Rubus idaeus), mountain ninebark, and snowberry (Symphoricarpos spp.). Rocky Mountain Douglas-fir and quaking aspen may cooccur with ponderosa pine above 7,000 feet (2,000 m) [68,77,229].
TX: Netleaf oak (Q. rugosa), silverleaf oak, and quaking aspen are common associates throughout western Texas. Colorado pinyon and alligator juniper (Juniperus deppeana) often form a woodland association with ponderosa pine on south slopes; Douglas-fir and southwestern white pine join ponderosa pine in a forest/bunchgrass association on moister slopes [2]. Understory associates include finestem tussockgrass (Nassella tenuissima), California brome (Bromus carinatus), Big Bend bluegrass (Poa strictiramea), bulb panicgrass (Panicum bulbosum), and pinyon ricegrass (Piptochaetium fimbriatum) [90]. In the Guadalupe and Chisos mountains, ponderosa pine dominates ponderosa pine-Douglas-fir-southwestern white pine parklands and is an associate in Chisos red oak (Q. gravesii)-Colorado pinyon woodlands. In the Davis Mountains, it occurs in ponderosa pine-southwestern white pine-Gambel oak associations [2,90,92,267].
Vegetation classifications describing plant communities dominated by Rocky Mountain or southwestern ponderosa pine are:
AZ [36,114,199,223,226,277,313]Rocky Mountain ponderosa pine has mostly 2-needle fascicles [16,73,310]. The variety's scientific name (scopulorum) derives from the stiff, tufted (scopulate) foliage and compact crown characteristic of that variety [73]. Southwestern ponderosa pine has mostly 3-needle fascicles and a relatively open crown [179,310].
Structure: Stands were historically open with varied age class distributions [213,215]. Cooper [75] found that on the Fort Apache Reservation of east-central Arizona, where frequent prescribed burning is practiced, ponderosa pine grew in a mosaic of small (about 0.2-acre (0.08 ha)), even-aged groups that were maintained by fire. Trees were widely and randomly spaced. In contrast, structure in an unburned virgin stand on the Fort Valley Experimental Forest was uneven-aged with trees in small, uneven-aged groups. Pole-sized trees predominated, with scattered presettlement trees 14.5 to 41 inches (37-104 cm) DBH, and dense thickets of saplings [77]. (The Fire Ecology section of this report compares of pre- and postsettlement structure of this stand.) Laven and others [198] found presettlement stands on the central Colorado Front Range tended to have widely spaced, uneven-aged trees. Grouping patterns were not evident [77].
Age: Ponderosa pine is long lived. Larger trees may attain ages of 700 or more years [192,262], although ponderosa pine between 350 and 425 years of age has a high mortality rate [221]. In a reconstruction of presettlement stand age classes near Flagstaff, Mast and others [213] found that prior to 1876, trees greater than 100 years old made up 68% of the overstory. The oldest Rocky Mountain ponderosa pine on record was a 1,047-year-old tree from Colorado [192]. In the Southwest, Swetnam and Brown [285] recorded a 742-year-old tree from northwestern Arizona.
Physiology: The tough, compact foliage of Rocky Mountain ponderosa pine probably confers cold tolerance [73]. A deep, extensive root system, high sapwood:heartwood ratio, and sunken stomata make ponderosa pine highly drought tolerant throughout its range [36,73]. In Rocky Mountain ponderosa pine's distribution, only limber pine and Rocky Mountain bristlecone pine (Pinus aristida) better withstand extended drought [36]. In the Southwest, Barton and Teeri [35] found that ponderosa pine seedlings from the Chiricahua Mountains of southeastern Arizona were more drought resistant than southwestern white pine but less so than Mexican pinyon, Chihuahua, and Apache pines.
RAUNKIAER [239] LIFE FORM:Seeds are mostly wind dispersed and do not usually carry more than 120 feet (37 m) from the parent tree [227]. Seeds cached by rodents may result in some seedling establishment. Clark's nutcrackers play a minor but important role in seed dispersal because seeds they cache are more likely to establish than rodent-cached seeds [194].
Seeds require mineral soil and do not germinate until the soil is continuously warm and moist [195]. There is some evidence that unlike Pacific ponderosa pine, Rocky Mountain ponderosa pine does not require cold/damp stratification prior to germination (M. G. Harrington 2003, pers. comm.). Schmid and Mitchell [259] found the number of sound seeds collected from trees on the Coconino and Kaibab national forests did not vary significantly between sites (P=0.05), but did vary significantly among trees. Mean number of sound seeds/cone/tree ranged from 35 to 58. In field trials, germination rate of seed collected and outplanted on the Fort Valley Experimental Forest showed 61 to 90% germination, with germination time varying from 12 to 37 days. First-year survivorship was greatest (13%-19%) in seedlings that germinated in early to mid-July, and least (2%-7%) in seedlings that germinated in August. Predation of germinants by dark-eyed juncos was responsible for over half of germinant mortality [195]. Seed germination is usually lowest, and seed predation highest, in years when seed crops are low [182]. Ponderosa pine is not a seed-banking species, although a minor amount of seed may germinate in its 2nd spring or summer [172].
Seedling establishment: Seedlings are relatively shade intolerant and require canopy-opening disturbances such as fire, logging, or tree death to establish [34,192,214,271]. Establishment pulses may occur on open sites when wet years follow a fire year, but wet postfire years are not always required for good establishment in the central Rocky Mountains and north. Two major pulses of seedling establishment in Rocky Mountain National Park, Colorado (1870-1890 and 1970-1980), occurred when fire was followed by normal precipitation [215]. Seedlings quickly develop a long taproot that enables them to reach moisture even on hot, burned-over soils [73,192]. Nevertheless, fall drought is a major factor in seedling mortality [171,197], and drought limits seedling establishment at interior ponderosa pine's lower elevational limit [34]. Since ponderosa pine in the Southwest has only a 2-month growing season before the onset of fall drought, winter dormancy, and spring drought, seedling mortality is usually highest in regions with bimodal precipitation [195]. Regeneration is further hindered in southwestern ponderosa pine on volcanic soils, which produce smaller seedlings (1-2 inches (3-5 cm)) that are more susceptible to frost-heaving compared to seedlings on other soils [155]. Rodent and ungulate browsing may result in considerable seedling loss. Artificial ponderosa pine regeneration on the Apache National Forest of east-central Arizona showed 95% mortality, and surviving seedlings showed little height gain, with mule deer and elk browsing. Seedlings in exclosures showed 2 to 3 times the height growth of unprotected seedlings [171]. However, some grazing regimes favor seedling establishment. Pearson [232] found that in the Southwest, heavy cattle grazing that reduced grass interference with ponderosa pine growth, followed by light cattle grazing that allowed tree seedlings to survive, favored ponderosa pine over herbaceous and shrub species.
On open sites with favorable moisture conditions, ponderosa pine seedlings often establish in large numbers. Dense seedlings often develop into "dog-hair" sapling thickets if stands are not thinned by fire or other means [77]. Boldt and Van Deusen [52] report that high tree densities (>1 tree/ft2) are common in naturally regenerated Rocky Mountain ponderosa pine stands in the Black Hills. An even-aged, 63-year-old stand that had never experienced fire contained 6,600 trees per acre (16 500/ha), with average DBH of 2.4 inches (6.1 cm). A number of researchers [61,257,284,285] have noted that ponderosa pine establishment in the Southwest tends to occur in pulses, during periods of relatively cooler, wetter climate conditions. Mast and others [213] found that rate of ponderosa pine seedling establishment varied 8-fold over the 3 centuries prior to fire exclusion; however, presettlement variability in seedling establishment was dwarfed by a 2-orders-of-magnitude increase in seedling establishment in the early 20th century compared to presettlement rates of ponderosa pine establishment. Brown and others [61] suggest that these pulses of dense tree establishment and rapid growth result in ladder fuels that increase the severity of the next fire.
Growth and mortality: Biomass allocation shifts as trees mature. As seedlings, ponderosa pine allocates relatively more biomass to roots compared to stems and leaves. Saplings tend to allocate relatively more of their biomass to foliage, and pole-sized trees allocate more biomass to woody tissue [132]. Open-grown ponderosa pines generally gain height rapidly in the sapling and pole stages [172], and mortality rates tend to level off at maturity. Knowles and Grant [184] found that ponderosa pines on the Colorado Front Range showed a sharp drop in mortality at age 115. The oldest trees sampled were 300+ years old. Minor [220] developed site index curves for ponderosa pine in the Southwest.
Dendrochronological and basal area analyses of trees in central Colorado showed that Rocky Mountain ponderosa pine growth was positively correlated with presence of Gambel oak and negatively correlated with presence of other Rocky Mountain ponderosa pines (r > 0.93). The authors attributed increased growth of ponderosa pine associated with Gambel oak to better soil nutrient status conferred by oak litter and/or increased density of ponderosa pine facilitated by Gambel oak nurse trees [42]. Klemmedson [183] also noted the benefit of Gambel oak litter to soil nutrient status and ponderosa pine growth. Gambel oak also grows in shrub form. Data on relative growth rate of young ponderosa pine in Gambel oak shrub stands are not available; however, Gambel oak shrubs may hinder growth of ponderosa pine rather than increase it (M. G. Harrington 2003, pers. comm).
SITE CHARACTERISTICS:Climate: Temperature regimens do not vary greatly across ponderosa pine's range: Pearson [233] found only a 2 oFahrenheit difference in mean annual temperature among ponderosa pine sites in Arizona, the Black Hills, Washington, and California. Length of frost-free period in ponderosa pine sites depends upon latitude and elevation. In much of its northern range, the frost-free period averages about 120 days. In the extreme Southwest, the frost-free period may lengthen to 240 days [69].
A common climatic factor occurs across Rocky Mountain and southwestern ponderosa pines' geographical range: Ponderosa pine grows on sites too dry to support other timber species. Although the drought season occurs at different times in different regions, periodic prolonged drought is characteristic across the pines' range and limits its expansion into drier types [33,69]. Overall, annual precipitation ranges from 15 to 20 inches (380-510 mm) per year. In low-elevation ponderosa pine sites at desert edges, mean precipitation is as low as 9 inches (230 mm) per year. In mixed-conifer forests at moderate elevations, mean annual precipitation may exceed 25 inches (640 mm) [33]. Climate grades from dry continental interior in the Northern Rocky Mountains to subhumid in south-central Wyoming, to semi-arid in the Southwest and the plains grassland-ponderosa pine woodland interface [33,84]. Climate is most favorable for regeneration in the Black Hills, where the wettest months are May and June and summer rains are frequent [33,68]. The Medicine Bow Range of Wyoming, which averages 25 inches (640 mm) annual precipitation, is the wettest portion of Rocky Mountain ponderosa pine's range. Precipitation on the east slope of the Medicine Bow ranges between 12 and 20 inches (300-510 mm), with 2/3rds received from April through September [84]. Transplant studies and abundant natural regeneration under fire exclusion suggest that climate is not limiting for Rocky Mountain ponderosa pine establishment on many plains grassland sites [134]. Climate in the Southwest has a distinct bimodal pattern of winter and summer precipitation. May and June are the driest months in Arizona and New Mexico [68,84]. Mean annual precipitation in ponderosa pine communities in western Texas is 16 to 20 inches (400-510 mm) [267].
Soils: Ponderosa pine occurs widely on igneous and sedimentary parent materials including basalt, volcanic cinder, limestone, and sandstone soils. Textures include clayey and silty loams, sandy loams, gravels, and cobbles [28,68,138,267]. Steep slopes with restricted soil profile development may support at least some ponderosa pine [68]. Degree of slope varies and can be steep: sites in the Santa Catalina Mountains of southeastern Arizona ranged from 10o to 36o [97].
Elevation: Ponderosa pine is most common from 6,000 to 8,500 feet (1,800-2,600 m) [33,84]. Elevation by state is:
| Arizona | 7,000-10, 000 feet | 2,300-3,050 m | [29,35,77,84,97] |
| Colorado | 6,360-9,500 feet | 2,120-2,900 m | [11,245] |
| Nebraska | around 4,000 feet | 1,200 m | [307] |
| New Mexico | 7,600-8,900 feet | 2,300-2,700 m | [135] |
| Nevada | 5,600-8,500 feet | 1,700-2,600 m | [45,192] |
| Texas | 6,000-7,000 feet | 1,800-2,000 m | [90,124] |
| Utah | 700-8,900 feet | 200-2,700 m | [216] |
Ponderosa pine is moderately shade tolerant. It tolerates shade better than quaking aspen, whitebark pine, and bur oak but less than western white pine, white spruce, and Rocky Mountain juniper. Unlike the first 4 species, ponderosa pine can establish beneath its own canopy [7,31].
On some sites, ponderosa pine can be a secondary colonizer in the absence of disturbance. GIS modelling of historic photographs of ponderosa pine-plains grassland ecotones on the Colorado Front Range shows a pattern of tree invasion into historic grassland, particularly on north slopes. Interior ponderosa pine invasion began around 1930, when effective fire exclusion began. Two strong pulses of ponderosa pine establishment onto grassland occurred in the 1970s and 1980s during years of above-average precipitation. Decreased livestock grazing may have also aided tree invasion into plains grassland in the 1970s and 1980s [214].
Rocky Mountain ponderosa pine is typically seral to white spruce in the Black Hills of Wyoming and South Dakota. It is climax on sites that are too dry to support white spruce. Quaking aspen may dominate a site following stand-replacing fire, but Rocky Mountain ponderosa pine typically establishes dominance after a few postfire decades. Bur oak may dominate some low-elevation stands in the northern Black Hills and Bearlodge Mountains of eastern Wyoming. Succession to Rocky Mountain ponderosa pine is "very slow" on sites where oak scrub stands develop after fire or other stand-replacing events [7].
East of the Continental Divide in Montana, Rocky Mountain ponderosa pine is a climax species. It forms heavily stocked, uneven-aged stands in the absence of fire [19,237]. In the Missouri Breaks region of Montana, bluebunch wheatgrass-western wheatgrass grasslands succeed to Rocky Mountain ponderosa pine/bluebunch wheatgrass habitats in the absence of fire. Shrubland areas of the region may succeed to Rocky Mountain ponderosa pine/Rocky Mountain juniper without fire [104].
Succession to conifers may be slow in southern New Mexico and Arizona. Aspect strongly influences rate of succession. In the White Mountains of south-central New Mexico, ponderosa pine establishment began 15 to 20 years after fire on northern and eastern exposures, while 75 or more years were sometimes required for forest development after fire on southern and western slopes. Once established, however, ponderosa pine cover was generally higher on drier slopes than on north-facing slopes [135]. Moir and Dietrich [221] present successional models for ponderosa pine in Arizona and New Mexico, both with fire and with fire exclusion.
SEASONAL DEVELOPMENT:Phenological development of ponderosa pine on the Fort Valley Experimental Forest of northern Arizona was as follows [261]:
| Event | Date |
| diameter growth begins | 15-30 May |
| vegetative buds swell | 1-15 May |
| vegetative bugs elongate | 15-25 May |
| root growth starts | 15-30 May |
| shoot elongation | 10-30 June |
| needle elongation | 15 June-30 July |
| shoot growth stops | 1 July-30 Aug. |
| seed germinates | 1 July-Aug. 30 |
| staminate buds appear | 20-31 May |
| pollination | 10-20 June |
| diameter growth stops | 1-20 Sept. |
| female cones developed | 15-20 Sept. |
| seeds ripe | 1-20 Oct. |
| needle shed | 1-30 Oct. |
| root growth stops | 15-30 Nov. |
Surface fire often kills ponderosa pine seedlings and saplings [50,57,87]; however, the effect is dependent upon fire severity and stand structure. Young trees in open canopies acquire fire-resistant traits rapidly, and 6-year-old saplings often survive low-severity surface fire. Fire is especially damaging in overcrowded young stands: the relatively denser foliage and thinner bark of trees in thick stands reduce resistance to surface fire. Such stands are also prone to crown fire [55].
Fire prepares a favorable seedbed for ponderosa pine regeneration. Periodic surface fire removes the heavy litter and duff that accumulate in ponderosa pine forests. Wind-borne seeds falling from the crowns of surviving or fire-killed trees land on a nutrient-enriched mineral seedbed under an open canopy that favors germination and seedling establishment [251,275,323]. Available soil moisture may increase when fire removes competing vegetation [133].
Fire regimes: Ponderosa pine evolved under a regime of frequent surface fires and infrequent mixed-severity and stand-replacement fires [52,281,307]. Presettlement fires in lower-elevation (<7,920 feet (2,400 m)) ponderosa pine communities were mostly low- to moderate-severity surface fires that maintained mostly open-grown, parklike stands, often with clusters of seedlings and saplings [33,61,63,74,77,117,149,278,280,298,305,311]. Prior to the 1900s, ponderosa pine was perpetuated by surface fires that recurred every 5 to 30 years. Fire-return intervals tended to be shorter in the warm, dry forests of the Southwest than in the cool, dry forests of the central Rocky Mountains or the cool, relatively moist forests east of the northern Rocky Mountains [61,77,95,141,198]. For example, Dieterich and Swetnam [96] report a 2-year mean fire-return interval for presettlement ponderosa pine on the Fort Valley Experimental Forest near Flagstaff; Laven and others [198] report a 45.8-year mean fire-return interval (range=20.9-66.0 years) for the Front Range of Colorado; and Brown and Sieg [63] report an average fire-return interval of 22 years for presettlement Rocky Mountain ponderosa pine forests of South Dakota. Gruell [131] provides an annotated record of wildfires that occurred throughout ponderosa pine's range during the settlement period (1776-1900). Historic fire regimes are summarized by state and region at the end of this section.
Fire history studies show mixed-severity fire regimes for ponderosa pine forests of the interior West and Southwest. Many forests experienced infrequent, large stand-replacement fires prior to the European-American settlement period [61]. For example, Laven and others [198] reported a range of 3 to 161 years (x̄ = 45.8) for the central Colorado Front Range. Small fires occurred on average every 20.9 years; large fire occurrence averaged 41.7 years. Higher-elevation (>7,920 feet (2,400 m)), relatively mesic mixed-conifer forests with Rocky Mountain ponderosa pine, Rocky Mountain Douglas-fir, and Rocky Mountain lodgepole pine tend to have more mixed-severity fires than lower-elevation Rocky Mountain ponderosa pine forests [280,315]. This is probably because herbaceous species recover from fire more quickly, and dry out earlier in the season, at low elevations. In the northern Colorado Front Range, mean fire-return interval for widespread (> 10 trees scarred), mixed-severity fire in higher-elevation forests during 1650 to 1920 ranged from 34 to 43 years; mean fire-return interval for widespread, mixed-severity fire in lower-elevation Rocky Mountain ponderosa pine forest was 14 to 24 years. During the same period, mean fire-return interval for localized (2-9 trees scarred) fires in higher- and lower-elevation ponderosa pine forest was 17 to 22 years and 8 to 18 years, respectively [298]. Ponderosa pine at 5,633 to 5,919 feet (1,707-1,804 m) in the Chiricahua Mountains experienced an historical fire-return interval ranging from 1 to 15 years (x̄ = 6.17 years) compared to a range of 1 to 31 years (x̄ = 7.96 years) in higher-elevation (6,801-7,002 feet (2,073-2,134 m)) mixed-conifer forest in the Chiricahua Mountains. Some higher-elevation mixed-conifer forests show an historical fire regime similar to lower-elevation ponderosa pine, however. Swetnam and Basian [280] suggest that mixed-conifer forests on dry, steep slopes—where fire can easily ignite and spread upslope from many directions—are most likely to experience frequent surface fire.
Native American burning influenced fire regimes in ponderosa pine ecosystems prior to and during European-American settlement, mostly by increasing fire frequency. For example, in the Chiricahua Mountains of southeastern Arizona, Seklecki and others [264] found that southwestern ponderosa pine showed a shorter fire-return interval (x̄ = 3.0 years, range=1.0-16.0 years) between 1700 and 1900, when Chiricahua Apaches inhabited the area, than in earlier periods when Apaches did not reside there. Dormant-season (spring) fires were also more frequent during that time period in southwestern ponderosa pine sites of the Chiricahua Mountains compared to southwestern ponderosa pine sites near the Mexican border, which were used as travel corridors but mostly unoccupied at that time [264]. Near the border, most fires occurred in the late May to early June growing season [280]. Fire frequency increased to 1 per year (an uncommonly low number for southwestern ponderosa pine) during the Spanish-Apache wars of 1760 to 1780, when Apaches probably used fire as a method of warfare. Fire occurrence terminated abruptly in the late 1880s, coincident with Apache resettlement to reservations (Geronimo surrendered in 1886) and increased livestock grazing by European-Americans [264,280].
Fire regimes in ponderosa pine also affect regimes of adjacent communities. In many cases, fire frequency has been reduced in adjacent communities because ignitions in ponderosa pine are suppressed and fire does not spread into adjacent communities. For example, stand structure of a shrub live oak-hairy mountain-mahogany (Quercus turbinella-Cercocarpus montanus var. paucidentatus) community on the Prescott National Forest of Arizona was historically a fire-maintained mosaic of different-aged chaparral. Mean fire frequency in the adjacent interior ponderosa pine/Arizona white oak (Q. arizonica) stand was 2 years. After over 100 years of fire exclusion, the chaparral stand was even-aged and senescent, with heavy accumulations of dead material. Ponderosa pine was encroaching into the chaparral [93].
Climate and fire frequency: Long-term fire history studies on the northern Colorado Front Range show that interannual variability in soil moisture is more conducive to widespread fire than drought alone. Fire occurrence, especially widespread fire, tends to increase 1 to 4 years after above-average moisture availability in spring-summer [298]. Similarly, fire occurrence tends to increase 2 to 3 years after above-average precipitation in winter-spring [30,281]. Climatic variation that produces widespread, stand-replacing fire has been associated with southern oscillation events. El Niño is associated with greater soil moisture and herbaceous fuel production in spring, with fire occurrence peaking several years after El Niño events. La Niña events are associated with dry springs, with fire occurrence peaking in the same year [298]. A decline in fire frequency in ponderosa pine forests of the Southwest coincided with reduced El Niño-La Niña events between 1780 and 1830 [284,298]. Alternating wet and dry years resulting from El Niño-La Niña events in the mid- to late 1800s increased fire frequency [298].
Fire exclusion: The ecological changes that have occurred in ponderosa pine forests over the last century have been well documented by a number of researchers [23,74,95,117,224,306,311,312]. Frequent, mostly light-severity surface fires thinned small trees, especially the less fire-resistant Rocky Mountain Douglas-fir and firs. The combined effects of 60 to 80 years of fire exclusion, logging that removed many overstory pines, heavy livestock grazing, and climate change have resulted in closed-canopy stands with dense understories and ladder fuels [57,62,63,76,77,117,256,281,292]. These changes have been documented throughout ponderosa pine's range in the Rocky Mountains and Southwest [76,209,271], and have also occurred in ponderosa pine/Douglas-fir and mixed-conifer types in those regions [178,298]. A fire history study of ponderosa pine stands near Flagstaff, Arizona, documented changes in stand structure over a 116-year period [77].
Stand structure in 1876 (reconstructed) and in 1992:| DBH class (inches) | 1876 | 1992 |
| 0-3.9 | 0.3 | 945 |
| 4-7.9 | 0.7 | 243 |
| 8-11.9 | 1.0 | 46 |
| 12-15.9 | 1.4 | 6.7 |
| 16-19.9 | 1.7 | 1.6 |
| 20-23.9 | 2.1 | 2.5 |
| 24-27.9 | 2.4 | 2.4 |
| 28-31.9 | 2.8 | 4.1 |
| 32-35.9 | 3.1 | 1.7 |
| 36-39.9 | 3.5 | 0.3 |
| 40-43.9 | 3.8 | 0.2 |
| Total | 22.8 | 1,253.5 |
When wildfire burns dense ponderosa pine stands under dry conditions, the abundant fuel quickly allows them to develop a high intensity and to spread into tree crowns. Severe, stand-replacing fires were less frequent in ponderosa pine forests in the past; now they are common [22,55,62]. Abundant litter and living and dead woody fuels feed wildfires of intensities and sizes that have not occurred for many centuries [280]. The increasingly frequent occurrence of large, crowning wildfires in ponderosa pine may indicate a shift to a fire regime characterized by very large (>100,000-acre (4,000 ha)) crown fires [76,255]. Data in Sackett and others [255] show a great increase in the number of acres burned by wildfire in Arizona and New Mexico since 1970. Over 100,000 acres (40,000 ha) burned from 1915 to 1990, with 70% of the fires occurring after 1970. Before 1970, total acreage burned per year never exceeded 130,000 acres (52 000 ha). After 1970 there were 8 years in which total acreage burned exceeded 119,000 acres (47 600 ha), with nearly 500,000 acres (200 000 ha) burning in 1989. On the Mexican side of the international border, where there is a lack of effective fire suppression, frequent, widespread surface fires have persisted in southwestern ponderosa pine and mixed-conifer forests [280].
Besides unprecedented, large-acreage severe fires, other ecological consequences of fire suppression in ponderosa pine ecosystems include [77,79,312]:
Organisms within ponderosa pine ecosystems have evolved with fire, and frequent fires are probably required to maintain ecosystem health [54,76,121,221,222]. Some researchers have questioned whether ponderosa pine ecosystems are sustainable under current conditions [23,77,265,280].
Fuels: Even within the same provenance, fuel loadings in ponderosa pine stands may vary greatly depending upon age class, stand structure, and understory composition [252]. Equations for predicting fuel loads in ponderosa pine are available [3,110,111,112,143,248]. Mean fuel loadings (tons/acre, 0-1 inch and > 1-inch fuels) have been calculated for ponderosa pine stands on 3 Reservations, 2 National Parks, and 8 National Forests of Arizona and New Mexico. The study involved 62 sites: mean forest floor loading for the entire 62 stands was 12.5 tons/acre (4.1 t/ha) [249].
Absence of fire in ponderosa pine and mixed-conifer forests has led to large accumulations of surface and ground fuels [29,175,252]. Structurally, fire exclusion has led to vertical continuity, with Douglas-fir, firs, and other shade-tolerant, less fire-resistant species in the understory. These late-successional species become ladder fuels that encourage crown fires in ponderosa pine and mixed-conifer forests [29,55]. In the dry southwestern climate, the natural accumulation of pine needles and woody fuels is exacerbated by slow decomposition [149].
State and regional fire regime studies
Black Hills and interior Northwest:
Fire season on the Rocky Mountain ponderosa pine-grassland savannas of eastern
Montana and the Dakotas peaks in July and August, when the majority (73%) of
lightning-strike ignitions occur. Wildfire season generally extends from April
to September [161]. Frequent surface fires historically burned litter and killed
young Rocky Mountain ponderosa pines and other nonsprouting woody species encroaching
into grasslands [50,53]. Rocky Mountain ponderosa pine
in the Black Hills was historically characterized by
2 communities: interior forest and savanna. These treed landscapes were
described as "islands" surrounded by plains grasslands. Brown and Sieg
[63] found that in
interior forest sites at Jewel Cave National Monument, South Dakota, fire-return
intervals from the 1500s to the late 1800s averaged 20 to 24 years, with a range
of 1 to 93 years. Fire-return intervals at savanna sites in Wind
Cave National Park, South Dakota, averaged 10 to 12 years, with a range of 2 to
23 years. The fire-return interval of Rocky Mountain ponderosa pine savannas is the shortest documented
for northern ponderosa pine ecosystems, and is similar to the short-return
intervals of ponderosa pine forests in the Southwest[63].
Invasion of Rocky Mountain ponderosa pine onto grasslands, and increased tree density in formerly open savanna, is thought to be largely attributable to reduced fire frequency, although grazing has probably contributed to increased Rocky Mountain ponderosa pine density on forest and grassland margins [50,53]. A little less than one-half of Rocky Mountain ponderosa pine in the Black Hills are single-storied, even-aged stands that developed after crown fires or mountain pine beetle epidemics [7]. "Dog-hair" Rocky Mountain ponderosa pine thickets are common on many sites [50].
Brown and others [62] report the following measures of fire frequency on Rocky Mountain ponderosa pine sites of less than 50 acres (20 ha) in size:
| Site | Period of Analysis | No. of Intervals | Median Fire Interval (years) | Range of Intervals (years) | Years Since Last Fire |
| Black Hills, SD | 1580-1887 | 9 | 23 | 11-74 | 110 (1887-1997) |
| Black Hills, SD | 1668-1890 | 7 | 22 | 13-72 | 107 (1890-1997) |
| Medicine Bow NF, WY | 1436-1911 | 15 | 26 | 8-74 | 86 (1911-1997) |
| Medicine Bow NF, WY | 1460-1909 | 12 | 33.5 | 8-82 | 88 (1909-1997) |
| Arapahoe-Roosevelt NF, WY | 1568-1861 | 4 | 80.5 | 10-122 | 136 (1861-1997) |
| Arapahoe-Roosevelt NF, WY | 1568-1887 | 3 | 117 | 80-122 | 110 (1887-1997) |
| Rio Grande NF, CO | 1528-1896 | 26 | 9.5 | 2-41 | 101 (1896-1997) |
Colorado: A fire history study of a 10,000-acre (4000 ha) Rocky Mountain ponderosa pine-Rocky Mountain Douglas-fir site in central Colorado showed a pattern of frequent surface fires from 1197 to 1851. Large stand-replacement fires were rare, but several landscape-level fires are documented. Intervals between fire years ranged from 1 to 128 years at the landscape scale and from 1 to 58 years for individual stands. Fires occurred throughout the growing season. Fire size varied across time; for example, numerous small fires occurred in the 1500s, while landscape-level fires occurred in 1631, 1696, and 1723. After 1723, there were few fires until 1851; that fire was a stand-replacement, mixed crown and severe surface fire that covered most of the landscape. There have been no extensive fires in the study area since 1851, and most stands have not experienced fire for nearly 100 years [61].
Utah: Fire history studies show a range of 3 to 47 years for ponderosa pine stands in southern Utah [64,209,272]. Studies in Zion National Park show a presettlement fire frequency range of 3 to 12 years for ponderosa pine and mixed-conifer forests. All study sites had experienced fire at least once a decade [64,209]. Large fires that burned more than 1,000 acres (400 ha) occurred nearly every 3 years on the Horse Pasture Plateau. Fire-return intervals declined greatly in the Park beginning in the late 1800s, when the study sites were subject to intense livestock grazing and grassy fuels were sparse. Although grazing has not been practiced in the Park for over half a century, fire events remained infrequent under fire exclusion [209].
Nebraska: Bragg [56] evaluated fire frequency for upland Rocky Mountain ponderosa pine-bur oak forest adjacent to sandhills prairie. He found a mean fire-return interval of 3.5 years from 1851 to 1900, and a mean of 8.5 years for 1901 to 1950. Without the frequent fires that killed the seedlings, upland interior ponderosa pine has invaded sandhills prairie communities [176].
Texas: A fire history study in mixed ponderosa pine-Rocky Mountain Douglas-fir-southwestern white pine in Guadalupe Mountain National Park showed a mean return interval between major fires of 17 years prior to the 1920s; mean return interval for all fires detected between 1696 and 1922 was 4.7 years. Evidence of fire scarring was not found after 1922. Portions of the montane forest have a 2-tiered structure of overstory ponderosa pine and Rocky Mountain Douglas-fir, and dog-hair thickets of ponderosa pine and Douglas-fir saplings in the understory [1,2].
New Mexico: Swetnam and Dieterich [286] found that extensive surface fires were common in the Gila Wilderness of southwestern New Mexico before 1900. Mean fire return interval from 1633 to 1900 was 4 to 8 years, with a range of 1 to 26 years. Most fires burned throughout the 40.5-hectare study sites, although a few fires appeared to be smaller. Fire history was similar in northern New Mexico, with extensive high-frequency, low-severity surface fires in low-elevation (<5,545 feet (1,690 m)) ponderosa pine forests and less frequent surface fires, along with some patchy crown fires, in higher-elevation mixed-conifer Rocky Mountain Douglas-fir-ponderosa pine-white fir forest. Across sites, the minimum fire-return interval for major fires (>10% of trees scarred) in ponderosa pine before 1900 ranged from 1 to 9 years; maximum fire-return interval for major fires ranged from 16 to 51 years (x̄ = 9.4 years). In mixed-conifer forests where ponderosa pine was codominant, minimum and maximum fire-return interval ranges were 1 to 4 and 21 to 46 years, respectively (x̄ = 10.4 years). Fires were rare in ponderosa and mixed-conifer forest after the 1850s. The authors attribute the initial drop in fire frequency to domestic sheep grazing by the Navajos, and by the 1900s, to fire exclusion by European-Americans [292].
Arizona: The sky islands of southeastern Arizona have among the highest incidences of lightning-caused fires in the United States [185,260]. The lightning fire season begins in late April, peaks in June, and runs into October. Maximum lightning fire incidence is above 6,000 feet (1,800 m): directly within the ponderosa pine zone [29]. Prior to the 1880s, surface fires burned through ponderosa pine sky islands once or twice a decade, but occurred at highly variable intervals. Fires were somewhat less frequent in higher-elevation, mixed-conifer forests [281]. Swetnam and others [282] reported a 9- to 22-year range (x̄ = 14.6) for presettlement fire-return intervals in ponderosa pine-Douglas fir in Chiricahua National Monument. Fire frequency in ponderosa pine-Arizona pine forests in southeastern Arizona was greatly reduced after Euro-American settlement in the 1870s. Heavy livestock overgrazing coupled with fire exclusion caused both the number of fires and the area burned to decline dramatically [238].
Historical fire frequency in the ponderosa pine-encinal oak woodland interface has been documented at least 1 fire per decade [85,174]. In ponderosa pine-oak woodlands of the Rincon Mountain Wilderness, fire regime was dominated by large-scale (> 500 acre (200 ha)), early-season (May-July) surface fires. Mean fire-return interval from 1657 to 1893 was 6.1 years, with a range of 1 to 13 years. From 1748 to 1996, mean fire-return interval in the high-elevation mixed-conifer type was 9.9 years, with a range of 3 to 19 years [30].
There are many plant communities and ecosystems in which southwestern or Rocky Mountain ponderosa pine is either dominant, an important component of the vegetation, or an invader. Historic fire-return intervals for these communities and ecosystems are summarized below. Find further fire regime information for the plant communities in which these varieties may occur by entering "ponderosa pine" in the FEIS home page under "Find Fire Regimes".
| Community or Ecosystem | Dominant Species | Fire Return Interval Range (years) |
| Nebraska sandhills prairie | Andropogon gerardii var. paucipilus-Schizachyrium scoparium | <10 |
| sagebrush steppe | Artemisia tridentata/Pseudoroegneria spicata | 20-70 [231] |
| mountain big sagebrush | A. t. var. vaseyana | 20-60 [21,65] |
| Wyoming big sagebrush | A. t. var. wyomingensis | 10-70 (40**) [299,321] |
| plains grasslands | Bouteloua spp. | <35 |
| blue grama-needle-and-thread grass-western wheatgrass | B. gracilis-Hesperostipa comata-Pascopyrum smithii | <35 |
| blue grama-buffalo grass | B. g.-Buchloe dactyloides | <35 |
| grama-galleta steppe | B. g.-Pleuraphis jamesii | <35 to < 100 [231] |
| curlleaf mountain-mahogany* | Cercocarpus ledifolius | 13-1000 [24,263] |
| mountain-mahogany-Gambel oak scrub | C. l.-Quercus gambelii | <35 to < 100 |
| Rocky Mountain juniper | Juniperus scopulorum | <35 |
| wheatgrass plains grasslands | Pascopyrum smithii | <35 [231] |
| Engelmann spruce-subalpine fir | Picea engelmannii-Abies lasiocarpa | 35 to > 200 [20] |
| pinyon-juniper | Pinus-Juniperus spp. | <35 [231] |
| Rocky Mountain lodgepole pine* | Pinus contorta var. latifolia | 25-300+ [17,20,244] |
| Colorado pinyon | P. edulis | 10-49 [231] |
| southwestern ponderosa pine* | P. ponderosa var. brachyptera | 2-46 |
| Rocky Mountain ponderosa pine* | P. ponderosa var. scopulorum | 2-46 [20,30,198] |
| Arizona pine* | P. arizonica | 2-10 [20] |
| quaking aspen (west of the Great Plains)* | Populus tremuloides | 7-120 [20,130,218] |
| mountain grasslands | Pseudoroegneria spicata | 3-40 (10**) [17,20] |
| Rocky Mountain Douglas-fir* | Pseudotsuga menziesii var. glauca | 25-100 [20] |
| bur oak | Quercus macrocarpa | <10 [300] |
| oak savanna | Q. m./Andropogon gerardii-Schizachyrium scoparium | 2-14 [231,300] |
POSTFIRE REGENERATION STRATEGY [275]:
Tree without adventitious bud/root crown
Crown residual colonizer (on-site, initial community)
Initial off-site colonizer (off-site, initial community)
Secondary colonizer - off-site seed
On the Coconino National Forest, a mixed-severity wildfire on 7-9 May 1972 killed about 1/4th of the standing ponderosa pine where surface fire severity was moderate. Mortality was greatest in the smaller diameter classes: 90% of "pulpwood" and 7% of "sawtimber" were killed by moderate-severity fire. Approximately 2/3rds of the trees were killed where severe surface or crown fire occurred. Ninety percent of pulpwood and 50% of sawtimber trees were killed where fire was severe. The summer after the fire, basal area was 211 ft2/acre (47.5 m2/ha) on an adjacent unburned area; 103 ft2/acre (23 m2/ha) on the moderate burn; and 40 ft2/acre (9 m2/ha) on the severe burn [66].
DISCUSSION AND QUALIFICATION OF FIRE EFFECT:Dormant-season studies indicate that bud kill, which is related to fire season, is more important than foliage kill in determining chances of ponderosa pine survival after burning [142,301,302]. Wagener [302] and Harrington [144] found the minimum requirement for ponderosa pine survival was 90% or less scorch with 50% or more of buds and twigs remaining. Five years after prescribed burning on the San Juan National Forest of Colorado, Harrington [144] found significant (P=0.05) differences in mortality of scorched ponderosa pine, depending upon season of burning. Mortality was lowest for fall-scorched trees (5%), and spring-scorched trees showed less mortality than summer-scorched trees (17% vs. 21%, respectively). Ninety percent of fire-damaged ponderosa pine that died had done so by postfire year 4. Most trees greater than 7.2 inches (18 cm) diameter survived fall burning even with 90% scorching. With spring and summer burning, trees less than 4 inches (10 cm) diameter died with greater than 50% scorching, while at least 90% scorching was required before trees larger than 4 inches (10 cm) in diameter were killed by spring or summer fire [142].
Mortality models: McHugh and Kolb [217] reported that model using total crown damage by fire (scorch + consumption) and bole char severity as independent variables best predicted mortality of individual ponderosa pine trees after prescribed or wildfires in northern Arizona. A study on the Colorado Front Range found that crown scorch, expressed as a percentage of the prefire live crown length, was the best determinant of postfire mortality of ponderosa pine [318]. Wyant and Zimmerman [319] found that degree of crown scorch and tree size were the most effective predictors of postfire survival potential after September prescribed burning in Colorado. They provide a model for estimating mortality based upon percent crown scorch and DBH [319].
Mortality studies: Harrington and Hawksworth [148] conducted prescribed burning treatments to ponderosa pine on the South Rim of Grand Canyon National Park, Arizona. They found that mortality generally increased with decreasing tree size and increasing crown scorch, bole char, and southwestern dwarf-mistletoe infection. Trees with greater than 87% crown scorch experienced 100% mortality, even in the 30- to 36-inch (76-92 cm) size class. Severe bole char resulted in 67% mortality. Mortality by size class is indicated in the the graph below [148].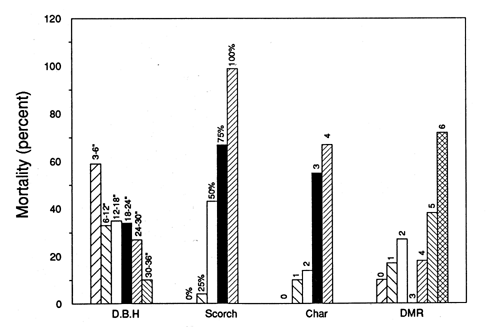 |
| Char rating: 0=unburned around the tree, 1=duff burned but no char, 2=light bole char,
3=moderate bold char, 4=severe bole char with much bark consumed DMR (dwarf-mistletoe rating): 0,=no mistletoe, 1=trace, 2=light, 3=moderate but not throughout tree, 4=moderate throughout tree, 5=most of tree heavily infected, 6=entire tree heavily infected [148]. |
On the Coconino National Forest, the general effect of a 3 October prescribed fire was a thinning from below and a 70% reduction in duff. No ponderosa pines less than 4.5 inches (10 cm) tall survived the fall fire. Mortality was less in pole-sized and larger trees. Severely damaged trees died within 2 years. Percent damage and postfire mortality of ponderosa pine by size class* and fire severity are given below [89].
| Saplings | Poles | Sawtimber | All trees | |||||||||
|
|
|
|
|||||||||
| Crown damage: | ||||||||||||
| none |
|
|
|
|
||||||||
| light |
|
|
|
|
||||||||
| moderate |
|
|
|
|
||||||||
| severe |
|
|
|
|
||||||||
| Dead after 2 years: | ||||||||||||
| none |
|
|
|
|
||||||||
| light |
|
|
|
|
||||||||
| moderate |
|
|
|
|
||||||||
| severe |
|
|
|
|
||||||||
Mature ponderosa pine may show increased growth following low-severity surface fires. Following prescribed fall burning on the Front Range of Colorado, mean fascicle lengths and bud sizes (length and diameter) were significantly greater (P<0.01) on Rocky Mountain ponderosa pines that received burning treatment than those on unburned control sites. Sutherland and others [279] present a linear regression model to predict postfire radial growth of southwestern ponderosa pine after prescribed fire.
DISCUSSION AND QUALIFICATION OF PLANT RESPONSE:Thinning to remove small-diameter trees, accompanied by prescribed fire, has been suggested as a means of restoring structure and function to degraded ponderosa pine ecosystems [18,43,76,103,103,140,147,255,306]. Frequent low-severity surface fires restore ecosystem function by thinning dense stands and reducing woody debris and other organic matter on the forest floor. This can result in increased soil moisture, increased soil temperature (with accompanying rates of increased litter decomposition, soil nutrient cycling, and fine root growth), increased productivity of understory herbs and shrubs, increased basal diameter growth of overstory ponderosa pine, and favorable seedbeds [6,7,77,78]. Fire-pruning of lower pine branches opens the canopy, aiding production of wildlife shrubs such as snowberry and chokecherry [128]. Frequent prescribed fires reduce fire hazard without damaging overstory ponderosa pine [6,7]. Biswell [43] listed several ways in which prescribed burning reduces wildfire hazard in ponderosa pine:
 |
| Grand Canyon National Park prescribed fire hand crews "stringing fire" in a North Rim burn unit, in cautious incremental strips, to keep burn severity to prescribed levels. |
If several fire cycles have been missed, thinning presettlement trees and manually removing heavy fuels from the base of large trees may be necessary to in order to protect old growth from severe scorching or death [76]. Harrington [144] recommends growing-season (spring or summer) burning in ponderosa pine forests if the management objective is thinning from below, and fall prescribed burning if stand losses must be minimized. Weather parameters for prescribed burning in southwestern ponderosa pine [149] and a logistic regression model predicting probability of ponderosa pine mortality by tree size, scorch class, and season of injury are available [142].
Allen and others [9] provide ecologically based recommendations for restoring southwestern ponderosa pine. They stress that restoration programs should include natural variability in southwestern ponderosa pine stands and the reestablishment of natural processes. Managers are encouraged to fully review their recommendations. A synopsis of their principles for restoration follows [9]:
Swetnam and Dieterich [286] recommend allowing large (>3,000 acres (1,200 ha)) prescribed natural surface fires in southwestern ponderosa pine in wilderness areas such as the Gila Wilderness. Based upon their fire history research, which showed evidence of mostly extensive but also small fires, they also recommend allowing small and patchy mixed-severity fires in approved areas, subject to the limitations of wilderness boundaries, visitor safety, and management and suppression capabilities [286].
Mixed and stand-replacing fire: High-elevation ponderosa pine forests of the Rocky Mounttains and Southwest were historically denser, with fewer fires of higher severities, than lower-elevation forests. Even at low elevations, north-facing slopes and mesic ravines probably supported dense forests that experienced some moderate-severity and stand-replacing fires. Evidence of a regime of frequent surface fires and occasional moderate-severity and stand-replacement fires suggests that a single prescription cannot capture historical variability of fire regimes in ponderosa pine types [298].
Historical occurrence of large, widespread prehistoric fires in ponderosa pine demonstrates the potential for large portions of montane zones to burn during a single year. Interannual climatic variability, particularly El Niño-La Niña events, can greatly increase fine fuels. Fuels management through thinning and prescribed burning reduces the probability of widespread wildfire in most years, but fuel management may not reduce fire hazard enough to prevent stand-replacing fire during years when dry weather is exceptionally conducive to rapid fire spread. It may be helpful for public education programs on fire hazard reduction to emphasize that although frequent surface fires were most common in ponderosa pine stands, mixed and stand-replacement fires did occur in prehistoric times, and that during years of high fire hazard, stand-replacement fires may occur again despite fuels reduction programs [298].
Fuels: Fuel loads in pole-sized thickets of ponderosa pine can be high. Ponderosa pine's long needles form a loose litter layer that burns readily [246]. Near Flagstaff, fuels in ponderosa pine stands averaged 28.3 tons per acre (63.4 t/ha), 21.3 tons (19.2 t) of which was duff [77]. Eakle and Wagle [100] provide a model, developed on the Fort Apache Reservation, for estimating fine fuels in southwestern ponderosa pine stands. Harrington [145] presents a model for estimating forest floor consumption in southwestern ponderosa pine forest based upon moisture content of the H surface soil layer.
Beaufait [37] found that backfires in ponderosa pine needles spread more slowly and had less flame depth, longer residence time, and a higher rate of energy release than headfires.
Boldt [51] provides prescriptions for thinning pole-sized "dog-hair" thickets of ponderosa pine using sequential treatments. Additional information on thinning "dog-hair" stands is found in Sackett and others [254].
Range: Prescribed fire can enhance understory forage production by reducing forest floor depth, tree density, and allelopathic toxins, and by increasing nutrient availability [13,150]. On the Fort Apache Reservation in Arizona, Gambel oak had reached mid-story in a ponderosa pine forest. Prescribed surface fires at 5- to 7-year intervals reduced Gambel oak to an understory shrub that was readily available to browsing animals. Coverage of the dominant grass, mountain muhly, increased with prescribed fire [43].
Pearson and others [234] reported that on the Wild Bill Range of Arizona, an area of ponderosa pine that had been thinned to 20 square feet basal area per acre (4.5 m2/ha) did not show a significant reduction in density following a wildfire. The fire crowned and killed the trees in an adjacent unthinned stand, however. Forage production increased as much as 578 pounds per acre (650 kg/ha) from prefire levels on sites were stand-replacement fire occurred, while pre- and postfire forage production were similar on thinned and burned sites [234].
Wild ungulate foraging, especially that of bighorn sheep, increased after late April prescribed fire in a ponderosa pine/sedge-bluegrass (Carex-Poa spp.) community in Custer State Park, South Dakota. Grass production did not increase after the fire, but forb production was significantly increased on burned compared to unburned sites (P<0.01) [102].
October prescribed burning brought a 2-fold increase in nitrogen content of Arizona fescue and associated grasses on an ponderosa pine/Arizona fescue community on the Fort Valley Experimental Forest near Flagstaff. Concentrations of potassium, phosphorus, calcium, and magnesium were also generally greater in forb and grass species on burned plots than in herbs on unburned control plots. Understory biomass was significantly greater on burned plots (P<0.05). The 1st fall after burning, understory yield was twice as great on burned plots than on unburned plots [150].
Overgrazing can greatly reduce fire frequency by removing understory fuels. Fire history studies of southwestern ponderosa pine document the near-cessation of fire in the mid-1800s due to livestock grazing in Arizona and New Mexico [292,293].
Exclusion of the recurrent fires that once swept the ponderosa pine-plains grasslands interface has resulted in interior ponderosa pine invasion into the grassland. Expansion is likely to continue without application of prescribed fire [50,273].
Since fire kills ponderosa pine seedlings and some saplings, prescribed fire can be used to reduce the density of encroaching pines. Late-April fire in a little bluestem-hairy grama-sideoats grama grassland in the Black Hills of South Dakota caused 79% mortality of Rocky Mountain ponderosa pine seedlings. Mean density of ponderosa pine seedlings was 8,132 per acre (3,251/ha) before the fire and 1,669 per acre (4,173/ha) in late summer, after burning. The fire reduced surface fuels by 32% [123].
Wildlife: Fire benefits most wildlife species inhabiting ponderosa pine ecosystems. Among bird species, fires in ponderosa pine tend to increase guilds that use open stands and snags. On the Prescott National Forest in Arizona, a stand-replacement burn in ponderosa pine attracted relatively more granivores, aerial insectivores such as flycatchers, and bark-feeding insectivores such as woodpeckers, while unburned areas attracted more ground and foliage insectivores [47]. A 20-year wildlife study on 4 stand-replacement burns near Flagstaff had similar findings for birds. Most rodent species, except chipmunks, also increased on burned ponderosa pine sites relative to adjacent unburned control sites. Wild ungulate use increased greatly on burned sites compared to unburned sites. Mule deer use of burned sites declined at postfire year 1, but increased to 2.5 times that of unburned sites for the next 19 years. Elk use also dropped at postfire year 1 relative to unburned sites, but increased to 3 times that of controls until postfire year 20, when use of burned and unburned sites was nearly equal. Elk use of the burns peaked at postfire year 7 [203].
Availability of ponderosa pines as nesting sites for cavity-nesters, particularly secondary cavity-nesters, is limited due to the scarcity of large, old-growth trees. Brawn and Balda [59] reported that violet-green swallow, pygmy nuthatch, and western bluebird populations increased after artificial nest boxes were placed in open and thinned plots, but not on untreated (dense) plots. Prescribed underburning that leaves large trees may encourage nesting. Prescribed fire in Rocky Mountain ponderosa pine stands in Wind Cave National Park, South Dakota, significantly increased the number of breeding bird pairs and deer mice on burned sites compared to unburned control sites (P<0.05) [49]. Creating small openings with prescribed fire may also promote nesting. Aulenbach and O'Shea-Stone [27] noted differential songbird use of burned sites created by a small, crowning wildfire compared to adjacent unburned sites in Rocky Mountain ponderosa pine on the Front Range of Colorado. Several species including red-breasted nuthatch, chipping sparrow, yellow-rumped warbler, and northern flicker occurred only on the burn. American robin, Steller's jay, and dark-eyed junco preferred the burn but used the control, while pygmy nuthatch, downy woodpecker, white-breasted nuthatch, and mountain chickadee occurred only on the unburned site [27].
Standing time of ponderosa pine snags is somewhat predictable. Harrington [146] found that 75% of fire-killed ponderosa pine snags on the San Juan National Forest of southwestern Colorado fell within 10 postfire years. Fall rates were not significantly different (P=0.1) among trees 2 to 16 inches (5-41 cm) DBH, but large trees that died quickly after 80% or greater crown scorch were likely to fall more quickly, while large trees that survived 2 or 3 postfire years before succumbing were likely to remain standing for longer periods of time [146].Bock, Jane H.; Bock, Carl E. 1984. Effects of fires on woody vegetation in the pine-grassland ecotone of the southern Black Hills. The American Midland Naturalist. 112(1): 35-42. [50].
SEASON/SEVERITY CLASSIFICATION:Two cool-season fires were conducted. The fall fire was a 157-acre (63 ha) burn on 17 October 1979. The fire was set at mid-day and passed over the area in 4 hours. A few snags smoldered until the next morning, when it started raining. Fire weather conditions were [49]:
windspeed 10 miles/h (16 km/h)The spring fire was an 815-acre (326 ha) burn on 14 April 1980. The fire was set at 10:30 a.m. and burned most of area within control lines within 36 hours. It was declared out on 15 May 1980. Fire weather conditions were [49]:
windspeed: 5 miles/h (8 km/h)The effects of the 2 fires were similar, so pre- and postfire litter and vegetation data were pooled for the 2 sites. Litter depth averaged 1.61 inches (4.02 cm) before fire [49].
FIRE EFFECTS ON TARGET SPECIES:| Variable | Prefire | Postfire year 1 | Postfire year 2 | Chi-square | |||
| Burn | Control | Burn | Control | Burn | Control | ||
| Canopy | 56.2 | 57.4 | 48.3 | 57.2 | 47.3 | 56.5 | 10.44 |
| Trunks and immatures | 3.3 | 2.5 | 1.2 | 2.3 | 1.2 | 2.4 | 22.26 |
Litter depth was significantly reduced to a 0.76-inch (1.89 cm) depth by burning (P<0.01) [49].
FIRE MANAGEMENT IMPLICATIONS:Harrington, Michael G. 1987. Ponderosa pine mortality from spring, summer, and fall crown scorching. Western Journal of Applied Forestry. 2: 14-16. [144].
SEASON/SEVERITY CLASSIFICATION:| Size class | Cover (%) |
| 0.5 to 4 inches (1-10 cm) | 8 |
| 4 to 7 inches (10-18 cm) | 31 |
| 7 to 10 inches (18-28 cm) | 42 |
| > 11 inches (> 28 cm) | 19 |
Precipitation was above the 16.8-inch (427 mm) annual mean 3 months preceding each burn, ranging from 2.8 inches (710 m) above normal before the spring burn to 0.4 inch (102 mm) above normal for the summer burn (fall ppt. not stated). The study site received about 2.2 inches (560 mm) of rain the July prior to summer burning, which probably encouraged tree growth. Precipitation following the fire treatments was 0.5 inch (130 mm), 3.8 inches (965 mm), and 3.5 inches (890 mm) above average for fall, spring, and summer treatments, respectively [144].
FIRE DESCRIPTION:| Fall | Spring | Summer | |
| Temperature (oF) | 56-63 | 65-79 | 72-77 |
| Humidity (%) | 32-41 | 14-41 | 21-37 |
| Wind speed (mph) | 2-5 | 1-6 | 1-5 |
| L-layer (% moisture) | 7.2 | 5.4 | 6.0 |
| F-layer (% moisture) | 11.8 | 6.9 | 7.5 |
| H-layer (% moisture) | 18.9 | 11.5 | 9.2 |
| Spread rate (ft/min) | 6 | 8 | 5 |
| Flame length (ft) | 3.0 | 3.5 | 3.5 |
At the end of 5 years, overall Rocky Mountain ponderosa pine mortality from fall crown scorch was significantly less than that from spring or summer (P=0.05). Tree mortality equaled 12, 26, and 29% for fall, spring, and summer fires, respectively. Differences in mortality between seasons were most substantial during the 1st postfire season: only 5% of fall-scorched trees died, compared with 17% of spring-scorched trees and 21% of summer-scorched trees. Subsequent mortality for all seasons was relatively similar over the length of the study, suggesting good survival potential for those trees not killed outright by scorching. Generally mortality decreased as DBH increased regardless of season. Trees less than 7 inches (17.5 cm) DBH accounted for approximately 85% of the observed mortality. Mortality from fall burns was comparable to that recorded on the control plots. Mortality in trees in the < 4-inch and 7 to 10.9-inch DBH classes was significantly greater on spring and summer burn plots than on fall burn and unburned plots. Percent mortality of scorched trees by diameter class and season of fire is given below. Means in rows followed by different letters differ at the 5% significance level. Means in columns followed by different numbers differ at the 5% level [144].
| Treatment | Tree diameter class (inches) | |||
| < 4.0 | 4.0-6.9 | 7.0-10.9 | 10.9 | |
| fall | 28a,1 | 18ab,1 | 0c,1 | 7bc,1 |
| spring | 55a,2 | 25b,1 | 11b,2 | 7b,1 |
| summer | 60a,2 | 37a,1 | 15b,2 | 5b,1 |
| control | 17a,1 | 6b,2 | 5b,1 | 1b,1 |
Scorch damage of up to 90% produced minimal mortality regardless of season of burn. When more than 90% of the crown was scorched, fall burning caused only 1/3rd the mortality produced by spring or summer burning. Percent mortality of scorched trees by scorch class and season of burn is presented below. Means in rows followed by different letters differ at the 5% significance level. Means in columns followed by different numbers differ at the 5% level [144].
| Treatment | Scorch class | |||||
| 0 | 10-33 | 34-66 | 67-89 | 90-99 | 100 | |
| fall | 5a | 8a,1 | 5a,1 | 0a,1 | 13a,1 | 39b,1 |
| spring | 5a | 5a,1 | 13a,1 | 12a,2 | 54ab,2 | 100b,1 |
| summer | 5a | 11a,1 | 12a,1 | 11a,2 | 42a,2 | 89a,1 |
Trees subjected to dormant-season burning were resistant to severe crown scorch damage. Of those trees sustaining 100% damage, only 10 of 28 (39%) ultimately died. All 8 trees sampled in the 7 to 10.9-inch (17.5-27.3 cm) category survived. Harrington [141] cautioned that this resistance to fire mortality is demonstrated for trees where crown scorch was the only type of fire damage recorded; such extensive damage would probably be fatal when combined with bole damage.
FIRE MANAGEMENT IMPLICATIONS:Sackett, Stephen S. 1984. Observations on natural regeneration in ponderosa pine following a prescribed fire in Arizona. Res. Note RM-435. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 8 p. [251].
Sackett, Stephen; Haase, Sally; Harrington, M. G. 1993. Restoration of southwestern ponderosa pine ecosystems with fire. In: Covington, M. Wallace; Debano, Leonard F.; Covington, W. W., tech. coords. Sustainable ecological systems: implementing an ecological approach to land management: Proceedings; 1993 July 12-15; Flagstaff, AZ. Gen. Tech. Rep. RM-247. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 115-121. [255].
Sackett, Stephen S.; Haase, Sally M.; Harrington, Michael G. 1996. Lessons learned from fire use for restoring southwestern ponderosa pine ecosystems. In: Covington, Wallace; Wagner, Pamela K., technical coordinators. Conference on adaptive ecosystem restoration and management: restoration of Cordilleran conifer landscapes of North America: Proceedings; 1996 June 6-8; Flagstaff, AZ. Gen. Tech. Rep. RM-GTR-278. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 54-61. [254].
SEASON/SEVERITY CLASSIFICATION:Seedlings: < 4.5 feet tall, 395 stems/acre (< 1.4 m, 975 stems/ha)
Saplings: 0.6 to 3.9 inches DBH, 1,114 stems/acre (1.5-9.9 cm DBH, 2,750 stems/ha), age = approximately 57 years
Poles: 4.0 to 10.9 inches DBH, 195 stems/acre (10.0-27.5 cm DBH, 700 stems/ha), age = 57 to 150 years
Sawtimber: > 11 inches DBH, 54 stems/acre ( >27.5 cm DBH, 130 stems/ha), age = 200 to 500 years
Relative tree cover on the experimental plots consisted of the following: sapling 17%; pole 62%, sawtimber 17.3%; small openings comprised the remaining 3.7%. Irregular distribution of the pine understory resulted in numerous grass-dominated openings [251]. Fendler's ceanothus (Ceanothus fendleri), mountain muhly (Muhlenbergia montana), bottlebrush squirreltail (Elymus elymoides), pine dropseed (Blepharoneuron tricholepis), Cainville thistle (Cirsium calcareum), groundsel (Senecio spp.), and goldenrod (Solidago spp.) were understory associates [255].
TARGET SPECIES PHENOLOGICAL STATE:Results of fire scar analysis indicate that the study area had not burned since 1876; prior to that time, mean fire-return interval was 2 years [255].
FIRE DESCRIPTION:Ignition began at 5:30 p.m. and continued until 11:00 p.m. Backing fires were used initially, but later in the evening when these ceased to carry effectively, short strip headfires with 30 to 40 feet (9-12 m) between strips were utilized. Although rate of spread approximated 4 to 6 feet per minute (1.2-1.8 m/min), it sometimes increased to as much as 12 feet per minute (3.7 m/min). Flame lengths in surface needles rarely exceeded 16 inches (40 cm). Air temperature dropped from 59 degrees Fahrenheit (15 oC) at 5:30 p.m. to 37 degrees Fahrenheit (3 oC) at 12:00 a.m., when most of the flaming combustion was complete. Measured surface needle moisture content increased from 8% to 12% during the night as temperatures fell; similarly, ground fuel (F & H layers) moisture content of the forest floor increased from 10% to 19%. Glowing combustion in deep duff fuels continued for 2 to 3 days; some large woody debris and stumps smoldered for up to 4 weeks. Forest floor material less than 1 inch (2.5 cm) in diameter was reduced 63%; material greater than 1 inch (2.5 cm) was reduced 69%. Mineral soil was exposed on about 19% of the area, mostly around large mature trees and rotten logs. Pre- and postfire fuel loads were as follows [251]:
| Dead fuels | Prefire fuel load (tons/acre) | Postfire fuel load (tons/acre) | fuel reduction (%) |
| Surface & ground fuels | |||
| needles/humus | 12.19 | 4.06 | 67 |
| 0 to 1/4th in. woody | 0.79 | 0.27 | 66 |
| 1-4th to 1-in. woody | 1.31 | 1.99 | 24 |
| other material | 0.88 | 1.35 | 60 |
| Total | 15.17 | 5.67 | 63 |
| Large woody fuel | |||
| 1 to 3-in. | 0.81 | 0.37 | 54 |
| > 3 in., sound | 2.09 | 1.79 | 14 |
| > 3 in., rotten | 4.26 | 0.04 | 99 |
| Total | 7.16 | 2.20 | 69 |
| Total, all dead fuels | 22.33 | 7.87 | 65 |
| Seedbed classification* | ||||||||||
| LFH | L | F | FH | H | M | A | CF | CH | Total | |
| unburned | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 |
| burned | 2 | 1 | 25 | 8 | 5 | 25 | 3 | 13 | 14 | 96 |
| *LFH=all 3 forest floor layers intact; no fire; L=newly cast needles on naturally bare soil, no fire; F=F layer intact but charred on top surface from burnt L layer, no H layer below; FH=F and H layers intact but charred on top surface from burnt L layer; H=H layer intact but charred on top surface from burnt L and F layers; M=mineral soil, all materials consumed; A=mineral soil, chunks of charcoal; CF=F layer heavily charred, L layer burnt off, no H layer below; CH=H layer heavily charred, L and F layers burnt off. | ||||||||||
Germination and initial establishment were excellent on burned areas despite below normal precipitation during the spring and summer of 1977. The enhanced survival under these conditions may have resulted from the improved moisture relations, increased nutrient availability, and more favorable soil temperatures associated with mineral soil seedbeds. However, subsequent mortality was quite high due to the combined effects of drought, frost heaving, animal damage, and interval burning (see below). By the fall of 1981, only 12% of tagged seedlings (25 on each of 18 plots treatment plots and 8 on controls) remained. Survival of 4-year-old tagged seedlings on plots subjected to interval burning was 6%; 19% of tagged seedlings survived on plots receiving the initial treatment only. Probable cause of death (by %) of seedlings selected 1 year after 1977 postfire germination is presented below [254].
| Rotation | Drought/ winter kill | Frost heaving | Fire | Browsing | Unknown/other |
| Control | 37 | 13 | NA | 0 | 50 |
| 1-yr burn rotation1 | 70 | 9 | 17 | 0 | 4 |
| 2-yr burn rotation2 | 25 | 10 | 56 | 3 | 6 |
| 4-yr burn rotation3 | 50 | 20 | 20 | 2 | 8 |
| 6-yr burn rotation4 | 52 | 13 | NA | 3 | 32 |
| 8-yr burn rotation4 | 48 | 28 | NA | 6 | 18 |
| 10-yr burn rotation4 | 67 | 9 | NA | 2 | 22 |
|
13 fires after seedling establishment. 22 fires after seedling establishment. 31 fire after seedling establishment. 4No fires after seedling establishment. |
|||||
Although very few sample seedlings were still alive by the fall of 1980, many 1977 seedlings remained on treatment plots. As a result, prior to interval burning in the fall of 1980, all seedlings on the 1-, 2-, and 4-year rotation plots were inventoried; a 100% inventory of 1977 seedlings on projected 6-, 8-, and 10-year rotations was done in the spring of 1981. Dry burning conditions in the fall of 1980 created more intense fires than had previously occurred during the study. Fireline intensities ranging between 260 to 650 btu/sec/fireline foot were produced on 4-year rotation plots, killing 94% of 3-year-old seedlings. Reduced fuel accumulations on annual and biennial plots produced less intense fires that resulted in 50% and 87% seedling mortality, respectively. Regardless of interval treatment, most surviving seedlings were located in areas of sparse or discontinuous fuels. Number of seedlings remaining from 1977 germination is presented below by treatment [254].
| Fall 1980 | Spring 1981 | Spring 1982 | |||||
| number | number | number | height, inches (mean) | height (range) | Stocking (#/acre) | ||
| Control | 0 | 0 | 0 | 0 | |||
| 1-yr burn rotation | 1431 | burned | 72 | 43 | 5.0 | 1.6-9.3 | 94 |
| 2-yr burn rotation | 3862 | burned | 49 | 43 | 5.9 | 2.1-15.3 | 24 |
| 4-yr burn rotation | 7253 | burned | 43 | 40 | 8.7 | 3.9-29.8 | 30 |
| 6-yr burn rotation | projected fire | 1952 | 222 | 5.6 | 1.9-17.1 | 110 | |
| 8-yr burn rotation | projected fire | 2462 | 283 | 4.6 | 1.7-27.1 | 120 | |
| 10-yr burn rotation | projected fire | 1832 | 165 | 5.1 | 1.5-14.4 | 130 | |
| 13 fires after seedling establishment. 21 fire after seedling establishment. 3No fires after seedling establishment. |
|||||||
Birds: Ponderosa pine communities are critical habitat for a wide variety of birds [32,39,120] including owls [113,120,168], other cavity nesters, and wild turkey [247]. Cavity-nesting birds use ponderosa pine snags for foraging and roosting as well as nesting [32,99]. Balda [32] estimated that secondary cavity nesters comprised 68% of the total density of wintering birds in southwestern ponderosa pine forests. Cunningham and others [83] provide a list of secondary cavity nesters on the Coconino National Forest of Arizona. They recommend a density of 5.2 ponderosa pine snags per hectare to sustain habitat for cavity-nesting birds [83].
Bennetts and Hawksworth [39] found that bird diversity and number of nests observed in Rocky Mountain ponderosa pine stands in northern Colorado were positively correlated with Rocky Mountainr ponderosa pine's level of infestation with southwestern pine dwarf-mistletoe (Arceuthobium vaginatum subsp. cryptopodum). Western tanagers, American robins, chipping sparrows, hermit thrush, and Cassin's finches nested in witch's brooms. Among guilds, number of insectivores, canopy, foliage, and ground omnivores, and ground granivores species was significantly higher in infected stands, while there were no significant differences in the number of flycatchers and nectivores due to level of stand infection. In northern Arizona, Dwyer [99] found that in postfire years 1 and 2, secondary cavity-nesters were more abundant in unburned, light-, and moderate-severity wildfire sites compared to sites that experienced severe wildfire. She attributed the difference to lack of suitable ponderosa pine snags on the severe burn. Secondary cavity-nesters used the severe burn when artificial nest boxes were attached to burned snags. In the short term, light and moderate wildfire promoted mountain bluebird and had little effect on pygmy nuthatches, white-breasted nuthatches, and violet-green swallows. Moderate-severity wildfire had a short-term negative effect on mountain chickadees. Other studies [169,287] have shown that mountain chickadees increase or return to prefire numbers 3 to 5 years after fire.
Southwestern ponderosa pine provides roosting, nesting, and foraging habitat for the Mexican spotted owl. The mixed-conifer, ponderosa pine, and ponderosa pine/Gambel oak types provide optimal habitat for the owl, which is federally listed as threatened [121]. Ganey and Balda [119] found that for roosting habitat, Mexican spotted owls in northern Arizona preferred closed-canopy, virgin stands of mixed conifers with a high density of snags and large logs. The owls preferred both ponderosa pine and mixed-conifer old-growth forests for foraging. A study on the Coconino National Forest showed the owls preferentially selected nest sites in areas with 41 to 70% canopy closure [129]. Fiedler and Cully [113] provide tree basal area and age class silvicultural prescriptions that optimize Mexican spotted owl habitat and promote old-growth ponderosa pine.
The flammulated owl, a cavity nester, is also dependent on old-growth southwestern ponderosa pine forest, but favors more open canopies for nesting and foraging than the Mexican spotted owl [168].
Reintroduced and rare in southeastern Arizona, thick-billed parrots are ecologically dependent on southwestern, Arizona, and Chihuahua pines for food and shelter. Pine seeds are their primary diet item, and the parrots nest in pine snag cavities [270].
Small mammals: Ponderosa pine provides habitat for many rodent species, and the seeds are an important food source for some rodents and shrews [126,182]. Tree squirrels (Sciurus and Tamiasciurus spp.) use ponderosa pines for nesting, and the seeds are among their most important foods. Abert's squirrel is ecologically dependent upon southwestern ponderosa pine [107,109,181,269]. Red squirrels beyond the range of Abert's squirrel use ponderosa pines heavily for food and nesting. Where ranges of the 2 squirrels overlap, Abert's squirrels tend to displace red squirrels to higher-elevation forests [109].
Deer: ponderosa pine communities provide valuable deer habitat. Their understories provide forage; production is highest in open stands. A study on the Apache-Sitgreaves National Forest of Arizona showed mean production of mule deer forage in ponderosa pine-Douglas-fir-southwestern white pine was 82 pounds/acre (92 kg/ha) at 50 feet2/acre (11 m2/ha) basal area and 4 pounds/acre (4.5 kg/ha) at 400 feet2/acre (90 m2/ha) basal area [291]. The tree itself provides minor browse [297]. White-tailed deer on Missouri River bottomlands of north-central Montana consumed Rocky Mountain ponderosa pine in trace amounts during fall. In winter, browsing frequency increased to 33%, comprising 2% of total volume intake. White-tailed deer were not detected browsing interior ponderosa pine in spring or summer [10].
PALATABILITY:Rodents and lagomorphs browse seedlings year-round [84]. Common porcupines consume the phloem of mature ponderosa pines; Abert's squirrels consume the seeds, buds, and twigs as well as the phloem. Palatability of the phloem varies among trees within a stand, with both rodent species preferring trees with relatively low monoterpene concentrations. Common porcupines prefer trees infected with southwestern pine dwarf-mistletoe to uninfected trees [201].
NUTRITIONAL VALUE:| Protein | Acid-detergent fiber | Ca | P | Digestible dry matter* |
| 8.0 | 41 | 0.13 | 0.15 | 41 |
Some exotic herbs that are commonly planted for rehabilitation may interfere with growth of ponderosa pine seedlings. Artificial ponderosa pine regeneration planted north of Flagstaff showed no significant difference in height and stem diameter (P<0.05) on plots seeded to native blue grama or bottlebrush squirreltail and then weeded of other species compared to control plots kept free of all species but ponderosa pine; however, ponderosa pine seedlings on plots seeded with the exotics small burnet (Sanguisorba minor), yellow sweetclover (Melilotus officinalis), orchard grass (Dactylis glomerata), or desert wheatgrass (Agropyron desertorum) were significantly shorter than seedlings on control plots or plots planted to the native grasses. Seedlings planted with yellow sweetclover or desert wheatgrass were significantly smaller in girth compared to those on weeded plots or plots seeded to other herbaceous species [105].
OTHER USES AND VALUES:Native Americans ate ponderosa pine seeds and the sweet, edible phloem in the inner bark [151,152]. The Cheyenne of Montana applied Rocky Mountain ponderosa pine pitch inside whistles and flutes to improve the instruments' tone. They made blue dye from a root extract [152]. The Nez Perce used the pitch as a torch fuel; the Nez Perce and Crow also used pitch as glue [151].
OTHER MANAGEMENT CONSIDERATIONS:Pests and diseases: Southwestern pine dwarf-mistletoe is a serious disease agent of ponderosa pines south of Wyoming [153,200]. Infection rates in the Southwest were at 33% in the 1990s [200], with about 20% infection of ponderosa pine on the Colorado Front Range. Stands that have had partial cuts or mountain pine beetle (Dendroctonus ponderosae) attacks are most susceptible to dwarf-mistletoe (Arceuthobium spp.) infections. New stands that have regenerated after stand-replacing fire are least likely to become infected [6]. Dwarf-mistletoes and ponderosa pine do not cooccur north of Colorado because the northern interior continental climate is inhospitable to dwarf-mistletoe growth [7,52].
At least 59 species of insects attack ponderosa pine. The mountain pine beetle is the most serious pest in the Black Hills and the central and southern Rocky Mountains; D. adjunctus is a closely related pine beetle that damages trees in the Southwest. Pine beetle epidemics have occurred throughout recorded history. Epidemic outbreaks are usually associated with large (>6-inch (15 cm) diameter), stressed tress in overcrowded stands [7,52]. Pine engraver beetles (Ips spp.) are usually secondary infesters in the central and southern Rocky Mountains but are often more damaging than pine beetles in the Southwest [7,227].
Bark beetle (Ips spp. and D. adjunctus) kill has escalated greatly in the Southwest. Prolonged drought and high tree density have probably increased the susceptibility of southwestern ponderosa pine to attacks. A 2002 survey of National Forests in Arizona and New Mexico showed a 4-fold increase in ponderosa pine mortality in 2002, with over 2 million pines killed on approximately a half-million acres. A combination of control actions including removing infested trees, application of insecticide, and thinning has been recommended. Because the beetle outbreaks can be extensive, control efforts are best targeted to accessible, high-value areas [14].
The most serious wood-decaying fungi are red rot and western gall rust. Shoestring root rot occasionally infects pole-sized and younger trees [7]. Armillaria (Armillaria spp.) is a serious fungal pathogen in the Black Hills and New Mexico [154]. Management and harvesting guidelines to minimize infection are available [6,7,154].
Age classes and stand structure: Ponderosa pine is rapidly losing its oldest age class. Mortality of old-growth ponderosa pine is high due to increased interference from dense younger trees and stagnated nutrient cycling in the absence of fire. For example, mortality rate of old-growth ponderosa pine on the Fort Valley Experimental Forest of Arizona increased 7-fold from the 1930s level (0.75 tree/ha/decade) to the 1970s (5.75 trees/ha/decade) [213]. Changes to interior ponderosa pine and mixed-conifer forests due to fire exclusion are discussed in the Fire Ecology section of this report.
Range: Open stands of ponderosa pine produce more forage than dense stands. Periodic surface fire promotes grass production by reducing shrubs and pine litter, which retard graminoid production. Postfire increases in forage production last about 5 to 10 years. Other benefits of range fires include better livestock distribution and range utilization, increased water yield, and improved wildlife habitat.
Forage productivity in southwestern ponderosa pine communities varies with understory, with blue grama understories generally the least productive and Arizona fescue and screwleaf muhly the most productive [228]. Stands of large-diameter trees in a patchy distribution help maintain graminoid diversity [70,225]. Overstory thinning increased production in the grass and forb understory on the Coconino National Forest. Herbage production was significantly* greater (P=0.10) on plots thinned to basal areas of less than 70 ft2/acre (16.3 m2/ha) compared to unthinned plots with similar basal areas. Herbage production 6 years after treatment was as follows [70]:
| Basal area (ft2/acre) | Difference (P=0.10) | ||
| thinned | unthinned | ||
| 20 | 566.6 | 376.8 | 189.8* |
| 40 | 384.2 | 277.1 | 107.1* |
| 60 | 277.5 | 218.8 | 58.7* |
| 80 | 201.8 | 177.4 | 24.4 |
| 100 | 143.0 | 145.2 | -2.2 |
| 120 | 95.1 | 119.0 | -23.9 |
Cattle and elk generally prefer oprn stands [71,72]. On thinned interior ponderosa pine sites in Arizona, elk preferred sites where some alligator juniper was left to sites where it had been completely removed. Mule deer showed no preference for thinned vs. unthinned areas, but tended to congregate on sites with alligator juniper, southwestern pine dwarf-mistletoe, or acorn-producing Gambel oak [72].
After monitoring cattle use on ponderosa pine/Arizona fescue-mountain muhly on the Manitou Experimental Forest near Colorado Springs, Colorado, Smith [268] recommended 30% to 40% utilization of the bunchgrasses by the end of the grazing season to maximize cattle weight gain while maintaining palatable bunchgrasses. Currie [84] provides suggestions for managing ponderosa pine/bunchgrass lands in the Central Rocky Mountains for cattle grazing.
Response of ponderosa pine to grazing: Light- to moderate-intensity grazing does not greatly impact advanced ponderosa pine regeneration on rangelands in good condition. On the Manitou Experimental Forest, neither rest-rotation nor season-long grazing resulted in much damage to natural and artificial regeneration under light or moderate use, but ponderosa pine seedlings were heavily browsed and incurred severe damage to terminal buds under heavy rest-rotation or season-long grazing use [84]. Thirteen years after exclosures were built, ponderosa pine in northern and central Arizona had increased slightly on both ungrazed and grazed plots, at 6% and 5% frequency, respectively. Initial frequency was 2% on both plots [26].
Mast and others [215] found that moderate cattle grazing combined with fire exclusion has favored ponderosa pine seedling establishment on ponderosa pine/grassland and ecotonal communities of the Colorado Front Range. Prior to fire exclusion, frequent fire probably excluded ponderosa pine seedlings at the ecotonal boundary [215] .1. Ahlstrand, Gary M. 1979. Preliminary report on the ecology of fire study, Guadalupe Mountains and Carlsbad Caverns National Parks. In: Genoways, Hugh H.; Baker, Robert J., eds. Biological investigations in the Guadalupe Mountains National Park: Proceedings of a symposium; 1975 April 4-5; Lubbock, TX. Proceedings and Transactions Series No. 4. Washington, DC: U.S. Department of the Interior, National Park Service: 31-44. [16015]
2. Ahlstrand, Gary M. 1980. Fire history of a mixed conifer forest in Guadalupe Mountains National Park. In: Stokes, Marvin A.; Dieterich, John H., technical coordinators. Proceedings of the fire history workshop; 1980 October 20-24; Tucson, AZ. Gen. Tech. Rep. RM-81. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 4-7. [16035]
3. Aldon, Earl F. 1968. Moisture loss and weight of the forest floor under pole-size ponderosa pine stands. Journal of Forestry. 66(1): 70-71. [37481]
4. Alexander, Billy G., Jr.; Fitzhugh, E. Lee; Ronco, Frank, Jr.; Ludwig, John A. 1987. A classification of forest habitat types of the northern portion of the Cibola National Forest, New Mexico. Gen. Tech. Rep. RM-143. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 35 p. [4207]
5. Alexander, Billy G., Jr.; Ronco, Frank, Jr.; Fitzhugh, E. Lee; Ludwig, John A. 1984. A classification of forest habitat types of the Lincoln National Forest, New Mexico. Gen. Tech. Rep. RM-104. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 29 p. [300]
6. Alexander, Robert R. 1986. Silvicultural systems and cutting methods for ponderosa pine forests in the Front Range of the central Rocky Mountains. Gen. Tech. Rep. RM-128. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 22 p. [13072]
7. Alexander, Robert R. 1987. Silvicultural systems, cutting methods, and cultural practices for Black Hills ponderosa pine. Gen. Tech. Rep. RM-139. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 32 p. [305]
8. Alexander, Robert R.; Hoffman, George R.; Wirsing, John M. 1986. Forest vegetation of the Medicine Bow National Forest in southeastern Wyoming: a habitat type classification. Res. Pap. RM-271. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 39 p. [307]
9. Allen, Craig D.; Savage, Melissa; Falk, Donald A.; Suckling, Kieran F.; Swetnam, Thomas W.; Schulke, Todd; Stacey, Peter B.; Morgan, Penelope; Hoffman, Martos; Klingel, Jon T. 2002. Ecological restoration of southwestern ponderosa pine ecosystems: a broad perspective. Ecological Applications. 12(5): 1418-1433. [43210]
10. Allen, Eugene O. 1968. Range use, foods, condition, and productivity of white-tailed deer in Montana. Journal of Wildlife Management. 32(1): 130-141. [16331]
11. Allen, Rogert B.; Peet, Robert K. 1990. Gradient analysis of forests of the Sangre de Cristo Range, Colorado. Canadian Journal of Botany. 68: 193-201. [11231]
12. Amstrup, Steven C. 1978. Activities and habitat use patterns of pronghorns on Montana-Wyoming coal lands. In: Proceedings, 8th biennial pronghorn antelope workshop; 1978 May 2-4; Jasper, AB. Edmonton, AB: Alberta Recreation, Parks, and Wildlife, Fish and Wildlife Division: 270-304. [3300]
13. Andariese, Steven W. 1982. Time-response graphs for understory production following fall prescribed burning in Arizona ponderosa pine on basalt soils. Flagstaff, AZ: Northern Arizona University. 42 p. Thesis. [20307]
14. Anhold, John. 2002. [Forest Service memo to Arizona Forest Supervisors]. October 29. 4 leaves. 2002 forest insect and disease aerial detection survey results. On file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; RWU 4403 files. [44430]
15. Armijo, Lou. 1979. Spare that old, dead tree! American Forests. 85(3): 32-35, 47. [15563]
16. Arno, Stephen F. 1979. Forest regions of Montana. Res. Pap. INT-218. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 39 p. [340]
17. Arno, Stephen F. 1980. Forest fire history in the Northern Rockies. Journal of Forestry. 78(8): 460-465. [11990]
18. Arno, Stephen F. 1988. Fire ecology and its management implications in ponderosa pine forests. In: Baumgartner, David M.; Lotan, James E., compilers. Ponderosa pine: The species and its management: Symposium proceedings; 1987 September 29 - October 1; Spokane, WA. Pullman, WA: Washington State University, Cooperative Extension: 133-139. [9410]
19. Arno, Stephen F. 1996. The concept: restoring ecological structure and process in ponderosa pine forests. In: Hardy, Colin C.; Arno, Stephen F., eds. The use of fire in forest restoration: A general session at the annual meeting of the Society for Ecological Restoration; 1995 September 4-16; Seattle, WA. Gen. Tech. Rep. INT-GTR-341. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 37-38. [28669]
20. Arno, Stephen F. 2000. Fire in western forest ecosystems. In: Brown, James K.; Smith, Jane Kapler, eds. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42-vol. 2. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 97-120. [36984]
21. Arno, Stephen F.; Gruell, George E. 1983. Fire history at the forest-grassland ecotone in southwestern Montana. Journal of Range Management. 36(3): 332-336. [342]
22. Arno, Stephen F.; Harrington, Michael G. 1995. Use thinning and fire to improve forest health and wildlife habitat. Tree Farmer. May/June: 6-8, 23. [26069]
23. Arno, Stephen F.; Harrington, Michael G.; Fiedler, Carl E.; Carlson, Clinton E. 1995. Restoring fire-dependent ponderosa pine forests in western Montana. Restoration and Management Notes. 13(1): 32-36. [27601]
24. Arno, Stephen F.; Wilson, Andrew E. 1986. Dating past fires in curlleaf mountain-mahogany communities. Journal of Range Management. 39(3): 241-243. [350]
25. Arnold, Joseph F. 1950. Changes in ponderosa pine bunchgrass ranges in northern Arizona resulting from pine regeneration and grazing. Journal of Forestry. February: 118-126. [352]
26. Arnold, Joseph F.; Jameson, Donald A.; Reid, Elbert H. 1964. The pinyon-juniper type of Arizona: effects of grazing, fire and tree control. Production Research Report No. 84. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 28 p. [353]
27. Aulenbach, Steve; O'Shea-Stone, Maureen. 1983. Bird utilization of a ponderosa pine forest after a fire. Colorado Field Ornithologists' Journal. 17(1): 14-19. [23658]
28. Avery, Charles C.; Larson, Frederic R.; Schubert, Gilbert H. 1976. Fifty-year records of virgin stand development in southwestern ponderosa pine. Gen. Tech. Rep. RM-22. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 71 p. [10989]
29. Bahre, Conrad J. 1998. Late 19th century human impacts on the woodlands and forests of southeastern Arizona's sky islands. Desert Plants. 14(1): 8-21. [28878]
30. Baisan, Christopher H.; Swetnam, Thomas W. 1990. Fire history on a desert mountain range: Rincon Mountain Wilderness, Arizona, U.S.A. Canadian Journal of Forest Research. 20: 1559-1569. [14986]
31. Baker, Frederick S. 1949. A revised tolerance table. Journal of Forestry. 47: 179-181. [20404]
32. Balda, Russell P. 1975. Vegetation structure and breeding bird diversity. In: Smith, Dixie R., technical coordinator. Proceedings of the symposium on management of forest and range habitats for nongame birds; 1975 May 6-9; Tucson, AZ. Gen. Tech. Rep. WO-1. Washington, DC: U.S. Department of Agriculture, Forest Service: 59-80. [17768]
33. Barrett, James W.; McDonald, Philip M.; Ronco, Frank, Jr.; Ryker, Russell A. 1980. Interior ponderosa pine. In: Eyer, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: U.S. Department of Agriculture, Forest Service: 114-115. [37242]
34. Barton, Andrew M. 1992. Factors controlling lower elevational limits of plants: responses of pines to drought in the Chiricahua Mountains, Arizona. In: Ffolliott, Peter F.; Gottfried, Gerald J.; Bennett, Duane A.; [and others], technical coordinators. Ecology and management of oak and associated woodlands: perspectives in the southwestern United States and northern Mexico: Proceedings; 1992 April 27-30; Sierra Vista, AZ. Gen. Tech. Rep. RM-218. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 191-194. [19764]
35. Barton, Andrew M.; Teeri, James A. 1993. The ecology of elevational positions in plants: drought resistance in five montane pine species in southwestern Arizona. American Journal of Botany. 80(1): 15-25. [20527]
36. Bates, Carlos G. 1923. Physiological requirements of Rocky Mountain trees. Journal of Agricultural Research. 24(2): 97-164. [10326]
37. Beaufait, William R. 1965. Characteristics of backfires and headfires in a pine needle fuel bed. Res. Note INT-39. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 7 p. [8099]
38. Bennett, Peter S.; Kunzmann, Michael R. 1992. The applicability of generalized fire prescriptions to burning of Madrean evergreen forest and woodland. Journal of the Arizona-Nevada Academy of Science. 24-25: 79-84. [18324]
39. Bennetts, Robert E.; Hawksworth, Frank G. 1991. The indirect effects of dwarf mistletoe on bird communities in Colorado ponderosa pine forests. In: Muir, John A., ed. Proceedings, 39th annual western international forest disease work conference; 1991 August 5-9; Victoria, BC. [Place of publication unknown]: [Publisher unknown]: 14-17. [25894]
40. Bernard, Stephen R.; Brown, Kenneth F. 1977. Distribution of mammals, reptiles, and amphibians by BLM physiographic regions and A.W. Kuchler's associations for the eleven western states. Tech. Note 301. Denver, CO: U.S. Department of the Interior, Bureau of Land Management. 169 p. [434]
41. Berndt, Herbert W.; Gibbons, Robert D. 1958. Root distribution of some native trees and understory plants growing on three sites within ponderosa pine watersheds in Colorado. Station Paper No. 37. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 14 p. [16337]
42. Biondi, Franco; Klemmedson, James O.; Kuehl, Robert O. 1992. Dendrochronological analysis of single-tree interactions in mixed pine-oak stands of central Arizona, USA. Forest Ecology and Management. 48: 321-333. [27611]
43. Biswell, Harold H.; Kallander, Harry R.; Komarek, Roy; [and others]. 1973. Ponderosa fire management. Misc. Publ. No. 2. Tallahassee, FL: Tall Timbers Research Station. 49 p. [4691]
44. Bjugstad, Ardell J. 1986. Wooded draws of the northern high plains: characteristics, value and restoration (North and South Dakota). Restoration & Management Notes. 4(2): 74-75. [4226]
45. Blackburn, Wilbert H.; Tueller, Paul T.; Eckert, Richard E., Jr. 1969. Vegetation and soils of the Pine and Mathews Canyon watersheds. Reno, NV: University of Nevada, Agricultural Experiment Station. 109 p. In cooperation with: U.S. Department of the Interior, Bureau of Land Management. [7437]
46. Blake, Elizabeth A.; Wagner, Michael R.; Koerber, Thomas W. 1989. Relative effect of seed and cone insects on ponderosa pine in northern Arizona. Journal of Economic Entomology. 82(6): 1691-1694. [20531]
47. Blake, John G. 1982. Influence of fire and logging on nonbreeding bird communities of ponderosa pine forests. Journal of Wildlife Management. 46(2): 404-415. [5114]
48. Blatner, Keith A.; Govett, Robert L. 1988. Ponderosa pine lumber market. In: Baumgartner, David M.; Lotan, James E., compilers. Ponderosa pine: The species and its management: Symposium proceedings; 1987 September 29 - October 1; Spokane, WA. Pullman, WA: Washington State University, Cooperative Extension: 7-9. [9396]
49. Bock, Carl E.; Bock, Jane H. 1983. Responses of birds and deer mice to prescribed burning in ponderosa pine. Journal of Wildlife Management. 47(3): 836-840. [476]
50. Bock, Jane H.; Bock, Carl E. 1984. Effects of fires on woody vegetation in the pine-grassland ecotone of the southern Black Hills. The American Midland Naturalist. 112(1): 35-42. [477]
51. Boldt, Charles E. 1970. Sequential thinnings boost productivity of a ponderosa pine stand in the Black Hills of South Dakota. Res. Note RM-172. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 7 p. [15775]
52. Boldt, Charles E.; Van Deusen, James L. 1974. Silviculture of ponderosa pine in the Black Hills: the status of our knowledge. Res. Pap. RM-124. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 45 p. [4055]
53. Bone, Steven D.; Klukas, Richard W. 1990. Prescribed fire in Wind Cave National Park. In: Alexander, M. E.; Bisgrove, G. F., technical coordinators. The art and science of fire management: Proceedings, 1st Interior West Fire Council annual meeting and workshop; 1988 October 24-27; Kananaskis Village, AB. Inf. Rep. NOR-X-309. Edmonton, AB: Forestry Canada, Northwest Region, Northern Forestry Centre: 297-302. [14145]
54. Boucher, Paul F.; Moody, Ronald D. 1998. The historical role of fire and ecosystem management of fires: Gila National Forest, New Mexico. In: Pruden, Teresa L.; Brennan, Leonard A., eds. Fire in ecosystem management: shifting the paradigm from suppression to prescription: Proceedings, Tall Timbers fire ecology conference; 1996 May 7-10; Boise, ID. No. 20. Tallahassee, FL: Tall Timbers Research Station: 374-379. [35663]
55. Bradley, Anne F.; Noste, Nonan V.; Fischer, William C. 1992. Fire ecology of forests and woodlands in Utah. Gen. Tech. Rep. INT-287. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 128 p. [18700]
56. Bragg, Thomas B. 1985. A preliminary fire history of the oak/pine bluff forest of northcentral Nebraska. In: Proceedings, 95th annual meeting of the Nebraska Academy of Sciences; [Date unknown]; Lincoln, NE. [Place of publication unknown]: [Publisher unknown]: 8. On file with: U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Fire Sciences Laboratory, Missoula, MT. [5790]
57. Brandegee, T. S. 1891. The vegetation of "burns". Zoe. 2: 118-122. [33056]
58. Bratton, Gerald F. 1990. Windbreak renovation studies--update, 1964-1989. In: Great Plains Agricultural Council, compiler. Windbreaks: Living with the wind: Proceedings, windbreak renovation workshop; 1990 October 23-25; Hutchinson, KS. Great Plains Agriculture Council Publ. No. 133. Manhattan, KS: Kansas State University, Cooperative Extension Service: 17-20. [15253]
59. Brawn, Jeffrey D.; Balda, Russell P. 1988. Population biology of cavity nesters in northern Arizona: do nest sites limit breeding densities? The Condor. 90: 61-71. [24926]
60. Brown, David E.; Lowe, Charles H.; Hausler, Janet F. 1977. Southwestern riparian communities: their biotic importance and management in Arizona. In: Johnson, R. Roy; Jones, Dale A., tech. coords. Importance, preservation and management of riparian habitat: a symposium: Proceedings; 1977 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment 201-211. [5348]
61. Brown, Peter M.; Kaufmann, Merrill R.; Shepperd, Wayne D. 1999. Long-term, landscape patterns of past fire events in a montane ponderosa pine forest of central Colorado. Landscape Ecology. 14(6): 513-532. [31161]
62. Brown, Peter M.; Ryan, Michael G.; Andrews, Thomas G. 2000. Historical surface fire frequency in ponderosa pine stands in Research Natural Areas, Central Rocky Mountains and Black Hills, USA. Natural Areas Journal. 20(2): 133-139. [35463]
63. Brown, Peter M.; Sieg, Carolyn H. 1999. Historical variability in fire at the ponderosa pine - Northern Great Plains prairie ecotone, southeastern Black Hills, South Dakota. Ecoscience. 6(4): 539-547. [35536]
64. Buchanan, Hayle; Tolman, Joseph Mark. 1983. The pre-historic fire regime of the forests of Bryce Canyon National Park, Utah. In: Fire history of the forests of Bryce Canyon National Park, UT. Final Report to the National Park Service: Contract No. PX 1330-3-0161. 40 p. Unpublished manuscript on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. [15602]
65. Burkhardt, Wayne J.; Tisdale, E. W. 1976. Causes of juniper invasion in southwestern Idaho. Ecology. 57: 472-484. [565]
66. Campbell, R. E.; Baker, M. B., Jr.; Ffolliott, P. F.; [and others]. 1977. Wildfire effects on a ponderosa pine ecosystem: an Arizona case study. Res. Pap. RM-191. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 12 p. [4715]
67. Carty, Dave. 1996. The drought busters. American Forests. 102(2): 33, 36-38. [27171]
68. Clary, Warren P. 1975. Range management and its ecological basis in the ponderosa pine type of Arizona: the status of our knowledge. Res. Pap. RM-158. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 35 p. [4688]
69. Clary, Warren P. 1987. Overview of ponderosa pine bunchgrass ecology and wildlife habitat enhancement with emphasis on southwestern United States. In: Fisser, Herbert G., ed. Wyoming shrublands: Proceedings, 16th Wyoming shrub ecology workshop; 1987 May 26-27; Sundance, WY. Laramie, WY: University of Wyoming, Department of Range Management, Wyoming Shrub Ecology Workshop: 11-21. [13913]
70. Clary, Warren P.; Ffolliott, Peter F. 1966. Differences in herbage-timber relationships between thinned and unthinned ponderosa pine stands. Res. Note RM-74. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 4 p. [4693]
71. Clary, Warren P.; Kruse, William H.; Larson, Frederic R. 1975. Cattle grazing and wood production with different basal areas of ponderosa pine. Journal of Range Management. 28(6): 434-437. [15823]
72. Clary, Warren P.; Larson, Frederic R. 1971. Elk and deer use are related to food sources in Arizona ponderosa pine. Res. Note RM-202. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 4 p. [15779]
73. Conkle, M. Thompson; Critchfield, William B. 1988. Genetic variation and hybridization of ponderosa pine. In: Baumgartner, David M.; Lotan, James E., compilers. Ponderosa pine: The species and its management: Symposium proceedings; 1987 September 29 - October 1; Spokane, WA. Pullman, WA: Washington State University, Cooperative Extension: 27-43. [9399]
74. Cooper, Charles F. 1960. Changes in vegetation, structure, and growth of southwestern pine forests since white settlement. Ecological Monographs. 30(2): 129-164. [3927]
75. Cooper, Charles F. 1961. Pattern in ponderosa pine forests. Ecology. 42(3): 493-499. [5780]
76. Covington, W. W.; Moore, M. M. 1992. Postsettlement changes in natural fire regimes: implications for restoration of old-growth ponderosa pine forests. In: Kaufmann, Merrill R.; Moir, W. H.; Bassett, Richard L., technical coordinators. Old-growth forests in the Southwest and Rocky Mountain regions: Proceedings of a workshop; 1992 March 9-13; Portal, AZ. Gen. Tech. Rep. RM-213. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 81-99. [19045]
77. Covington, W. Wallace; Fule, Peter Z.; Moore, Margaret M.; [and others]. 1997. Restoring ecosystem health in ponderosa pine forests of the Southwest. Journal of Forestry. 95(4): 23-29. [27618]
78. Covington, W. Wallace; Sackett, Stephen S. 1984. The effect of a prescribed burn in Southwestern ponderosa pine on organic matter and nutrients in woody debris and forest floor. Forest Science. 30(1): 183-192. [7624]
79. Covington, Wallace. 1996. Implementing adaptive ecosystem restoration in western long-needled pine forests. In: Covington, Wallace; Wagner, Pamela K., technical coordinators. Conference on adaptive ecosystem restoration and management: restoration of Cordilleran conifer landscapes of North America: Proceedings; 1996 June 6-8; Flagstaff, AZ. Gen. Tech. Rep. RM-GTR-278. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 44-48. [26922]
80. Critchfield, William B.; Allenbaugh, Gordon L. 1969. The distribution of Pinaceae in and near northern Nevada. Madrono. 20(1): 12-25. [714]
81. Critchfield, William B.; Little, Elbert L., Jr. 1966. Geographic distribution of the pines of the world. Misc. Publ. 991. Washington, DC: U.S. Department of Agriculture, Forest Service. 97 p. [20314]
82. Cronquist, Arthur; Holmgren, Arthur H.; Holmgren, Noel H.; Reveal, James L. 1972. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Vol. 1. New York: Hafner Publishing Company, Inc. 270 p. [717]
83. Cunningham, James B.; Balda, Russell P.; Gaud, William S. 1980. Selection and use of snags by secondary cavity-nesting birds of the ponderosa pine forest. Res. Pap. RM-222. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 15 p. [15540]
84. Currie, Pat O. 1975. Grazing management of ponderosa pine-bunchgrass ranges of the central Rocky Mountains. Res. Pap. RM-159. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 24 p. [12600]
85. Danzer, Shelley R.; Baisan, Chris H.; Swetnam, Thomas W. 1996. The influence of fire and land-use history on stand dynamics in the Huachuca Mountains of southeastern Arizona. In: Ffolliott, Peter F.; DeBano, Leonard F.; Baker, Malchus, B., Jr.; [and others], tech. coords. Effects of fire on Madrean Province ecosystems: a symposium proceedings; 1996 March 11-15; Tucson, AZ. Gen. Tech. Rep. RM-GTR-289. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 265-270. [28111]
86. Daubenmire, R. 1960. A seven-year study of cone production as related to xylem layers and temperature in Pinus ponderosa. The American Midland Naturalist. 64(1): 187-193. [37425]
87. Daubenmire, R. F. 1943. Vegetational zonation in the Rocky Mountains. Botanical Review. 9(6): 326-393. [737]
88. Daubenmire, Rexford. 1953. Notes on the vegetation of forested regions of the far northern Rockies and Alaska. Northwest Science. 27: 125-138. [10816]
89. Davis, James R.; Ffolliott, Peter F.; Clary, Warren P. 1968. A fire prescription for consuming ponderosa pine duff. Res. Note RM-115. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 4 p. [11988]
90. Denyes, H. Arliss. 1956. Natural terrestrial communities of Brewster County, Texas. The American Midland Naturalist. 55(2): 289-320. [26765]
91. DeVelice, Robert L.; Ludwig, John A.; Moir, William H.; Ronco, Frank, Jr. 1986. A classification of forest habitat types of northern New Mexico and southern Colorado. Gen. Tech. Rep. RM-131. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 59 p. [781]
92. Dick-Peddie, William A.; Alberico, Michael S. 1977. Fire ecology study of the Chisos Mountains, Big Bend National Park, Texas: Phase I. CDRI Contribution No. 35. Alpine, TX: The Chihuahuan Desert Research Institute. 47 p. [5002]
93. Dieterich, J. H.; Hibbert, A. R. 1990. Fire history in a small ponderosa pine stand surrounded by chaparral. In: Krammes, J. S., technical coordinator. Effects of fire management of southwestern natural resources: Proceedings of the symposium; 1988 November 15-17; Tucson, AZ. Gen, Tech, Rep. RM-191. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 168-173. [11285]
94. Dieterich, John H. 1979. Recovery potential of fire-damaged Southwestern ponderosa pine. Res. Note RM-379. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 8 p. [4698]
95. Dieterich, John H. 1980. Chimney Spring Forest fire history. Res. Pap. RM-220. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 8 p. [5222]
96. Dieterich, John H.; Swetnam, Thomas W. 1984. Dendrochronology of a fire-scarred ponderosa pine. Forest Science. 30: 238-247. [5221]
97. Dodge, Richard Archie. 1963. Investigations into the ecological relationships of ponderosa pine in southwest Arizona. Tucson, AZ: University of Arizona. 118 p. Dissertation. [20315]
98. Duffield, J. W. 1953. Pine pollen collection dates--annual and geographic variation. For. Res. Notes No. 85. Berkeley, CA: U.S. Department of Agriculture, Forest Service, California Forest and Range Experiment Station. 9 p. [17970]
99. Dwyer, Jill K. 2000. Response of secondary cavity-nesting birds to wildlife in ponderosa pine forests of northern Arizona. Flagstaff, AZ: Northern Arizona University. 59 p. M.S. thesis. [15570]
100. Eakle, T. W.; Wagle, R. F. 1979. Method of estimating ground fuels under two inches in diameter in southwestern ponderosa pine stands. Res. Note PNW-33. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 6 p. [20864]
101. Eakle, Thomas William. 1976. The effects of controlled burning on a Southwestern ponderosa pine understory: a factor analysis. Tucson, AZ: University of Arizona. 124 p. Dissertation. [10454]
102. Easterly, Thomas G.; Jenkins, Kurt J. 1991. Forage production and use on bighorn sheep winter range following spring burning in grassland and ponderosa pine habitats. Prairie Naturalist. 23(4): 193-200. [19277]
103. Edminster, Carleton B.; Olsen, William K. 1996. Thinning as a tool in restoring and maintaining diverse structure in stands of Southwestern ponderosa pine. In: Covington, Wallace; Wagner, Pamela K., technical coordinators. Conference on adaptive ecosystem restoration and management: restoration of Cordilleran conifer landscapes of North America: Proceedings; 1996 June 6-8; Flagstaff, AZ. Gen. Tech. Rep. RM-GTR-278. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 62-68. [26925]
104. Eichhorn, Larry C.; Watts, C. Robert. 1984. Plant succession on burns in the river breaks of central Montana. Proceedings, Montana Academy of Science. 43: 21-34. [15478]
105. Elliott, Katherine J.; White, Alan S. 1987. Competitive effects of various grasses and forbs on ponderosa pine seedlings. Forest Science. 33(2): 356-366. [860]
106. Eyre, F. H., ed. 1980. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters. 148 p. [905]
107. Farentinos, R. C. 1972. Nests of the tassel-eared squirrel. Journal of Mammalogy. 53(4): 900-903. [25360]
108. Feeney, Shelly R.; Kolb, Thomas E.; Covington, W. Wallace; Wagner, Michael R. 1998. Influence of thinning and burning restoration treatments on presettlement ponderosa pines at the Gus Pearson Natural Area. Canadian Journal of Forestry. 28: 1295-1306. [29464]
109. Ferner, John W. 1974. Habitat relationships of Tamiasciurus hudsonicus and Sciurus aberti in the Rocky Mountains. The Southwestern Naturalist. 18: 470-473. [25245]
110. Ffolliott, Peter F.; Clary, Warren P.; Davis, James R. 1968. Some characteristics of the forest floor under ponderosa pine in Arizona. Research Note RM-127. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 8 p. [37482]
111. Ffolliott, Peter F.; Clary, Warren P.; Larson, Frederic R. 1976. Fire: 11 years after. Progressive Agriculture in Arizona. 28(1): 12-13. [34533]
112. Ffolliott, Peter F.; Larson, Frederic K.; Thill, Ronald E. 1977. Some characteristics of Arizona's mixed conifer forest floor. Res. Note RM-342. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 4 p. [8172]
113. Fielder, Carl E.; Cully, Jack F., Jr. 1995. A silvicultural approach to develop Mexican spotted owl habitat in Southwest forests. Western Journal of Applied Forestry. 10(4): 144-148. [26027]
114. Fitzhugh, E. Lee; Moir, William H.; Ludwig, John A.; Ronco, Frank, Jr. 1987. Forest habitat types in the Apache, Gila, and part of the Cibola National Forests, Arizona and New Mexico. Gen. Tech. Rep. RM-145. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 116 p. [4206]
115. Flint, H. R. 1925. Fire resistance of northern Rocky Mountain conifers. U.S. Department of Agriculture, Forest Service, Applied Forestry Note 61. Missoula, MT: Northern Rocky Mountain Forest Experiment Station. 5 p. [34634]
116. Flora of North America Association. 2000. Flora of North America north of Mexico. Volume 2: Pteridophytes and gymnosperms, [Online]. Available: http://hua.huh.harvard.edu/FNA/ [2002, March 27]. [36990]
117. Fule, Peter Z.; Covington, W. Wallace; Moore, Margaret M. 1997. Determining reference conditions for ecosystem management of southwestern ponderosa pine forests. Ecological Applications. 7(3): 895-908. [28496]
118. Gallina, Sonia; Ffolliott, Peter F. 1983. Overstory-understory relationships: oak-pine forests of Sierra Madre Occidental, Mexico. In: Bartlett, E. T.; Betters, David R., eds. Overstory-understory relationships in western forests. Western Regional Res. Publ. No. 1. Fort Collins, CO: Colorado State University Experiment Station: 19-20. [3312]
119. Ganey, Joseph L.; Balda, Russell P. 1994. Habitat selection by Mexican spotted owls in northern Arizona. Auk. 111(1): 162-169. [24368]
120. Ganey, Joseph L.; Block, William M.; Boucher, Paul F. 1996. Effects of fire on birds in Madrean forests and woodlands. In: Ffolliott, Peter F.; DeBano, Leonard F.; Baker, Malchus, B., Jr.; [and others], tech. coords. Effects of fire on Madrean Province ecosystems: a symposium proceedings; 1996 March 11-15; Tucson, AZ. Gen. Tech. Rep. RM-GTR-289. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 146-154. [28072]
121. Ganey, Joseph L.; Block, William M.; Jenness, Jeffrey S.; Wilson, Randolph A. 1999. Mexican spotted owl home range and habitat use in pine--oak forest: implications for forest management. Forest Science. 45(1): 127-135. [29463]
122. Garrison, George A.; Bjugstad, Ardell J.; Duncan, Don A.; Lewis, Mont E.; Smith, Dixie R. 1977. Vegetation and environmental features of forest and range ecosystems. Agric. Handb. 475. Washington, DC: U.S. Department of Agriculture, Forest Service. 68 p. [998]
123. Gartner, F. Robert; Thompson, Wesley W. 1973. Fire in the Black Hills forest-grass ecotone. In: Proceedings, annual Tall Timbers fire ecology conference; 1972 June 8-9; Lubbock, TX. No. 12. Tallahassee, FL: Tall Timbers Research Station: 37-68. [1002]
124. Gehlbach, Frederick R. 1979. Biomes of the Guadalupe Escarpment: vegetation, lizards, and human impact. In: Genoways, Hugh H.; Baker, Robert J., eds. Biological investigations in the Guadalupe Mountains National Park: Proceedings of a symposium; 1975 April 4-5; Lubbock, TX. Proceedings and Transactions Series No. 4. Washington, DC: U.S. Department of the Interior, National Park Service: 427-439. [16024]
125. Girard, Michele M.; Goetz, Harold; Bjugstad, Ardell J. 1989. Native woodland habitat types of southwestern North Dakota. Res. Pap. RM-281. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 36 p. [6319]
126. Goodwin, John G., Jr.; Hungerford, C. Roger. 1979. Rodent population densities and food habits in Arizona ponderosa pine forests. Res. Pap. RM-214. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 12 p. [15888]
127. Gottfried, Gerald J. 1978. Five-year growth and development in a virgin Arizona mixed conifer stand. Res. Pap. RM-203. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 22 p. [15661]
128. Grier, Bob. 1981. Burning with a purpose. Nebraskaland. 59(3): 38-42. [15545]
129. Grubb, Teryl G.; Ganey, Joseph L.; Masek, Sharon R. 1997. Canopy closure around nest sites of Mexican spotted owls in northcentral Arizona. Journal of Wildlife Management. 61(2): 336-342. [28909]
130. Gruell, G. E.; Loope, L. L. 1974. Relationships among aspen, fire, and ungulate browsing in Jackson Hole, Wyoming. Lakewood, CO: U.S. Department of the Interior, National Park Service, Rocky Mountain Region. 33 p. In cooperation with: U.S. Department of Agriculture, Forest Service, Intermountain Region. [3862]
131. Gruell, George E. 1985. Fire on the early western landscape: an annotated record of wildland fire. Northwest Science. 59(2): 97-107. [15660]
132. Grulke, Nancy E.; Retzlaff, William A. 2001. Changes in physiological attributes of ponderosa pine from seedling to mature tree. Tree Physiology. 21(5): 275-286. [37141]
133. Haase, Sally M. 1986. Effect of prescribed burning on soil moisture and germination of southwestern ponderosa pine seed on basaltic soils. Res. Note RM-462. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 6 p. [4741]
134. Hadley, Elmer B. 1969. Physiological ecology of Pinus ponderosa in southwestern North Dakota. The American Midland Naturalist. 81(2): 289-314. [4900]
135. Hanks, Jess P.; Dick-Peddie, W. A. 1974. Vegetation patterns of the White Mountains, New Mexico. The Southwestern Naturalist. 18(4): 371-382. [4635]
136. Hanks, Jess P.; Fitzhugh, E. Lee; Hanks, Sharon R. 1983. A habitat type classification system for ponderosa pine forests of northern Arizona. Gen. Tech Rep. RM-97. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 22 p. [1072]
137. Hansen, Paul L.; Hoffman, George R. 1988. The vegetation of the Grand River/Cedar River, Sioux, and Ashland Districts of the Custer National Forest: a habitat type classification. Gen. Tech. Rep. RM-157. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 68 p. [771]
138. Hansen, Paul L.; Hoffman, George R.; Steinauer, Gerry A. 1984. Upland forest and woodland habitat types of the Missouri Plateau, Great Plains Province. In: Noble, Daniel L.; Winokur, Robert P., eds. Wooded draws: characteristics and values for the Northern Great Plains: Symposium proceedings; 1984 June 12-13; Rapid City, SD. Great Plains Agricultural Council Publ. No. 111. Rapid City, SD: South Dakota School of Mines and Technology, Biology Department: 15-26. [1078]
139. Hansen, Paul L.; Pfister, Robert D.; Boggs, Keith; [and others]. 1995. Classification and management of Montana's riparian and wetland sites. Miscellaneous Publication No. 54. Missoula, MT: The University of Montana, School of Forestry, Montana Forest and Conservation Experiment Station. 646 p. [24768]
140. Hardy, Colin C.; Arno, Stephen F., eds. 1996. The use of fire in forest restoration: A general session at the annual meeting of the Society for Ecological Restoration; 1995 September 14-16; Seattle, WA. Gen. Tech. Rep. INT-GTR-341. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 86 p. [26833]
141. Harrington, M. G. 1985. The effects of spring, summer, and fall burning on Gambel oak in a southwestern ponderosa pine stand. Forest Science. 31(1): 156-163. [1092]
142. Harrington, M. G. 1993. Predicting Pinus ponderosa mortality from dormant season and growing season injury. International Journal of Wildland Fire. 3(2): 65-72. [21360]
143. Harrington, Michael G. 1986. Comparison of forest floor depth to loading relationships from several Arizona ponderosa pine stands. Res. Note RM-463. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 5 p. [20327]
144. Harrington, Michael G. 1987. Ponderosa pine mortality from spring, summer, and fall crown scorching. Western Journal of Applied Forestry. 2: 14-16. [4978]
145. Harrington, Michael G. 1987. Predicting reduction of natural fuels by prescribed burning under ponderosa pine in southeastern Arizona. Res. Note RM-472. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 4 p. [4725]
146. Harrington, Michael G. 1996. Fall rates of prescribed fire-killed ponderosa pine. Res. Pap. INT-RP-489. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 7 p. [27118]
147. Harrington, Michael G. 1996. Prescribed fire applications: restoring ecological structure and process in ponderosa pine forests. In: Hardy, Colin C.; Arno, Stephen F., eds. The use of fire in forest restoration: A general session of the Society for Ecological Restoration; 1995 September 14-16; Seattle, WA. Gen. Tech. Rep. INT-GTR-341. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 41. [26811]
148. Harrington, Michael G.; Hawksworth, Frank G. 1990. Interactions of fire and dwarf mistletoe on mortality of southwestern ponderosa pine. In: Krammes, J. S., technical coordinator. Effects of fire management of southwestern natural resources: Proceedings of the symposium; 1988 November 15-17; Tucson, AZ. Gen. Tech. Rep. RM-191. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 234-240. [11296]
149. Harrington, Michael G.; Sackett, Stephen S. 1992. Past and present fire influences on Southwestern ponderosa pine old growth. In: Kaufmann, Merrill R.; Moir, W. H.; Bassett, Richard L., technical coordinators. Old-growth forests in the Southwest and Rocky Mountain regions: Proceedings of a symposium; 1992 March 9-13; Portal, AZ. Gen. Tech. Rep. RM-213. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 44-50. [21977]
150. Harris, Gary R.; Covington, W. Wallace. 1983. The effect of a prescribed fire on nutrient concentration and standing crop of understory vegetation in ponderosa pine. Canadian Journal of Forest Research. 13: 501-507. [4699]
151. Hart, J. 1976. Montana--native plants and early peoples. Helena, MT: Montana Historical Society. 75 p. [9979]
152. Hart, Jeffrey A. 1981. The ethnobotany of the northern Cheyenne Indians of Montana. Journal of Ethnopharmacology. 4: 1-55. [35893]
153. Hawksworth, Frank G. 1961. Dwarfmistletoe of ponderosa pine in the Southwest. Technical Bulletin No. 1246. Washington, DC: U.S. Department of Agriculture, Forest Service. 112 p. [37485]
154. Hawksworth, Frank G.; Shaw, Charles G., III. 1988. Damage and control of major diseases of ponderosa pine. In: Baumgartner, David M.; Lotan, James E., compilers. Ponderosa pine: The species and its management: Symposium proceedings; 1987 September 29 - October 1; Spokane, WA. Pullman, WA: Washington State University, Cooperative Extension: 99-108. [9405]
155. Heidmann, L. J. 1988. Regeneration strategies for ponderosa pine. In: Baumgartner, David M.; Lotan, James E., compilers. Ponderosa pine: The species and its management: Symposium proceedings; 1987 September 29 - October 1; Spokane, WA. Pullman, WA: Washington State University, Cooperative Extension: 227-233. [9422]
156. Henrickson, James; Johnston, Marshall C. 1986. Vegetation and community types of the Chihuahuan Desert. In: Barlow, Jon C.; Powell, A. Michael; Timmermann, Barbara N. eds. Chihuahuan Desert--U.S. and Mexico, II: Proceedings of the 2nd symposium on resources of the Chihuahuan Desert region; 1983 October 20-21; Alpine, TX. Alpine, TX: Sul Ross State University, Chihuahuan Desert Research Institute: 20-39. [12979]
157. Herman, F. R. 1950. Survival of fire-damaged ponderosa pine. Research Note No. 119. Tucson, AZ: U.S. Department of Agriculture, Forest Service, Southwestern Forest and Range Experiment Station. 3 p. [4705]
158. Herman, F. R. 1954. A guide for marking fire-damaged ponderosa pine in the Southwest. Res. Notes No. 13. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 4 p. [37428]
159. Hess, Karl; Alexander, Robert R. 1986. Forest vegetation of the Arapaho and Roosevelt National Forests in central Colorado: a habitat type classification. Res. Pap. RM-266. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 48 p. [1141]
160. Hess, Karl; Wasser, Clinton H. 1982. Grassland, shrubland, and forestland habitat types of the White River-Arapaho National Forest. Final Report. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 335 p. [1142]
161. Higgins, Kenneth F. 1984. Lightning fires in North Dakota grasslands and in pine-savanna lands of South Dakota and Montana. Journal of Range Management. 37(2): 100-103. [1148]
162. Hill, Ralph R. 1946. Palatability ratings of Black Hills plants for white-tailed deer. Journal of Wildlife Management. 10(1): 47-54. [3270]
163. Hitchcock, C. Leo; Cronquist, Arthur; Ownbey, Marion. 1969. Vascular plants of the Pacific Northwest. Part 1: Vascular cryptogams, gymnosperms, and monocotyledons. Seattle, WA: University of Washington Press. 914 p. [1169]
164. Hoffman, George R.; Alexander, Robert R. 1976. Forest vegetation of the Bighorn Mountains, Wyoming: a habitat type classification. Res. Pap. RM-170. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 38 p. [1180]
165. Hoffman, George R.; Alexander, Robert R. 1983. Forest vegetation of the White River National Forest in western Colorado: a habitat type classification. Res. Pap. RM-249. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 36 p. [1178]
166. Hoffman, George R.; Alexander, Robert R. 1987. Forest vegetation of the Black Hills National Forest of South Dakota and Wyoming: a habitat type classification. Res. Pap. RM-276. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 48 p. [1181]
167. Horton, Scott P.; Mannan, R. William. 1988. Effects of prescribed fire on snags and cavity-nesting birds in southeastern Arizona pine forests. Wildlife Society Bulletin. 16: 37-44. [5549]
168. Illg, Cathy; Illg, Gordon. 1994. The ponderosa and the flammulated. American Forests. 100(3&4): 36-37, 58. [22791]
169. Johnson, Terrell H.; Wauer, Roland H. 1996. Avifaunal response to the 1977 La Mesa Fire. In: Allen, Craig D., ed. Fire effects in Southwestern forests: Proceedings, 2nd La Mesa fire symposium; 1994 March 29-31; Los Alamos, NM. RM-GTR-286. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 70-94. [27284]
170. Johnston, Barry C. 1987. Plant associations of Region Two: Potential plant communities of Wyoming, South Dakota, Nebraska, Colorado, and Kansas. 4th ed. R2-ECOL-87-2. Lakewood, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Region. 429 p. [3519]
171. Jones, John R. 1967. Regeneration of mixed conifer clearcuttings on the Apache National Forest, Arizona. Res. Note RM-79. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 8 p. [16082]
172. Jones, John R. 1974. Silviculture of southwestern mixed conifers and aspen: The status of our knowledge. Res. Pap. RM-122. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 44 p. [16081]
173. Jones, Stanley D.; Wipff, Joseph K.; Montgomery, Paul M. 1997. Vascular plants of Texas. Austin, TX: University of Texas Press. 404 p. [28762]
174. Kaib, Mark; Baisan, Christopher H.; Grissino-Mayer, Henri D.; Swetnam, Thomas W. 1996. Fire history in the gallery pine-oak forests and adjacent grasslands of the Chiricahua Mountains of Arizona. In: Ffolliott, Peter F.; DeBano, Leonard F.; Baker, Malchus, B., Jr.; [and others], tech. coords. Effects of fire on Madrean Province ecosystems: a symposium proceedings; 1996 March 11-15; Tucson, AZ. Gen. Tech. Rep. RM-GTR-289. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 253-264. [28109]
175. Kallander, Harry. 1969. Controlled burning on the Fort Apache Indian Reservation, Arizona. In: Proceedings, annual Tall Timbers fire ecology conference; 1969 April 10-11; Tallahassee, FL. No. 9. Tallahassee, FL: Tall Timbers Research Station: 241-249. [19351]
176. Kantak, Gail E. 1995. Terrestrial plant communities of the middle Niobrara Valley, Nebraska. The Southwestern Naturalist. 40(2): 129-138. [26698]
177. Kartesz, J. T.; The Biota of North America Program (BONAP). 2015. Taxonomic Data Center, [Online]. Chapel Hill, NC: The Biota of North America Program (Producer). Available online: bonap.org. [maps generated from Kartesz, J. T. 2010. Floristic synthesis of North America, Version 1.0. Biota of North America Program (BONAP). (in press)]. [84789]
178. Keane, Robert E.; Arno, Stephen F.; Brown, James K. 1990. Simulating cumulative fire effects in ponderosa pine/Douglas-fir forests. Ecology. 71(1): 189-203. [11517]
179. Kearney, Thomas H.; Peebles, Robert H.; Howell, John Thomas; McClintock, Elizabeth. 1960. Arizona flora. 2d ed. Berkeley, CA: University of California Press. 1085 p. [6563]
180. Keeley, Jon E.; Zedler, Paul H. 1998. Evolution of life histories in Pinus. In: Richardson, David M., ed. Ecology and biogeography of Pinus. Cambridge, United Kingdom: The Press Syndicate of the University of Cambridge: 219-250. [37705]
181. Keith, James O. 1965. The Abert squirrel and its dependence on ponderosa pine. Ecology. 46: 150-163. [1320]
182. Keyes, Christopher R. 2000. Natural regeneration of ponderosa pine: Pest management strategies for seed predators. The Forestry Chronicle. 76(4): 623-626. [35894]
183. Klemmedson, James O. 1987. Influence of oak in pine forests of central Arizona on selected nutrients of forest floor and soil. Soil Science Society of America Journal. 51(6): 1623-1628. [3041]
184. Knowles, Peggy; Grant, Michael C. 1983. Age and size structure analyses of Engelmann spruce, ponderosa pine, lodgepole pine, and limber pine in Colorado. Ecology. 64(1): 1-9. [1366]
185. Komarek, E. V., Sr. 1968. The nature of lightning fires. In: Proceedings, California Tall Timbers fire ecology conference; 1967 November 9-10; Hoberg, CA. No. 7. Tallahassee, FL: Tall Timbers Research Station: 5-41. [18442]
186. Komarkova, Vera. 1986. Habitat types on selected parts of the Gunnison and Uncompahgre National Forests. Final report: Contract No. 28-K2-234. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 270 p. [1369]
187. Komarkova, Vera; Alexander, Robert R.; Johnston, Barry C. 1988. Forest vegetation of the Gunnison and parts of the Uncompahgre National Forests: a preliminary habitat type classification. Gen. Tech. Rep. RM-163. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 65 p. [5798]
188. Krugman, Stanley L.; Jenkinson, James L. 1974. Pinus L. pine. In: Schopmeyer, C. S., tech. cood. Seeds of woody plants in the United States. Agric. Handb. 450. Washington, D.C.: U.S. Department of Agriculture, Forest Service: 598-638. [37725]
189. Kuchler, A. W. 1964. United States [Potential natural vegetation of the conterminous United States]. Special Publication No. 36. New York: American Geographical Society. 1:3,168,000; colored. [3455]
190. Kufeld, Roland C.; Bowden, David C.; Schrupp, Donald L. 1988. Habitat selection and activity patterns of female mule deer in the Front Range, Colorado. Journal of Range Management. 41(6): 515-522. [5719]
191. Lackschewitz, Klaus. 1991. Vascular plants of west-central Montana--identification guidebook. Gen. Tech. Rep. INT-227. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 648 p. [13798]
192. Lanner, Ronald M. 1983. Trees of the Great Basin: A natural history. Reno, NV: University of Nevada Press. 215 p. [1401]
193. Lanner, Ronald M. 1998. Seed dispersal in Pinus. In: Richardson, David M., ed. Ecology and biogeography of Pinus. Cambridge, United Kingdom: The Press Syndicate of the University of Cambridge: 281-295. [37707]
194. Lanner, Ronald M.; Vander Wall, Stephen B. 1980. Dispersal of limber pine seed by Clark's nutcracker. Journal of Forestry. 78(10): 637-639. [1410]
195. Larson, M. M. 1961. Seed size, germination dates, and survival relationships of ponderosa pine in the Southwest. Research Note No. 66. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 4 p. [5044]
196. Larson, M. M.; Schubert, Gilbert H. 1969. Effect of osmotic water stress on germination and initial development of ponderosa pine seedlings. Forest Science. 15(1): 30-36. [4712]
197. Larson, M. M.; Schubert, Gilbert H. 1970. Cone crops of ponderosa pine in central Arizona, including the influence of Abert squirrels. Res. Pap. RM-58. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 15 p. [4714]
198. Laven, R. D.; Omi, P. N.; Wyant, J. G.; Pinkerton, A. S. 1980. Interpretation of fire scar data from a ponderosa pine ecosystem in the central Rocky Mountains, Colorado. In: Stokes, Marvin A.; Dieterich, John H., technical coordinators. Proceedings of the fire history workshop; 1980 October 20-24; Tucson, AZ. Gen. Tech. Rep. RM-81. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 46-49. [7183]
199. Layser, Earle F.; Schubert, Gilbert H. 1979. Preliminary classification for the coniferous forest and woodland series of Arizona and New Mexico. Res. Pap. RM-208. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 27 p. [1428]
200. Lightle, Paul C.; Weiss, Melvyn J. 1974. Dwarf mistletoe of ponderosa pine in the Southwest. Forest Pest Leaflet 19. Washington, D.C.: U.S. Department of Agriculture, Forest Service. 8 p. [1454]
201. Linhart, Yan B.; Snyder, Marc A.; Habeck, Susan A. 1989. The influence of animals on genetic variability within ponderosa pine stands, illustrated by the effects of Abert's squirrel and porcupine. In: Multiresource management of ponderosa pine: Proceedings of a conference; 1989 November 14-16; Flagstaff, AZ. Gen. Tech. Rep. RM-185. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 141-148. [25364]
202. Little, Elbert L., Jr. 1971. Atlas of the United States trees. Volume 1. Conifers and important hardwoods. Misc. Publ. 1146. Washington, DC: U.S. Department of Agriculture, Forest Service. 320 p. [1462]
203. Lowe, Philip O.; Ffolliott, Peter F.; Dieterich, John H.; Patton, David R. 1978. Determining potential wildlife benefits from wildfire in Arizona ponderosa pine forests. Gen. Tech. Rep. RM-52. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 12 p. [4481]
204. Lundquist, J. E. 1995. Characterizing disturbance in managed ponderosa pine stands in the Black Hills. Forest Ecology and Management. 74: 61-74. [26673]
205. Lundquist, John E.; Negron, Jose F. 2000. Endemic forest disturbances and stand structure of ponderosa pine (Pinus ponderosa) in the Upper Pine Creek Research Natural Area, South Dakota, USA. Natural Areas Journal. 20(2): 126-132. [35462]
206. Lynch, Dennis L.; Romme, William H.; Floyd, M. Lisa. 2000. Forest restoration in southwestern ponderosa pine. Journal of Forestry. 98(8): 17-24. [36307]
207. Lynch, Donald W. 1959. Effects of a wildfire on mortality and growth of young ponderosa pine trees. Res. Note No. 66. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 8 p. [4748]
208. Mackie, Richard J. 1970. Range ecology and relations of mule deer, elk, and cattle in the Missouri River Breaks, Montana. Wildlife Monographs No. 20. 79 p. [5897]
209. Madany, Michael H.; West, Neil E. 1980. Fire history of two montane forest areas of Zion National Park. In: Stokes, Marvin A.; Dieterich, John H., technical coordinators. Proceedings of the fire history workshop; 1980 October 20-24; Tucson, AZ. Gen. Tech. Rep. RM-81. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 50-56. [16042]
210. Maguire, William P. 1956. Are ponderosa pine cone crops predictable? Journal of Forestry. 54: 778-779. [37483]
211. Martin, Robert E.; Anderson, Hal E.; Boyer, William D.; [and others]. 1979. Effects of fire on fuels: A state-of-knowledge review. Gen. Tech. Rep. WO-13. Washington, DC: U.S. Department of Agriculture, Forest Service. 64 p. [Prepared for: National fire effects workshop; 1978 April 10-14; Denver, CO]. [28838]
212. Martin, William C.; Hutchins, Charles R. 1981. A flora of New Mexico. Volume 2. Germany: J. Cramer. 2589 p. [37175]
213. Mast, Joy Nystrom; Fule, Peter Z.; Moore, Margaret M.; [and others]. 1999. Restoration of presettlement age structure of an Arizona ponderosa pine forest. Ecological Applications. 9(1): 228-239. [33054]
214. Mast, Joy Nystrom; Veblen, Thomas T.; Hodgson, Michael E. 1997. Tree invasion within a pine/grassland ecotone: an approach with historic aerial photography and GIS modeling. Forest Ecology and Management. 93: 181-194. [27729]
215. Mast, Joy Nystrom; Veblen, Thomas T.; Linhart, Yan B. 1998. Disturbance and climatic influences on age structure of ponderosa pine at the pine/grassland ecotone, Colorado Front Range. Journal of Biogeography. 25: 743-755. [30141]
216. Mauk, Ronald L.; Henderson, Jan A. 1984. Coniferous forest habitat types of northern Utah. Gen. Tech. Rep. INT-170. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 89 p. [1553]
217. McHugh, Charles W.; Kolb, Thomas E. 2003. Ponderosa pine mortality following fire in northern Arizona. International Journal of Wildland Fire. 12: 1-16. [43550]
218. Meinecke, E. P. 1929. Quaking aspen: A study in applied forest pathology. Tech. Bull. No. 155. Washington, DC: U.S. Department of Agriculture. 34 p. [26669]
219. Millspaugh, Joshua J.; Raedeke, Kenneth J.; Brundige, Gary C.; Willmott, Charles C. 1998. Summer bed sites of elk (Cervus elaphus) in the Black Hills, South Dakota: considerations for thermal cover management. The American Midland Naturalist. 139(1): 133-140. [27870]
220. Minor, Charles O. 1964. Site-index curves for young-growth ponderosa pine in northern Arizona. Res. Note RM-37. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 8 p. [1380]
221. Moir, W. H.; Dieterich, J. H. 1988. Old-growth ponderosa pine from succession in pine-bunchgrass forests in Arizona and New Mexico. Natural Areas Journal. 8(1): 17-24. [6781]
222. Moody, Ron; Buchanan, Les; Melcher, Ron; Wistrand, Hunter. 1992. Fire and forest Health: Southwestern Region. Albuquerque, NM: U.S. Department of Agriculture, Forest Service, Southwestern Region. 39 p. [22348]
223. Muldavin, Esteban H.; De Velice, Robert L.; Ronco, Frank, Jr. 1996. A classification of forest habitat types: Southern Arizona and portions of the Colorado Plateau. RM-GTR-287. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 130. [27968]
224. Mutch, Robert W.; Arno, Stephen F.; Brown, James K.; [and others]. 1993. Forest health in the Blue Mountains: a management strategy for fire-adapted ecosystems. Gen. Tech. Rep. PNW-GTR-310. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 14 p. [20849]
225. Naumburg, Elke; DeWald, Laura E. 1999. Relationships between Pinus ponderosa forest structure, light characteristics, and understory graminoid species presence and abundance. Forest Ecology and Management. 124: 205-215. [31128]
226. Nichol, A. A. [revisions by Phillips, W. S.]. 1952. The natural vegetation of Arizona. Tech. Bull. 68 [Revised]. Tucson, AZ: University of Arizona, Agricultural Experiment Station: 189-230. [3928]
227. Oliver, William W.; Ryker, Russell A. 1990. Pinus ponderosa Dougl. ex Laws. ponderosa pine. In: Burns, Russell M.; Honkala, Barbara H., technical coordinators. Silvics of North America. Volume 1. Conifers. Agric. Handb. 654. Washington, DC: U.S. Department of Agriculture, Forest Service: 413-424. [13399]
228. Pase, Charles P.; Granfelt, Carl Eric, tech. coords. 1977. The use of fire on Arizona rangelands. Arizona Interagency Range Committee Publication No. 4. [Place of publication unknown]: [Arizona Interagency Range Committee]. 15 p. [1827]
229. Pase, Charles P.; Pond, Floyd W. 1964. Vegetation changes following the Mingus Mountain burn. Res. Note RM-18. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 8 p. [5700]
230. Payne, Gene F. 1973. Vegetative rangeland types in Montana. Bull. 671. Bozeman, MT: Montana State University, Montana Agricultural Experiment Station. 15 p. [1847]
231. Paysen, Timothy E.; Ansley, R. James; Brown, James K.; [and others]. 2000. Fire in western shrubland, woodland, and grassland ecosystems. In: Brown, James K.; Smith, Jane Kapler, eds. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42-volume 2. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 121-159. [36978]
232. Pearson, G. A. 1934. Grass, pine seedlings and grazing. Journal of Forestry. 32: 545-555. [37426]
233. Pearson, G. A. 1951. A comparison of the climate in four ponderosa pine regions. Journal of Forestry. 49(4): 256-258. [4720]
234. Pearson, H. A.; Davis, J. R.; Schubert, G. H. 1972. Effects of wildfire on timber and forage production in Arizona. Journal of Range Management. 25: 250-253. [5664]
235. Peloquin, R. L. 1971. Variation and hybridization patterns in Pinus ponderosa and Pinus engelmannii. Santa Barbara, CA: University of California at Santa Barbara. 196 p. Dissertation. [20319]
236. Peloquin, R. L. 1984. The identification of three-species hybrids in the ponderosa pine complex. The Southwestern Naturalist. 29(1): 115-122. [20320]
237. Pfister, Robert D.; Kovalchik, Bernard L.; Arno, Stephen F.; Presby, Richard C. 1977. Forest habitat types of Montana. Gen. Tech. Rep. INT-34. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 174 p. [1878]
238. Pyne, Stephen J.; Andrews, Patricia L.; Laven, Richard D. 1984. Introduction to wildland fire. 2nd ed. New York: Wiley & Sons, Inc. 769 p. [37243]
239. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press. 632 p. [2843]
240. Read, Ralph A. 1964. Tree windbreaks for the Central Great Plains. Agric. Handb. 250. Washington, DC: U.S. Department of Agriculture, Forest Service. 68 p. [2897]
241. Roberts, David W.; Sibbernsen, John I. 1979. Forest and woodland habitat types of north central Montana. Volume 2: The Missouri River Breaks. Missoula, MT: University of Montana, School of Forestry. In cooperation with: Intermountain Forest and Range Experiment Station, Forestry Sciences Laboratory. 24 p. [2001]
242. Roberts, David W.; Sibbernsen, John I. [n.d.]. Forest habitat types of the Bear's Paw Mountains. Report prepared for the Bureau of Indian Affairs in cooperation with: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Forestry Sciences Lab. Order No. 6055-0100032, Amendment #1. Unpublished report on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Lab, Missoula, MT. 60 p. [29857]
243. Roberts, David W.; Sibbernsen, John I. [n.d.]. Forest habitat types of the Little Rocky Mountains. Report prepared for the Bureau of Indian Affairs in cooperation with: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Forestry Sciences Lab. Order No. 6055-0100430. Unpublished report on file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Lab, Missoula, MT. 60 p. [29856]
244. Romme, William H. 1982. Fire and landscape diversity in subalpine forests of Yellowstone National Park. Ecological Monographs. 52(2): 199-221. [9696]
245. Romme, William H.; Jamieson, David W.; Redders, Jeffery S.; [and others]. 1992. Old-growth forests of the San Juan National Forest in southwestern Colorado. In: Kaufmann, Merrill R.; Moir, W. H.; Bassett, Richard L., technical coordinators. Old-growth forests in the Southwest and Rocky Mountain regions: Proceedings of a workshop; 1992 March 9-13; Portal, AZ. Gen. Tech. Rep. RM-213. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 154-165. [19050]
246. Rothermel, Richard C. 1972. A mathematical model for predicting fire spread in wildland fuels. Res. Pap. INT-115. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 40 p. [16845]
247. Rumble, Mark A.; Anderson, Stanley H. 1993. Habitat selection of Merriam's turkey (Meleagris gallopavo merriami) hens with poults in the Black Hills, South Dakota. The Great Basin Naturalist. 53(2): 131-136. [21518]
248. Sackett, S. S.; Haase, S. M. 1991. Predicting forest floor weights indirectly. In: Andrews, Patricia L.; Potts, Donald F., eds. Proceedings, 11th annual conference of fire and forest meteorology; 1991 April 16-19; Missoula, MT. SAF Publication 91-04: Bethesda, MD: Society of American Foresters: 382-386. [37244]
249. Sackett, Stephen S. 1979. Natural fuel loadings in ponderosa pine and mixed conifer forests of the Southwest. Res. Pap. RM-213. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 10 p. [5665]
250. Sackett, Stephen S. 1980. Reducing natural ponderosa pine fuels using prescribed fire: two case studies. Res. Note RM-392. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 6 p. [590]
251. Sackett, Stephen S. 1984. Observations on natural regeneration in ponderosa pine following a prescribed fire in Arizona. Res. Note RM-435. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 8 p. [8093]
252. Sackett, Stephen S.; Haase, Sally M. 1996. Fuel loadings in southwestern ecosystems of the United States. In: Ffolliott, Peter F.; DeBano, Leonard F.; Baker, Malchus, B., Jr.; [and others], tech. coords. Effects of fire on Madrean Province ecosystems: a symposium proceedings; 1996 March 11-15; Tucson, AZ. Gen. Tech. Rep. RM-GTR-289. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 187-192. [28076]
253. Sackett, Stephen S.; Haase, Sally M. 1998. Two case histories for using prescribed fire to restore ponderosa pine ecosystems in northern Arizona. In: Pruden, Teresa L.; Brennan, Leonard A., eds. Fire in ecosystem management: shifting the paradigm from suppression to prescription: Proceedings, Tall Timbers fire ecology conference; 1996 May 7-10; Boise, ID. No. 20. Tallahassee, FL: Tall Timbers Research Station: 380-389. [35664]
254. Sackett, Stephen S.; Haase, Sally M.; Harrington, Michael G. 1996. Lessons learned from fire use for restoring southwestern ponderosa pine ecosystems. In: Covington, Wallace; Wagner, Pamela K., technical coordinators. Conference on adaptive ecosystem restoration and management: restoration of Cordilleran conifer landscapes of North America: Proceedings; 1996 June 6-8; Flagstaff, AZ. Gen. Tech. Rep. RM-GTR-278. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 54-61. [26924]
255. Sackett, Stephen; Haase, Sally; Harrington, M. G. 1993. Restoration of southwestern ponderosa pine ecosystems with fire. In: Covington, M. Wallace; Debano, Leonard F.; Covington, W. W., tech. coords. Sustainable ecological systems: implementing an ecological approach to land management: Proceedings; 1993 July 12-15; Flagstaff, AZ. Gen. Tech. Rep. RM-247. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 115-121. [37424]
256. Savage, Melissa. 1991. Structural dynamics of a southwestern pine forest under chronic human influence. Annals of the Association of American Geographers. 8(12): 271-289. [37427]
257. Savage, Melissa; Brown, Peter M.; Feddema, Johannes. 1996. The role of climate in a pine forest regeneration pulse in the southwestern United States. Ecoscience. 3(3): 310-318. [37430]
258. Saveland, James M.; Bunting, Stephen C. 1988. Fire effects in ponderosa pine forests. In: Baumgartner, David M.; Lotan, James E., compilers. Ponderosa pine: the species and its management: Symposium proceedings; 1987 September 29 - October 1; Spokane, WA. Pullman, WA: Washington State University, Cooperative Extension: 125-131. [9409]
259. Schmid, J. M.; Mata, S. A.; Mitchell, J. C. 1986. Number and condition of seeds in ponderosa pine cones in central Arizona. The Great Basin Naturalist. 46: 449-451. [13196]
260. Schroeder, Mark J.; Buck, Charles C. 1970. Fire weather...a guide for application of meteorological information to forest fire control operations. Agricultural Handbook 360. Washington, DC: U.S. Department of Agriculture, Forest Service. 229 p. [37423]
261. Schubert, Gilbert H. 1974. Silviculture of southwestern ponderosa pine: the status of our knowledge. Res. Pap. RM-123. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 71 p. [10888]
262. Schubert, Gilbert H.; Wellner, Charles A.; Hudson, Russell H.; Barrett, James W. 1981. Interior ponderosa pine. In: Choices in silviculture for American forests. Washington, DC: Society of American Foresters: 54-63. [6529]
263. Schultz, Brad W. 1987. Ecology of curlleaf mountain mahogany (Cercocarpus ledifolius) in western and central Nevada: population structure and dynamics. Reno, NV: University of Nevada. 111 p. Thesis. [7064]
264. Seklecki, Mariette T.; Grissino-Mayer, Henri D.; Swetnam, Thomas W. 1996. Fire history and the possible role of Apache-set fires in the Chiricahua Mountains of southeastern Arizona. In: Ffolliott, Peter F.; DeBano, Leonard F.; Baker, Malchus, B., Jr.; [and others], tech. coords. Effects of fire on Madrean Province ecosystems: a symposium proceedings; 1996 March 11-15; Tucson, AZ. Gen. Tech. Rep. RM-GTR-289. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 238-246. [28082]
265. Servis, Steve; Boucher, Paul F. 1999. Restoring fire to southwestern ecosystems: is it worth it? In: Gonzalez-Caban, Armando; Omi, Philip N., tech. coords. Proceedings of the symposium on fire economics, planning, and policy: bottom lines; 1999 April 5-9; San Diego, CA. Gen. Tech. Rep. PSW-GTR-173. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 247-253. [37051]
266. Shiflet, Thomas N., ed. 1994. Rangeland cover types of the United States. Denver, CO: Society for Range Management. 152 p. [23362]
267. Simpson, Benny J. 1988. A field guide to Texas trees. Austin, TX: Texas Monthly Press. 372 p. [11708]
268. Smith, Dwight R. 1967. Effects of cattle grazing on a ponderosa pine-bunchgrass range in Colorado. Technical Bulletin No. 1371. Washington, DC: U.S. Department of Agriculture, Forest Service. 60 p. [4763]
269. Snyder, Marc A. 1993. Interactions between Abert's squirrel and ponderosa pine: the relationship between selective herbivory and host plant fitness. The American Naturalist. 141(6): 866-879. [22172]
270. Snyder, Noel F. R.; Koenig, Susan; Johnson, Terry B. 1995. Ecological relationships of the thick-billed parrot with the pine forests of southeastern Arizona. In: DeBano, Leonard F.; Ffolliott, Peter F.; Ortega-Rubio, Alfredo; [and others], technical coordinators. Biodiversity and management of the Madrean Archipelago: the sky islands of southwestern United States and northwestern Mexico: Proceedings; 1994 September 19-23; Tucson, AZ. Gen. Tech. Rep. RM-GRT-264. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 288-293. [26243]
271. Stein, Steven J. 1988. Explanations of the imbalanced age structure and scattered distribution of ponderosa pine within a high-elevation mixed conifer forest. Forest Ecology and Management. 25: 139-153. [6236]
272. Stein, Steven J. 1988. Fire history of Panusaugunt Plateau in southern Utah. The Great Basin Naturalist. 48(1): 58-63. [4603]
273. Steinauer, Ernest M.; Bragg, Thomas B. 1987. Ponderosa pine (Pinus ponderosa) invasion of Nebraska sandhills prairie. The American Midland Naturalist. 118(2): 358-365. [6620]
274. Stephens, H. A. 1973. Woody plants of the North Central Plains. Lawrence, KS: The University Press of Kansas. 530 p. [3804]
275. Stickney, Peter F. 1989. Seral origin of species originating in northern Rocky Mountain forests. Unpublished draft on file at: U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Fire Sciences Laboratory, Missoula, MT. 10 p. [20090]
276. Stoms, David M.; Davis, Frank W.; Driese, Kenneth L.; [and others]. 1998. Gap analysis of the vegetation of the Intermountain semi-desert ecoregion. The Great Basin Naturalist. 58(3): 199-216. [30151]
277. Stuever, Mary C.; Hayden, John S. 1996. Plant associations (habitat types) of the forests and woodlands of Arizona and New Mexico. Final report: Contract R3-95-27. Placitas, NM: Seldom Seen Expeditions, Inc. 520 p. [28868]
278. Sutherland, Elaine Kennedy. 1989. The natural and unnatural fire history of Southwestern ponderosa pine. Women in Natural Resources. 11(1): 12-16. [11515]
279. Sutherland, Elaine Kennedy; Covington, W. Wallace; Andariese, Steve. 1991. A model of ponderosa pine growth response to prescribed burning. Forest Ecology and Management. 44(2-4): 161-173. [17662]
280. Swetnam, Thomas W.; Baisan, Christopher H. 1996. Fire histories of montane forests in the Madrean Borderlands. In: Ffolliott, Peter F.; DeBano, Leonard F.; Baker, Malchus, B., Jr.; [and others], tech. coords. Effects of fire on Madrean Province ecosystems: a symposium proceedings; 1996 March 11-15; Tucson, AZ. Gen. Tech. Rep. RM-GTR-289. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 15-36. [28061]
281. Swetnam, Thomas W.; Baisan, Christopher H. 1996. Historical fire regime patterns in the southwestern United States since AD 1700. In: Allen, Craig D., ed. Fire effects in Southwestern forests: Proceedings, 2nd La Mesa fire symposium; 1994 March 29-31; Los Alamos, NM. RM-GTR-286. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 11-32. [27281]
282. Swetnam, Thomas W.; Baisan, Christopher H.; Brown, Peter M.; Caprio, Anthony C. 1989. Fire history of Rhyolite Canyon, Chiricahua National Monument. Tech. Rep. No. 32. Tucson, AZ: University of Arizona, School of Renewable Natural Resources, Cooperative National Park Resources Studies Unit. 47 p. [10573]
283. Swetnam, Thomas W.; Betancourt, Julio L. 1990. Fire-southern oscillation relations in the southwestern United States. Science. 249: 1017-1020. [12106]
284. Swetnam, Thomas W.; Betancout, Julio L. 1998. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. Journal of Climate. 11: 3128-3147. [37431]
285. Swetnam, Thomas W.; Brown, Peter M. 1992. Oldest known conifers in the southwestern United States: temporal and spatial patterns of maximum age. In: Kaufmann, Merrill R.; Moir, W. H.; Bassett, Richard L., technical coordinators. Old-growth forests in the Southwest and Rocky Mountain regions: Proceedings of a workshop; 1992 March 9-13; Portal, AZ. Gen. Tech. Rep. RM-213. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 24-38. [19039]
286. Swetnam, Thomas W.; Dieterich, John H. 1985. Fire history of ponderosa pine forests in the Gila Wilderness, New Mexico. In: Lotan, James E.; Kilgore, Bruce M.; Fisher, William C.; Mutch, Robert W., technical coordinators. Proceedings, symposium and workshop on wilderness fire; 1983 November 15-18; Missoula, MT. General Technical Report INT-182. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 390-397. [7369]
287. Taylor, Dale L.; Barmore, William J., Jr. 1980. Post-fire succession of avifauna in coniferous forests of Yellowstone and Grand Teton National Parks, Wyoming. In: DeGraaf, Richard M., technical coordinator. Management of western forests and grasslands for nongame birds: Workshop proceedings; 1980 February 11-14; Salt Lake City, UT. Gen. Tech. Rep. INT-86. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 130-145. [17902]
288. Texas Natural Heritage Program. 1993. Plant communities of Texas (Series level). Austin, TX: Texas Parks and Wildlife Department. Unpublished report on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Missoula, MT. 26 p. [23810]
289. Thilenius, John F. 1972. Classification of deer habitat in the ponderosa pine forest of the Black Hills, South Dakota. Res. Pap. RM-91. Fort Collins, CO: U.S. Department of Agriculture, Forest Service. 28 p. [2317]
290. Thilenius, John F.; Brown, Gary R.; Medina, Alvin L. 1995. Vegetation on semi-arid rangelands, Cheyenne River Basin, Wyoming. Gen. Tech. Rep. RM-GTR-263. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 60 p. [26478]
291. Thill, Ronald E.; Ffolliott, Peter F.; Patton, David R. 1983. Deer and elk forage production in Arizona mixed conifer forests. Res. Pap. RM-248. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 13 p. [14381]
292. Touchan, Ramzi; Allen, Craig D.; Swetnam, Thomas W. 1996. Fire history and climatic patterns in ponderosa pine and mixed conifer forests of the Jemez Mountains, northern New Mexico. In: Allen, Craig D., ed. Fire effects in Southwestern forests: Proceedings, 2nd La Mesa fire symposium; 1994 March 29-31; Los Alamos, NM. RM-GTR-286. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 33-46. [27282]
293. Touchan, Ramzi; Swetnam, Thomas W.; Grissino-Mayer, Henri D. 1995. Effects of livestock grazing on pre-settlement fire regimes in New Mexico. In: Brown, James K.; Mutch, Robert W.; Spoon, Charles W.; Wakimoto, Ronald H., technical coordinators. Proceedings: symposium on fire in wilderness and park management; 1993 March 30 - April 1; Missoula, MT. Gen. Tech. Rep. INT-GTR-320. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 268-272. [26228]
294. Turkowski, Frank J. 1980. Carnivora food habits and habitat use in ponderosa pine forests. Res. Pap. RM-125. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 9 p. [15852]
295. U.S. Department of Agriculture, Natural Resources Conservation Service. 2015. PLANTS Database, [Online]. Available: https://plants.usda.gov /. [34262]
296. Uresk, Daniel W.; Paintner, Wayne W. 1985. Cattle diets in a ponderosa pine forest in the northern Black Hills. Journal of Range Management. 38(5): 440-442. [2401]
297. Urness, P. J.; Neff, D. J.; Watkins, R. K. 1975. Nutritive value of mule deer forages on ponderosa pine summer range in Arizona. Res. Note RM-304. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 6 p. [15854]
298. Veblen, Thomas T.; Kitzberger, Thomas; Donnegan, Joseph. 2000. Climatic and human influences on fire regimes in ponderosa pine forests in the Colorado Front Range. Ecological Applications. 10(4): 1178-1195. [35647]
299. Vincent, Dwain W. 1992. The sagebrush/grasslands of the upper Rio Puerco area, New Mexico. Rangelands. 14(5): 268-271. [19698]
300. Wade, Dale D.; Brock, Brent L.; Brose, Patrick H.; [and others]. 2000. Fire in eastern ecosystems. In: Brown, James K.; Smith, Jane Kapler, eds. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42-vol. 2. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 53-96. [36983]
301. Wagener, Willis W. 1955. Preliminary guidelines for estimating the survival of fire-damaged trees. Res. Note. No. 98. Berkeley, CA: U.S. Department of Agriculture, Forest Service, California [Pacific Southwest] Forest and Range Experiment Station. 9 p. [12345]
302. Wagener, Willis W. 1961. Guidelines for estimating the survival of fire-damaged trees in California. Misc. Paper 60. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 11 p. [4611]
303. Wasser, C. H.; Hess, Karl. 1982. The habitat types of Region II, U.S. Forest Service: a synthesis. Final Report Cooperative Agreement No. 16-845-CA. Lakewood, CO: U.S. Department of Agriculture, Forest Service, Region 2. 140 p. [5594]
304. Weaver, Harold. 1951. Fire as an ecological factor in the Southwestern ponderosa pine forests. Journal of Forestry. 49(2): 93-98. [37484]
305. Weaver, Harold. 1951. Observed effects on perennial grasses of prescribed burning tests in ponderosa pine. Arizona Cattelog. 6(6): 24-29. [4740]
306. Weaver, Harold. 1952. A preliminary report on prescribed burning in virgin ponderosa pine. Journal of Forestry. 50: 662-667. [18642]
307. Weaver, Harold. 1968. Fire and its relationship to ponderosa pine. In: Proceedings, California Tall Timbers fire ecology conference; 1967 November 9-10; Hoberg, CA. Number 7. Tallahassee, FL: Tall Timbers Research Station: 127-149. [16903]
308. Weaver, J. E. 1968. Prairie plants and their environment: A fifty-year study in the Midwest. Lincoln, NE: University of Nebraska Press. 276 p. [17548]
309. Weber, William A.; Wittmann, Ronald C. 1996. Colorado flora: eastern slope. 2nd ed. Niwot, CO: University Press of Colorado. 524 p. [27572]
310. Wells, Osborn O. 1964. Geographic variation in ponderosa pine. I. The ecotypes and their distribution. Silvae Genetica. 13(4): 89-103. [15928]
311. White, Alan S. 1985. Presettlement regeneration patterns in a Southwestern ponderosa pine stand. Ecology. 66(2): 589-594. [4621]
312. White, Carleton S. 1991. The interaction of prescribed fire, monoterpenes, and soil N-cycling processes in a stand of ponderosa pine (Pinus ponderosa). In: Nodvin, Stephen C.; Waldrop, Thomas A., eds. Fire and the environment: ecological and cultural perspectives: Proceedings of an international symposium; 1990 March 20-24; Knoxville, TN. Gen. Tech. Rep. SE-69. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: 138-144. [16644]
313. White, M. A.; Vankat, J. L. 1993. Middle and high elevation coniferous forest communities of the North Rim region of Grand Canyon National Park, Arizona, USA. Vegetatio. 109: 161-174. [25919]
314. Wiedmeier, R. D.; Pfister, J. A.; Adams, D. C.; [and others]. 1990. Effects of ponderosa pine needle consumption on ruminal microflora in cattle. Proceedings, Western Section, American Society of Animal Science. 41: 478-481. [15574]
315. Wolf, Joy J.; Mast, Joy Nystrom. 1998. Fire history of mixed-conifer forests on the North Rim, Grand Canyon National Park, Arizona. Physical Geography. 19(1): 1-14. [34709]
316. Wright, Henry A.; Bailey, Arthur W. 1982. Fire ecology: United States and southern Canada. New York: John Wiley & Sons. 501 p. [2620]
317. Wyant, James G.; Laven, Richard D.; Omi, Phillip N. 1983. Fire effects on shoot growth characteristics of ponderosa pine in Colorado. Canadian Journal of Forest Research. 13: 620-625. [4623]
318. Wyant, James G.; Omi, Philip N.; Laven, Richard D. 1986. Fire induced tree mortality in a Colorado ponderosa pine/Douglas-fir stand. Forest Science. 32(1): 49-59. [4624]
319. Wyant, James G.; Zimmerman, G. Thomas. 1984. Factors contributing to postfire tree mortality in central Rocky Mountain forests. In: New forests for a changing world: Proceedings of the 1983 Convention of The Society of American Foresters; 1983 October 16-20; Portland, OR. Bethesda, MD: Society of American Foresters: 271-275. [4625]
320. Yeager, A. F. 1935. Root systems of certain trees and shrubs grown on prairie soils. Journal of Agricultural Research. 51(12): 1085-1092. [3748]
321. Young, James A.; Evans, Raymond A. 1981. Demography and fire history of a western juniper stand. Journal of Range Management. 34(6): 501-505. [2659]
322. Youngblood, Andrew P.; Mauk, Ronald L. 1985. Coniferous forest habitat types of central and southern Utah. Gen. Tech. Rep. INT-187. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 89 p. [2684]
323. Zwolinski, Malcolm J. 1996. Effects of fire on montane forest ecosystems. In: Ffolliott, Peter F.; DeBano, Leonard F.; Baker, Malchus, B., Jr.; [and others], tech. coords. Effects of fire on Madrean Province ecosystems: a symposium proceedings; 1996 March 11-15; Tucson, AZ. Gen. Tech. Rep. RM-GTR-289. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 55-63. [28062]
324. Callaham, Robert Z. 2013. Pinus ponderosa: A taxonomic review with five subspecies in the United States. Res. Pap. PSW-RP-264. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station. 52 p. [89267]