Index of Species Information
SPECIES: Abies lasiocarpa
|
|
 |
| Subalpine fir in Glacier National Park. Public domain image by Ansel Adams from the series "Ansel Adams Photographs of National Parks and Monuments", compiled from 1941-1942. |
Introductory
SPECIES: Abies lasiocarpa
AUTHORSHIP AND CITATION:
Uchytil, Ronald J. 1991. Abies lasiocarpa. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station,
Fire Sciences Laboratory (Producer). Available:
https://www.fs.usda.gov/database/feis/plants/tree/abilas/all.html [].
Revisions:
Images were added on 22 March 2018.
ABBREVIATION:
ABILAS
ABILASA
ABILASL
SYNONYMS:
Abies balsamea ssp. lasiocarpa (Hook.) Boivin
Abies balsamea var. fallax (Engelm.) Boivin
NRCS PLANT CODE:
ABLA
ABLAA
ABLAL
COMMON NAMES:
subalpine fir
alpine fir
western balsam fir
balsam
white balsam
balsam fir
white fir
Rocky Mountain fir
pino real blanco
cork-bark fir
corkbark fir
Arizona fir
TAXONOMY:
The genus Abies consists of about 40 species of evergreen trees found in
the northern hemisphere. Nine species of Abies, including subalpine
fir, are native to the United States [75]. The currently accepted
scientific name of subalpine fir is Abies lasiocarpa (Hook.) Nutt.
[75]. Subalpine fir is widely distributed and exhibits geographic
variation. Two varieties are recognized based on morphological
differences [75]:
Abies lasiocarpa var. arizonica, corkbark fir
Abies lasiocarpa var. lasiocarpa, typical variety of subalpine fir
Subalpine fir hybridizes with balsam fir (A. balsamea) where their
ranges overlap in the Canadian Rockies [41].
LIFE FORM:
Tree
FEDERAL LEGAL STATUS:
No special status
OTHER STATUS:
NO-ENTRY
DISTRIBUTION AND OCCURRENCE
SPECIES: Abies lasiocarpa
GENERAL DISTRIBUTION:
Subalpine fir is the mostly widely distributed fir in North America,
spanning more than 32 degrees of latitude [11]. It occurs chiefly in
mountainous areas from the Yukon interior near treeline and along the
coast of southeastern Alaska south through western Alberta and British
Columbia to southern Colorado and scattered mountain ranges of Arizona
and New Mexico [54,75]. In the western portion of its range, subalpine
fir does not occur along the western slope of the Coast Range in
southern British Columbia or along the Coast Ranges of Washington and
Oregon but does occur on Vancouver Island and in the Olympic Mountains
of Washington [11]. It occurs on both slopes of the Cascade Mountains
as far south as southern Oregon [11]. The two varieties are distributed
as follows [11,75]:
A. l. var. lasiocarpa (typical variety) - almost the same as the species,
but not in central and southeastern Arizona.
A. l. var. arizonica - from central Colorado to southwestern New Mexico, and
in southeastern and central Arizona.
Subalpine fir and corkbark fir occur together in scattered mountain
ranges in southwestern Colorado, northern, western, and southwestern New
Mexico, and in the high mountains of Arizona [11].
 |
|
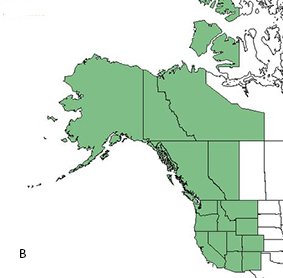 |
| Distributions of corkbark fir (A) and the typical variety (B). Maps courtesy of USDA, NRCS. 2018. The PLANTS Database.
National Plant Data Team, Greensboro, NC [117] [2018, March 22]. |
ECOSYSTEMS:
FRES20 Douglas-fir
FRES22 Western white pine
FRES23 Fir - spruce
FRES24 Hemlock - Sitka spruce
FRES25 Larch
FRES26 Lodgepole pine
FRES44 Alpine
STATES:
AK AZ CO ID MT NV NM OR UT WA
WY AB BC YT
BLM PHYSIOGRAPHIC REGIONS:
1 Northern Pacific Border
2 Cascade Mountains
5 Columbia Plateau
6 Upper Basin and Range
8 Northern Rocky Mountains
9 Middle Rocky Mountains
11 Southern Rocky Mountains
12 Colorado Plateau
KUCHLER PLANT ASSOCIATIONS:
K004 Fir - hemlock forest
K012 Douglas-fir forest
K015 Western spruce - pine forest
K018 Pine - Douglas-fir forest
K020 Spruce - fir - Douglas-fir forest
K021 Southwestern spruce - fir forest
K052 Alpine meadows and barren
SAF COVER TYPES:
201 White spruce
202 White spruce - paper birch
205 Mountain hemlock
206 Engelmann spruce - subalpine fir
208 Whitebark pine
209 Bristlecone pine
210 Interior Douglas-fir
212 Western larch
213 Grand fir
215 Western white pine
216 Blue spruce
217 Aspen
218 Lodgepole pine
219 Limber pine
223 Sitka spruce
224 Western hemlock
226 Coastal true fir - hemlock
253 Black spruce - white spruce
SRM (RANGELAND) COVER TYPES:
NO-ENTRY
HABITAT TYPES AND PLANT COMMUNITIES:
Forests in which subalpine fir attains climax dominance or codominance
are widespread throughout the mountains of western North America. The
subalpine fir series generally occupies cold, high elevation mountain
forests. Engelmann spruce (Picea engelmannii) is usually associated
with subalpine fir. It occurs as either a climax codominant or as a
persistent, long-lived seral species in most subalpine fir habitat
types.
Published classification schemes listing subalpine fir as a dominant
part of the vegetation in habitat types (hts), community types (cts),
plant associations (pas), ecosystem associations (eas), site types (sts)
or dominance types (dts) are presented below:
Area Classification Authority
AK: ----- general veg. cts Viereck & Dyrness 1980
AZ: San Francisco forest, alpine &
Peaks RNA meadow cts Rominger & Paulik 1983
AZ, NM: ----- forest & woodland hts Layser & Schubert 1979
Apache, Gila,
Cibola NFs forest hts Fitzhugh & others 1987
s of Mogollon
Rim forest hts Develice & Ludwig 1983b
n AZ, n NM forest hts Larson & Moir 1987
CO: Arapaho &
Roosevelt NFs forest hts Hess & Alexander 1986
Gunnison &
Uncompahgre NFs forest hts Komarkova & others 1988
Routt NF forest hts Hoffman & Alexander 1980
White River- grassland, shrubland,
Arapaho NF & forestland hts Hess & Wasser 1982
White River NF forest hts Hoffman & Alexander 1983
w CO riparian pas Baker 1989a
ID: Sawtooth, White
Cloud, Boulder,
& Pioneer Mtns general veg. cts Schlatterer 1972
c ID forest hts Steele & others 1981
n ID forest hts Cooper & others 1987
se ID aspen cts Mueggler & Campbell 1986
e ID, w WY forest hts Steele & others 1983
MT: ----- forest hts Pfister & others 1977
----- riparian dts Hansen & others 1988
c, e MT riparian cts, hts Hansen & others 1990
nw MT riparian hts, cts Boggs & others 1990
sw MT riparian rst, cts, hts Hansen & others 1989
NM: Cibola NF forest hts Alexander & others 1987
Lincoln NF forest hts Alexander & others 1984
n NM, s CO forest hts Develice & Ludwig 1983a
n NM, s CO forest hts Develice & others 1986
OR: Wallowa-Whitman NF steppe & forest pas Johnson & Simon 1987
Eagle Cap
Wilderness general veg. cts Cole 1982
OR, WA: ----- general veg. cts Franklin & Dyrness 1973
Blue Mtns general veg. pas Hall 1973
UT: ----- aspen cts Mueggler & Campbell 1986
c, s UT forest hts Youngblood & Mauk 1985
n UT forest hts Mauk & Henderson 1984
WA: Okanogan NF forest pas Williams & Lillybridge 1983
Mount Rainier NP forest pas Franklin & others 1988
North Cascades NP forest pas Agee & Kertis 1987
e WA, n ID forest hts, cts Daubenmire & Daubenmire 1968
WY: Bridger-Teton NF aspen cts Youngblood & Mueggler 1981
Medicine NF forest hts Alexander & others 1986
Bighorn Mtns forest hts Hoffman & Alexander 1976
Wind River Mtns forest hts Reed 1976
USFS R-2 general veg. pas Johnston 1987
USFS R-2 general veg. hts,pas Wasser & Hess 1982
USFS R-4 aspen cts Mueggler 1988
w-c AB forest cts Corns 1983
BC: ----- grassland, forest hts McLean 1970
----- general veg. eas Pojar & others 1984
nw BC forest eas Haeussler & others 1985
MANAGEMENT CONSIDERATIONS
SPECIES: Abies lasiocarpa
WOOD PRODUCTS VALUE:
Subalpine fir wood is odorless, light-weight, soft, and low in bending
and compressive strength [11]. It is easy to work, glues well, and
holds nails and screws fairly well. The wood is primarily used for
products such as lumber for home construction and for prefabricated wood
products [9]. Subalpine fir has excellent pulping properties [9]. Use
for poles and pilings requires large amounts of preservatives because
the wood decays rapidly [108].
IMPORTANCE TO LIVESTOCK AND WILDLIFE:
Subalpine-fir-dominated stands generally do not produce enough forage
for livestock but do provide browse and cover for large and small
wildlife species. Mule deer, elk, moose, woodland caribou, black bear,
and grizzly bear often use subalpine fir habitats as summer range
[10,23,113]]. Subalpine fir forests are generally not suitable winter
range for deer and elk because of heavy snowpack, but some lower
elevation subalpine fir habitat types are used by moose and woodland
caribou during the winter [23,61,89]. Subalpine fir forests support
numerous species of small mammals and birds. The snowshoe hare, flying
squirrel, red squirrel, porcupine, pine marten, fisher, lynx, and
several species of mice, voles, chipmunks, and shrews all inhabit
subalpine fir forests [26,104,113]. Numerous species of birds nest and
feed in subalpine fir forests, including several woodpeckers,
flycatchers, kinglets, nuthatches, juncos, thrushes, chickadees,
crossbills, the pine siskin, owls, and grouse [104,113].
The young growth of subalpine fir is sometimes eaten by mule deer, elk,
bighorn sheep, and snowshoe hares, but it is not an important food item.
Subalpine fir comprises only a small portion of the summer diet of
mountain goats but can be a major food source in the winter and spring
[99]. Throughout much of Montana, Idaho, and Wyoming, subalpine fir is
an important winter food of moose [89]. On moose winter range near
Jackson Hole, Wyoming, an average of 13 to 18 percent of small subalpine
fir trees were browsed by moose, and 44 to 78 percent of the branches on
trees browsed were utilized [61]. In Yellowstone National Park, grizzly
bears sometimes strip the bark of subalpine fir to feed on the
underlying cambium [19]. The winter diet of blue grouse consists
primarily of conifer needles. These grouse often winter in subalpine
stands and may feed heavily on the needles and buds of subalpine fir
[103,113].
Subalpine fir seeds are eaten by several species of small mammals and
birds. Red squirrels eat seeds from cached subalpine fir cones [71].
Fir seeds are also eaten by chipmunks and mice. Several birds,
including chickadees, nuthatches, crossbills, the pine siskin, and the
Clark's nutcracker remove and eat the seeds from fir cones [49,77].
Because subalpine fir seeds are large, comprising about 26 percent of a
cone's weight, they are an energy-efficient food source for small birds
[49]. Small birds may make considerable use of fir seeds, but their
foraging is scattered and sporadic throughout subalpine forests [49].
PALATABILITY:
The palatability of subalpine fir to domestic livestock is low [30].
Its palatability to big game animals is generally low also, but in some
locations it is highly palatable to moose and mountain goats during
winter and spring [89,99]. The seeds are palatable to numerous small
mammal species [77]. Red squirrels generally eat subalpine fir seeds
after other cached conifer seeds have been consumed [71]. The needles
are highly palatable to blue grouse.
The relish and degree of use shown by livestock and wildlife species for
subalpine fir in several western states is rated as follows
[30,61,89,96]:
CO MT WY UT ID
Cattle poor poor poor poor ----
Sheep poor poor poor poor ----
Horses poor poor poor poor ----
Pronghorn ---- ---- poor poor ----
Elk ---- poor poor poor ----
Moose ---- good good ---- fair
Mule deer ---- poor poor poor ----
White-tailed deer ---- ---- poor ---- ----
Small mammals ---- fair fair good ----
Small nongame birds ---- ---- fair good ----
Upland game birds ---- fair fair good ----
Waterfowl ---- ---- poor poor ----
NUTRITIONAL VALUE:
Subalpine fir is low in protein value but fair in energy value [30].
Percent composition of subalpine fir browse collected near Jackson Hole,
Wyoming, was as follows [61]:
date crude ether crude nitrogen
collected protein extract fiber free extract
11/25 5.57 7.53 20.19 50.26
A study in Montana found the following concentration of elements in
subalpine fir needles and twigs [110]:
1-yr-old green needles twigs < 0.25 inch (0.64 cm) in diameter
(micrograms/gram [mean]) (micrograms/gram [mean])
Ca 9722 5840
Cu 7.4 7.9
Fe 64 182
K 5553 7031
Mg 819 1038
Mn 1020 587
N 10690 4962
Na 103 124
P 1450 2254
Zn 43 5
(percent [mean]) (percent [mean])
Ash 3.5 3.5
COVER VALUE:
Big game: Subalpine fir habitat types provide excellent hiding cover
for deer, elk, mountain goats, moose, and bear [10,113]. Certain low
elevation subalpine fir forests may be used by elk during calving, and
high elevation subalpine fir forests by bighorn sheep during lambing and
lamb rearing [113]. Dense stands provide cool summertime shade for big
game animals [71]. In Yellowstone National Park, grizzly bear daybeds
are often found in subalpine fir stands [19].
Small mammals and birds: Small subalpine firs provide good year-round
hiding cover. Dense thickets of small trees are often nearly
impenetrable and provide hiding places for small mammals such as
snowshoe hares and porcupines [13,71]. Blue grouse often overwinter in
subalpine trees and rely almost exclusively on them for escape cover
[103]. Subalpine fir snags are used by numerous cavity-nesting birds,
but are generally less preferred than those of associated conifers
[105].
The degree to which subalpine fir provides environmental protection
during one or more seasons for wildlife species is as follows [30]:
CO MT WY UT
Pronghorn ---- ---- poor poor
Elk good fair good good
Mule deer good fair good good
White-tailed deer ---- ---- poor ----
Small mammals good good good good
Small nongame birds good fair good good
Upland game birds good good good fair
Waterfowl ---- ---- poor poor
VALUE FOR REHABILITATION OF DISTURBED SITES:
Subalpine fir can be planted on disturbed sites within forest vegetation
types where it naturally occurs [92]. It is generally recommended for
cool and moist sites within subalpine areas [120]. Its erosion control
potential is listed as medium in Utah and Montana, and high in Colorado
[30]. Because this wide-ranging tree exhibits a large degree of genetic
variation, seed or nursery stock for rehabilitation projects should come
from a local source. Transplanting nursery stock is generally more
successful than direct seeding [92]. Seedlings exhibit very slow
initial growth and are therefore usually outplanted as 2- to 3-year-old
seedlings [41]. Wild seedlings may also be transplanted [120]. A
maximum spacing of 10 x 10 feet (3 x 3 m) has been recommended for
seedlings or transplants [120]. Methods for collecting, processing,
testing, storing, and planting subalpine fir seeds have been discussed
in detail [32,41].
OTHER USES AND VALUES:
Subalpine fir is sometimes used as a landscape plant to produce
screenings or windbreaks [114]. In the Pacific Northwest it is
sometimes transplanted into rock gardens or simulated subalpine settings
[69].
Native Americans used various parts of subalpine fir for numerous
purposes. A hair tonic was prepared by mixing powdered needles with
deer grease. Finely ground needles were also sprinkled on open cuts.
Sticky resin collected from the bark was boiled and used as an
antiseptic for wounds or as a tea for colds. Boughs were placed in
rooms for their aroma, and pulverized needles were used as a body scent
or as perfume for clothing [53].
Resin from the bark is used in the optical industry and in laboratories
as a cement for lenses and microscope slides [71].
OTHER MANAGEMENT CONSIDERATIONS:
Timber harvest: Shelterwood and individual tree selection silvicultural
methods favor subalpine fir over Engelmann spruce, lodgepole pine (Pinus
contorta), and Douglas-fir (Pseudotsuga menziesii); clearcutting and
group selection cutting favor subalpine fir over Pacific silver fir
(Abies amabilis), grand fir (A. grandis), and mountain hemlock (Tsuga
mertensiana) where they grow together [11]. The seed tree method is
generally not used because of the susceptibility of subalpine fir to
windthrow [11]. In the Rocky Mountains, clearcutting and shelterwood
cutting have been the most commonly used harvesting methods in
old-growth Engelmann spruce-subalpine fir stands because these stands
tend to be even-aged and overmature [8]. Uneven-aged silviculture can
pose a problem because residual subalpine fir trees damaged during
thinning operations are susceptible to attack by decay fungi.
Silvicultural systems and cutting methods for managing subalpine fir
have been discussed in detail [7,8,9,11].
Pests and diseases: Subalpine fir is attacked by numerous insects. The
most destructive seem to be the western spruce budworm, western balsam
bark beetle, and balsam woolly aphid [11]. Subalpine fir is one of the
most common hosts of the western spruce budworm. This pest generally
attacks low and middle elevation subalpine fir forests but is largely
absent from high elevation forests [21]. The balsam woolly aphid has
virtually eliminated subalpine fir from some stands in the Cascades
[11]. Other insect pests include the Douglas-fir tussock moth, western
black-headed budworm, and fir engraver beetle.
Subalpine fir is susceptible to annosus root disease, caused by the
fungus Heterobasidion annosum, which results in root and butt decay.
Outbreaks of this disease are often centered around large 20-year-old or
older fir stumps that contain the fungus' fruiting bodies [102].
Subalpine fir is most seriously affected by this disease in the northern
and central Rocky Mountains [123], and is affected to a lesser extent in
the Pacific Northwest [102]. Subalpine fir is susceptible to several
other wood rotting fungi that cause heart, trunk, butt, or root rots,
including brown stringy rot, red heart rot, red ring rot, shoestring
rot, brown cubical rot, white spongy root rot, and white pocket rot
[39]. Trees weakened by wood rots often become infested by fir engraver
beetles and usually succumb to windfall and breakage [11]. Fir broom
rust is another common problem in Engelmann spruce-subalpine fir stands
and causes bole deformation, spike tops and wind breakage, and makes
trees more susceptible to decay fungi [11].
Habitat for threatened and endangered species: Old-growth subalpine fir
stands in northern Idaho may provide critical habitat for woodland
caribou [23]. Numerous subalpine fir habitat types, especially those
containing huckleberries (Vaccinium spp.), provide critical habitat for
grizzly bears [127].
BOTANICAL AND ECOLOGICAL CHARACTERISTICS
SPECIES: Abies lasiocarpa
GENERAL BOTANICAL CHARACTERISTICS:
Subalpine fir is a native, coniferous, evergreen tree. It is the
smallest of the eight species of fir native to the western United
States. Five growth forms, each apparently an adaptation to a
particular environment, are described below [9,54]:
1. The typical form is found throughout much of the subalpine zone.
These trees have an extremely narrow and dense crown with short
branches. Trees growing in openings retain their lower branches, which
often droop and extend down to the ground. Trees growing in the
overstory may be clear of lower branches for 20 to 30 percent of the
tree's height.
2. A somewhat broad-crowned, bullet-shaped tree is more typical of
older specimens and drier climates.
3. A mature tree with a layered apron is occasionally found in some
areas.
4. A flag form tree often occurs at timberline. These individuals are
characterized by an upright trunk that extends above a krummholzlike
mat. Branches on the trunk generally grow only along the leeward side
of the trunk, giving the plant a flaglike appearance.
5. The krummholz form is typical of alpine areas above timberline. In
these areas, because of cold temperatures and severe winds, subalpine
fir grows in dwarfed, shrubby mats along the ground, and is often much
broader than it is tall.
The typical form often grows to heights of 60 to 100 feet (18-30 m), and
trunk diameters reach 18 to 24 inches (46-61 cm) [39]. Trees up to 130
feet (40 m) tall and 30 inches (76 cm) in diameter have been found but
are rare [39]. Subalpine fir grows very slowly; 150- to 200-year-old
trees are usually only 10 to 20 inches (25-50 cm) in diameter [39].
Trees seldom live more than 250 years because they are very susceptible
to heart rots [9].
Needles are blunt tipped, flattened, and 1 to 1.2 inches (2.5-3 cm) long
[25,57]. Bark on young trees is thin, gray, and smooth, with numerous
resin vesicles; on older trees it is shallowly fissured and scaly [9].
Corkbark fir is generally recognized by its creamy-white, thick, corky
bark [25]. The root system generally is shallow but under favorable
conditions may develop relatively deep laterals [9].
Subalpine fir is monoecious. Single or small bunches of erect female
cones occur in the upper part of the crown on the upper side of young
branches. Dense clusters of the smaller male cones occur lower on the
crown on the underside of 1-year-old twigs. Subalpine fir seeds are
0.23 to 0.28 inch (6-7 mm) long and have broad wings about 0.4 inch (1
cm) long [25,121]. Corkbark fir seeds are about 70 percent larger than
subalpine fir seeds [39].
RAUNKIAER LIFE FORM:
Phanerophyte
REGENERATION PROCESSES:
Cone and seed production: Subalpine fir can begin producing cones when
20 years old and 4 or 5 feet (1.2-1.5 m) tall, but under closed forest
conditions seed production is generally not significant until trees are
older and taller [11]. Corkbark fir generally does not produce cones
until about 50 years old [39]. Nearly all cones are produced on the
uppermost part of the crown. Maximum seed production is by dominant
trees between 150 and 200 years old [39]. Yearly seed production is
very erratic; good seed crops are produced every 3 to 5 years, with
light crops or crop failures in between [39,40,54,86]. Corkbark fir is
also a poor seed producer, having more crop failure years than good seed
crop years [9]. Subalpine fir averages 34,800 seeds per pound
(76,700/kg), while corkbark fir seeds average 22,300 per pound
(49,150/kg) [41].
Seed predation: Insect pests reduce seed yields by feeding on cones and
seeds; however, the magnitude of loss is variable [11]. Red squirrels
cut and cache large quantities of subalpine fir cones [9]. After
dispersal, numerous small rodents and birds consume seeds from the
ground.
Dispersal: Mature subalpine and corkbark fir seeds have a large wing
and are dispersed primarily by wind in the fall as cones disintegrate on
the tree. Seeds travel primarily in the direction of prevailing winds,
but upslope drafts can influence dispersal at low and middle elevations
[9]. Studies in Colorado showed that about one-half of subalpine fir
seeds dispersed into clearcuts fell within 100 feet (30 m) of the
clearcut's windward edge, while the remainder fell within 260 feet (80
m) of the edge [86]. Some seeds are also dispersed by red squirrels
which cut and cache cones before they disintegrate; seeds commonly
germinate from these middens, forming thickets [71].
Germination and viability: Seeds overwinter under or in snow. This
cold, moist stratification is required for germination [41].
Germination begins in the spring a few days after snowmelt and is
usually completed within a few weeks [9,106]. Percent germination is
low due to unsound seed; about 31 to 38 percent for subalpine fir and 26
to 33 percent for corkbark fir [39,41]. Under natural conditions seeds
remain viable for 1 year [41]. Stratification procedures for stored
seeds have been described in detail [41,74].
Seedling establishment and survival: Seedlings establish best on
mineral soil seedbeds but will also establish on other surfaces
including litter, duff, and decaying wood [11]. Because Engelmann
spruce requires a mineral soil seedbed, subalpine fir seedlings usually
outnumber spruce seedlings in the understory of spruce-fir stands.
Thus, even though it is short-lived, many ecologists consider subalpine
fir better able to regenerate under climax conditions than Engelmann
spruce. Subalpine fir is very shade tolerant and easily establishes
under a closed canopy. Throughout the Rocky Mountains subalpine fir is
reproducing abundantly under conditions of dense shade and is often
abundant as seedlings and saplings in spruce-fir forests, even where
Engelmann spruce dominates the overstory [11,67].
At higher elevations, seedling survival is sometimes greater on duff
seedbeds because the duff helps protect seedlings from high-intensity
summer rain storms and frost heaving [35]. At lower elevations,
seedling densities are often greater on mineral soils [35]. In a
spruce-fir forest in southeastern Wyoming, Knapp and Smith [67] found
that 42 percent of subalpine fir seedlings were on litter deeper than 1
inch (2.5 cm), compared with only 5 percent of Engelmann spruce
seedlings. Subalpine fir is able to establish in duff because of its
rapid root growth. Comparing seedlings grown in a greenhouse, subalpine
fir's taproot length (29 mm) 2 weeks after germination was over 200
percent greater than the taproot length of Engelmann spruce (9.4 mm)
[67].
Growth: Subalpine fir seedlings grow very slowly. One-year-old
seedlings are frequently less than 1 inch (2.5 cm) tall [11]. One study
found 15-year-old seedlings averaged only 11 inches (28 cm) in height on
burned-over slopes, 10 inches (25 cm) on cut-over dry slopes, and 6
inches (15 cm) on cut-over wet flats [11]. Under favorable conditions
trees reach a height of 4 to 5 feet (1.2-1.5 m) in 20 to 40 years [11].
Under a closed canopy, trees 4 to 6 feet tall (1.2-1.8 m) are often 35
to 50 years old.
Vegetative reproduction: Near timberline subalpine fir frequently
reproduces by layering, probably as a result of heavy snow, wind, and
cold temperatures which restrict growth away from the ground. Layering
often results in clusters of subalpine fir growing near timberline [13].
Under closed forest canopy, reproduction by layering is negligible.
SITE CHARACTERISTICS:
Subalpine fir is a middle to upper elevation mountain conifer. It
generally occupies sites with a short growing season caused by cold
winters, cool summers, frequent summer frosts, and heavy snowpack. It
forms extensive forests between warm and dry lower elevation forests of
Douglas-fir, white fir (Abies concolor), grand fir, lodgepole pine, or
blue spruce (Picea pungens) and higher elevation alpine tundra
[23,29,112,125]. At its lower elevational limits, subalpine fir is
often restricted to streambottoms, ravines, frosty basins, or north
exposures. It increasingly occupies westerly and easterly aspects with
increasing elevation and may occupy all aspects at upper timberline
[23,91].
Stand condition and associated conifers: Throughout its range,
subalpine fir is most commonly associated with Engelmann spruce. These
two species frequently occur as codominants forming widespread subalpine
forests. In the central and southern Rocky Mountains, Engelmann spruce
commonly makes up 70 percent of overstory trees, with subalpine fir
dominating the understory [9]. Within spruce-fir forests of this
region, Engelmann spruce tends to be more important at higher elevations
and on wetter sites, while subalpine fir is more abundant on drier lower
elevation sites [9]. In the northern Rocky Mountains, subalpine fir
typically dominates climax stands, but Engelmann spruce becomes
increasingly important on moist, cool sites [23,91]. Other associates
which vary by latitude and elevation are listed below [39]:
Location Elevation Associates
northern Rocky Mtns low western white pine (Pinus monticola),
Douglas-fir, western larch (Larix
occidentalis), grand fir, western hemlock
(Tsuga heterophylla), western redcedar
(Thuja plicata)
high lodgepole pine, subalpine larch (Larix
lyallii), whitebark pine (Pinus
albicaulis), mountain hemlock
central Rocky Mtns low lodgepole pine, Douglas-fir, aspen
(Populus tremuloides), blue spruce
high whitebark pine, limber pine (Pinus
flexilis), bristlecone pine (P. aristata)
southern Rocky Mtns low white fir, Douglas-fir, blue spruce, aspen
high corkbark fir
Cascade Mtns low Pacific silver fir, mountain hemlock,
lodgepole pine
high mountain hemlock, whitebark pine
Understory associates: Understory vegetation is extremely variable,
changing with elevation, exposure, and soil moisture. Habitat type and
plant association guides describe characteristic understory plants for
differing sites.
Elevation: Alexander and others [11] described the following
elevational ranges for subalpine fir:
Coast Range of southeastern Alaska - subalpine fir is found from sea
level to 3,500 feet (0-1,067 m).
Coast Range and interior plateaus of Yukon Territory and British
Columbia - subalpine fir is found from 2,000 to 5,000 feet (610-1,524
m).
Olympic and Cascade Mountains of Washington and Oregon - subalpine fir
is generally found from 4,000 to 6,000 feet (1,219-1,829 m), but may be
found as low as 2,000 feet (610 m) along cold streambottoms and on lava
flows, and as high as 8,000 feet (2,438 m) on sheltered slopes.
Rocky Mountains of British Columbia and Alberta south of the Peace River
- subalpine fir is found from 3,000 to 7,000 feet (914-2,134 m) but is
more abundant above 5,000 feet (1,524 m).
Rocky Mountains of Montana and Idaho and associated ranges of eastern
Oregon and Washington - subalpine fir grows from 2,000 to 11,000 feet
(610-3,353 m) but is most common at 5,000 to 9,000 feet (1,524-2,743 m).
Rocky Mountains of Wyoming, Utah, and Colorado - subalpine fir occurs
from 8,000 to 11,500 feet (2,438-3,506 m) but is most common at 9,000 to
11,000 feet (2,743-3,353 m).
Rocky Mountains of New Mexico and Arizona - subalpine fir occurs from
8,000 to 12,000 feet (2,438-3,658 m) but is usually found on north
slopes from 9,500 to 11,000 feet (2,896-3,353 m).
SUCCESSIONAL STATUS:
In the Rocky Mountains, subalpine fir is a shade-tolerant climax species
favored by long fire-free intervals. Its seedlings outcompete spruces,
lodgepole pine, and Douglas-fir when light intensities are less than 50
percent of full sunlight, but cannot compete with these conifers under
brighter light [11]. In Montana and Idaho and in the mountains of
eastern Washington and eastern Oregon, subalpine fir often forms pure
stands at climax, but it may also mix with Engelmann spruce, which,
although considered to be seral to subalpine fir, outlives it and
persists to climax. In the Rocky Mountains north and south of Montana
and Idaho, Engelmann spruce and subalpine fir may codominate at climax
[11].
Throughout much of the Cascade Mountains subalpine fir grows as a
shade-intolerant, seral species and is gradually replaced by more
shade-tolerant associates such as Pacific silver fir, grand fir, and
mountain hemlock [43]. It often invades recently disturbed areas with
lodgepole pine. It also pioneers harsh sites on raw geologically young
surfaces such as lava flows and talus slopes and on climatically harsh
sites near timberline [43].
In areas where subalpine fir is a climax dominant, succession following
disturbance varies depending upon the severity and type of disturbance,
elevation, and the availability of conifer seeds. Subalpine fir may
establish immediately following disturbances if mature trees survive to
provide seeds and seral species such as lodgepole pine and aspen are
scarce. Near treeline, it may take 100 years or more for subalpine fir
to establish seedlings following fire because an increase in herbaceous
species prevents seeds from reaching mineral soil and the harsh climate
kills many seedlings that do establish [18,109]. Aspen and lodgepole
pine are the most common seral species. They often form pure stands and
completely dominate low and middle elevation stands within the subalpine
fir zone following large fires [5,68]. These species grow rapidly and
quickly overtop any subalpine fir seedlings that may establish at the
same time. Aspen stands can sometimes persist for decades or even
centuries when conifer seed trees are eliminated [29]. When lodgepole
pine establishes immediately following stand-destroying fires, it often
forms even-aged dense stands that dominate for 100 to 300 years.
Because it is very shade tolerant, subalpine fir eventually establishes
under the pine canopy, usually within 100 years, and attains dominance
as the pine stand begins to break up [90,98].
In many of the warmer and lower elevation subalpine fir habitat types,
subalpine fir has not achieved climax dominance because of repeated
fires which favor shade-intolerant seral conifers. Many of these
habitat types are in midsuccessional stages. Lodgepole pine, western
larch, western white pine, or Douglas-fir dominate the overstory, but
subalpine fir seedlings and saplings occur in the understory [93,113].
SEASONAL DEVELOPMENT:
Subalpine fir requires 2 years to complete its reproductive cycle [115].
Cones are initiated in the spring of the first year as microscopic
primordia within vegetative buds. Bud differentiation occurs in
midsummer, and separate seed-cone and pollen-cone buds develop until
each becomes dormant in the fall [115]. During the spring of the second
year, cone buds resume growth and conelets are recognizable in the early
spring. During the second year, reproduction phenology generally
proceeds as follows:
Phenological event Location Timing of event Reference
flowering sw MT, nw WY mid-June - early July
cones full size " late August
seeds dispersed " early Sept - early Oct [41,101]
flowering nw MT, n ID mid-June - early July
cones full size " late July - early Aug
seeds dispersed " mid-Sept [41,101]
flowering OR late May - early July
seed dispersal " early Oct [41]
male bud burst Linn, OR early to mid-May
female bud burst " mid to late May
pollen shed " June
seed dispersal begins " early October [45]
flowering AZ, San Fran.Peaks late June
cone ripening " mid-Sept - early Oct
seed dispersal " late Sept - early Oct [41]
FIRE ECOLOGY
SPECIES: Abies lasiocarpa
FIRE ECOLOGY OR ADAPTATIONS:
Plant adaptations to fire: Subalpine fir is very fire sensitive and
generally suffers high mortality even from low intensity fires. It
relies on wind-dispersed seeds which readily germinate on fire-prepared
seedbeds to colonize burned areas. The occasional mature tree which
survives fire, those escaping fire in small, unburned pockets, and trees
adjacent to burned areas provide seeds to colonize burned sites. In
subalpine habitats, scattered subalpine fir trees often escape fire
because of discontinuous fuels, broken and rocky terrain, and the moist
and cool environment [78,87,91].
Fire regime: Subalpine fir habitat types vary from cold and wet at
higher elevations to warm and moist or cool and dry at lower elevations.
This environmental gradient influences the mean fire return interval
(MFRI). Relatively dry lower elevation subalpine fir habitat types have
more frequent and less intense fires than moist middle and upper
elevation subalpine fir habitat types [12,91]. Such forests in the
Bitterroot National Forest in Montana have a MFRI of 17 to 28 years
[14]. Fires at this frequency kill subalpine fir and keep these forests
dominated by seral conifers such as lodgepole pine, Douglas-fir, or
western larch. Moist, middle and upper elevation subalpine fir habitat
types, however, generally experience high intensity stand-replacing
fires at intervals of 100 years or more. Mean fire return intervals for
middle and upper elevation subalpine fir habitat types in several areas
are presented below:
Location Community dominants MFRI Reference
(years)
Kananaskis Park, AB subalpine fir, spruce, 90 [12]
lodgepole pine
northern Cascades, WA subalpine fir 154 [2]
northern Cascades, WA subalpine fir-lodgepole pine 109 [2]
Olympic NP, WA subalpine fir 150 [116]
Yellowstone NP, WY subalpine fir 300-350 [98]
Coram Exp. Forest, nw MT western larch, Douglas-fir, 117-146 [129]
lodgepole pine, subalpine fir
Fuels and fire behavior: The fuel structure in subalpine fir-dominated
stands promotes highly destructive stand-destroying fires. Fuel loads
in subalpine fir stands are greater than in lower elevation montane
stands because the cool and moist environment slows the decomposition of
organic matter allowing fuels to accumulate more rapidly [1]. Fuel beds
tend to be irregular, with over twice as much fuel accumulating under
the narrow-crowned trees as between them [116]. The needles are small
and fine and form a compact fuel bed in which fire spreads slowly [34].
These concentrated, slow burning fuels frequently produce flames high
enough to reach subalpine fir's low-growing dead branches [116]. Thus
crowning is common in subalpine fir stands.
 |
| Downed woody fuels resulting from a 2017 avalanche on the Gallatin National Forest. Image by Garon Smith, used with permission. |
Once a crown fire begins, it spreads easily because subalpine fir has a
tendency to grow in dense stands and has highly flammable foliage. A
lightning strike on May 7, 1987, in a subalpine fir-mountain hemlock
stand in Mount Rainier National Park started a crown fire even though
the ground was still partially snow covered. The fire spread slowly
through the tree crowns by (1) igniting lichens draped along the fine
branches, (2) preheating and igniting the foliage, and (3) spreading to
a nearby tree by igniting its lichens [62].
FIRE REGIMES:
Find fire regime information for the plant communities in which this
species may occur by entering "subalpine fir" in the FEIS home page
under "Find Fire Regimes".
POSTFIRE REGENERATION STRATEGY:
crown-stored residual colonizer; short-viability seed in on-site cones
secondary colonizer; off-site seed carried to site after year 2
off-site colonizer; seed carried by wind; postfire years 1 and 2
FIRE EFFECTS
SPECIES: Abies lasiocarpa
IMMEDIATE FIRE EFFECT ON PLANT:
Subalpine fir is one of the least fire-resistant western conifers. It
is very susceptible to fire because it has (1) thin bark that provides
little insulation for the cambium, (2) bark which ignites readily, (3)
shallow roots which are susceptible to soil heating, (4) low-growing
branches, (5) a tendency to grow in dense stands, (6) highly flammable
foliage, and (7) moderate to heavy lichen growth [37,111].
Subalpine fir forests are normally subject to highly destructive crown
fires that occur at 100-year or longer intervals. Such fires typically
kill all subalpine fir trees. Subalpine fir is also very susceptible to
surface fires because fine fuels which are often concentrated under
mature trees burn slowly and girdle the thin-barked bole [34].
DISCUSSION AND QUALIFICATION OF FIRE EFFECT:
NO-ENTRY
PLANT RESPONSE TO FIRE:
Following fire, subalpine fir reestablishes via seeds dispersed by wind
from trees surviving in protected pockets or from trees adjacent to
burned areas. Subalpine fir readily establishes on burned mineral soil
seedbeds [107]. Ash does not affect germination, but if it is deep, it
can prevent a seedling's roots from reaching mineral soil [85].
Although seedling establishment is often favored by shade, it will
establish in full sunlight following fire [87].
The rate of establishment is quite variable, and depends on the
proximity of the seed source (because the heavy seeds are dispersed over
short distances) and seed production during the year of the fire and
immediate postfire years. In general, subalpine fir seedling
establishment is very slow in areas suffering large, continuous crown
fires but is relatively rapid on small burned-over areas where a seed
source is nearby [90,124,128]. Three years after a late August wildfire
in northern Colorado, in a dense, mature stand composed of Engelmann
spruce, subalpine fir, and lodgepole pine, subalpine fir had established
15,200 seedlings per acre (37,500/ha) on small burns that were less than
one-tenth of an acre in size. But on areas within the middle of the
main burn, subalpine fir had established only 12 seedling per acre
(30/ha) 3 years after the fire [16]. In Colorado, Peet [90] found a
75-year-old burn that had few conifer seedlings even though an
old-growth subalpine fir-Engelmann spruce stand was 218 yards (200 m)
away.
Reinvasion into large burns is slow because much of the seed source is
destroyed. However, sometimes sporadic survivors provide a limited seed
source so that a small number of seedlings establish quickly following
fire. When this occurs, large quantities of seeds are dispersed several
decades later as the early invading seedlings mature and reach cone
bearing age [128].
On areas where subalpine fir is abundant and lodgepole pine scarce
before burning, subalpine fir establishes quickly following fire if
sufficient numbers of seed trees survive or are near the burn. However,
if lodgepole pine is present prior to burning, it usually seeds in
aggressively and assumes a dominant role because it quickly overtops any
fir seeding in at the same time [34]. Subalpine fir can be suppressed
for several decades in seral lodgepole stands which develop following
fire; one-hundred-year-old individuals may be only 3 feet (0.9 m) tall
[90]. It may take 50 to 150 years after a fire for substantial
subalpine fir establishment under dense lodgepole pine stands
[18,109,128].
In the Olympic Mountains, tree seedling establishment following fires in
closed mountain hemlock-subalpine fir forests was higher during wet
growing seasons than during dry growing seasons. Establishment rates
were higher near the edge of a fire or near survivors than in areas
removed from a seed source [4]. On many burned areas, subalpine fir did
not establish seedlings for several years because of poor seed crops.
On some burns there was a lag time of 40 to 50 years after fire before
there was substantial seedling establishment. This was a result of
early invading trees maturing and dispersing seeds [4].
High elevation subalpine fir stands that have burned often remain open
for several decades or more [18,31]. The harsh environment near
treeline makes it difficult for tree seedlings to establish and survive
[18]. Grasses and sedges may form a mat in subalpine meadows which
prevents tree seeds from reaching mineral soil [109].
DISCUSSION AND QUALIFICATION OF PLANT RESPONSE:
For further information on subalpine fir response to fire, see
Fire Case Studies. Hamilton's Research Project Summary and Research Papers
(Hamilton 2006a, Hamilton 2006b)provide information on prescribed fire and
postfire response of plant community species, including subalpine fir,
that was not available when this species review was originally written.
FIRE MANAGEMENT CONSIDERATIONS:
Subalpine fir is very fire sensitive and is often killed even by surface
fires. Following timber harvest, on sites where subalpine fir is not a
preferred species, light surface fires may be used to kill subalpine
fir and promote the establishment of other conifers [93].
Fuels remain moist in many high elevation subalpine fir habitat types
during most of the year, leaving only a short time period during certain
years when prescribed burning can take place [63,91].
Subalpine fir seeds germinate poorly in soils under burned slash piles
[130] but readily germinate on mineral soil seedbeds prepared by
broadcast burning [16,107].
SPECIES: Abies lasiocarpa
FIRE CASE STUDY CITATION:
Uchytil, Ronald J., compiler. 1991. Effects of a prescribed crown fire on
Table Mountain, Washington, on subalpine fir. In: Abies lasiocarpa.
In: Fire Effects Information System, [Online]. U.S. Department of
Agriculture, Forest Service, Rocky Mountain Research Station, Fire
Sciences Laboratory (Producer). Available:
https://www.fs.usda.gov
/database/feis/plants/tree/abilas/all.html#FireCaseStudies
[].
REFERENCE:
Woodard, Paul Michael. 1977. Effects of prescribed burning on two
different-aged high-elevation plant communities in eastern Washington.
Seattle, WA: University of Washington. 228 p. Dissertation. [124].
SEASON/SEVERITY CLASSIFICATION:
fall/severe
STUDY LOCATION:
The study took place in the Cascade Mountains in central Washington
approximately midway between the towns of Ellensburg and Wenatchee.
PREFIRE VEGETATIVE COMMUNITY:
The study area is within the subalpine fir (Abies lasiocarpa) zone. Two
adjacent but different-aged stands were burned. One area was a young,
lodgepole pine (Pinus contorta) thicket, the other was a old, decadent
stand of lodgepole pine with subalpine fir and Engelmann spruce (Picea
engelmannii) attaining dominance. Each is described below.
Lodgepole pine thicket - Lodgepole pine is the overstory dominant, but
subalpine fir and Engelmann spruce are also principal components.
Subalpine fir and Engelmann spruce are codominants in the intermediate
size classes. Subalpine fir is the only tree regenerating under the
canopy. Understory plants include elk sedge (Carex geyeri), Hood sedge
(C. hoodii), heartleaf arnica (Arnica cordifolia), broadleaf arnica (A.
latifolia), bigleaf lupine (Lupinus polyphyllus), and dwarf bilberry
(Vaccinium myrtillus).
Snag area - This is a decadent lodgepole pine stand, with subalpine fir
and Engelmann spruce dominating the overstory canopy. Fir and spruce
codominate the intermediate size classes, but subalpine fir is the only
tree regenerating under the canopy. Predominant understory plants
include subalpine fir, elk sedge, broadleaf arnica, and mosses
(Rhacomitrium canascens, Polytridum commune).
TARGET SPECIES PHENOLOGICAL STATE:
NO-ENTRY
SITE DESCRIPTION:
The burned site is a southwest facing gentle slope at an elevation of
5,596 to 5,776 feet (1,706-1,761 m). A total of 27 acres (10.9 ha) were
burned.
Climate: The climate is typical of most areas within the subalpine
zone. The winters are cold and wet and the summers cool and dry. Frost
and freezing temperatures can occur during any month of the year. Over
70 percent of precipitation falls as snow between October and March.
Soil and duff: The two stands have similar soils, but the soil in the
snag area is more fertile and better developed. In both areas soils
are derived from basalt residium, have a clay-loam texture, and average
6 inches (15.2 cm) deep. The effective rooting depth was about 20
inches (51 cm) in the thicket area, and 20 to 40 inches (51-102 cm) in
the snag area. On both areas, duff was generally from 1 to 4 inches
(0.4-1.6 cm) thick. The mean depth of duff was 2.3 inches (5.94 cm) on
the thicket area, and 1.9 inches (4.92 cm) on the snag area.
Fuel loading: Prior to burning, mean fuel loads were as follows:
fuels thicket area snag area
tons/acre tonnes/ha tons/acre tonnes/ha
dead and down wood
0.0-0.25 inch (0.0-0.6 cm) 0.6 1.3 0.9 2.0
0.26-0.99 inch (0.61-2.5 cm) 2.1 4.6 2.9 6.4
1.0-3.0 inches (2.6-7.6 cm) 4.4 9.8 5.1 11.4
> 3.0 inches (7.6 cm) rotten 17.4 38.9 30.9 69.2
> 3.0 inches (7.6 cm) solid 17.0 38.1 50.1 112.2
litter 41.4 92.7 89.8 201.2
duff 32.5 72.8 30.0 67.1
FIRE DESCRIPTION:
The fire was ignited with drip torches on September 30, 1975, taking
approximately 1 hour and 20 minutes to ignite the entire 27 acres (10.9
ha). Actual fire prescriptions at the time of ignition were as follows:
ambient air temperature = 60 to 63 degrees F (16-17 C)
relative humidity = 19-21 percent
wind = calm gusts to 15.6 miles/hr (26 km/hr), from the south-southwest
days since last rain = 15
fine fuel moisture content = 13 percent
Within about 10 minutes after ignition, trees began to crown out.
Fire behavior for each area is summarized below:
Snag area - Crowning occurred throughout most of the snag area. Flame
heights were estimated to be 125 feet (38 m) by one observer, and 50
feet (15 m) above the tops of 90 foot (27 m) crowns by two other
observers. The fire consumed all small downed and dead wood from 0.0 to
1 inch (0-2.5 cm) in diameter, as well as needles and small twigs on
live standing trees. Ninety-six percent of down and dead fuels less
than 3 inches (7.6 cm) in diameter were consumed. In general, 90 to 100
percent of the duff layer was removed. Many trees less than 3 inches
(7.6 cm) in diameter at the base were completely consumed, and nearly
all standing snags were blown down or burned down. Where crowning
occurred, the only thing that remained immediately following the fire
was reddened soil, ash-covered soil, large-diameter logs, and dead
trees.
Lodgepole thicket - Fire within the lodgepole thicket was much less
severe. The crown fire which occurred within the snag area stopped when
it met the boundary of the lodgepole thicket. Dead and down fuels less
than 3 inches (7.6 cm) in diameter were reduced by 70 percent. Dead and
down fuels greater than 3 inches (7.6 cm) in diameter were reduced 34
percent. Duff was reduced about 25 percent.
FIRE EFFECTS ON TARGET SPECIES:
Where crowning occurred subalpine fir suffered 100 percent mortality.
Fire burning within the crown of live trees consumed all needles and
small twigs leaving scorched skeletons. Many live trees less than 3
inches (7.6 cm) in diameter at the base were completely consumed.
One year after this fire, subalpine fir seedlings established from seed
produced by unburned trees adjacent to the burned area. However,
stocking was considered low and attributed to a poor seed crop.
Seedling establishment was restricted to areas along the margin of the
burn.
Subalpine fir pre and postfire frequency and cover are summarized below:
thicket area snag area
prefire postfire prefire postfire
frequency 100% 38% 100% 12%
relative ground cover 37% 12% 20% 4%
FIRE MANAGEMENT IMPLICATIONS:
High intensity crown fires which are common in subalpine fir forests,
kill all or nearly all seed trees within a burned area. If subalpine
fir is to naturally regenerate on this type of burn, seeds must come
from adjacent unburned stands. Because subalpine fir seeds are
dispersed over relatively short distances, initial seedling
establishment is restricted to the burn's edge. Subalpine fir cone
production can be erratic from year to year, with the best regeneration
occurring during good seed crop years.
REFERENCES
SPECIES: Abies lasiocarpa
REFERENCES:
1. A. D. Revill Associates. 1978. Ecological eff. of fire and its mgmt. in
Canada's national parks: a synthesis of the literature. Vols 1&2. Lit.
Rev. & Annot. Bibliography. Ottawa, ON: Parks Canada, National Parks
Branch, Natural Resources Division. 345 p. [3416]
2. Agee, James K.; Finney, Mark; DeGouvenain, Roland. 1990. Forest fire
history of Desolation Peak, Washington. Canadian Journal of Forest
Research. 20: 350-356. [11035]
3. Agee, James K.; Kertis, Jane. 1987. Forest types of the North Cascades
National Park Service Complex. Canadian Journal of Botany. 65:
1520-1530. [6327]
4. Agee, James K.; Smith, Larry. 1984. Subalpine tree reestablishment after
fire in the Olympic Mountains, Washington. Ecology. 65(3): 810-819.
[6102]
5. Alexander, Billy G., Jr.; Fitzhugh, E. Lee; Ronco, Frank, Jr.; Ludwig,
John A. 1987. A classification of forest habitat types of the northern
portion of the Cibola National Forest, New Mexico. Gen. Tech. Rep.
RM-143. Fort Collins, CO: U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station. 35 p.
[4207]
6. Alexander, Billy G., Jr.; Ronco, Frank, Jr.; Fitzhugh, E. Lee; Ludwig,
John A. 1984. A classification of forest habitat types of the Lincoln
National Forest, New Mexico. Gen. Tech. Rep. RM-104. Fort Collins, CO:
U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest
and Range Experiment Station. 29 p. [300]
7. Alexander, Robert R. 1974. Silviculture of subalpine forests in the
central and southern Rocky Mountains: the status of our knowledge. Res.
Pap. RM-121. Fort Collins, CO: U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station. 88 p.
[8223]
8. Alexander, Robert R. 1986. Silvicultural systems and cutting methods for
old-growth spruce-fir forests in the central and southern Rocky
Mountains. Gen. Tech. Rep. RM-126. Fort Collins, CO: U.S. Department of
Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment
Station. 33 p. [8221]
9. Alexander, Robert R. 1987. Ecology, silviculture, and management of the
Engelmann spruce-subalpine fir type in the central and southern Rocky
Mountains. Agric. Handb. 659. Washington, DC: U.S. Department of
Agriculture, Forest Service. 144 p. [8399]
10. Alexander, Robert R.; Hoffman, George R.; Wirsing, John M. 1986. Forest
vegetation of the Medicine Bow National Forest in southeastern Wyoming:
a habitat type classification. Res. Pap. RM-271. Fort Collins, CO: U.S.
Department of Agriculture, Forest Service, Rocky Mountain Forest and
Range Experiment Station. 39 p. [307]
11. Alexander, Robert R.; Shearer, Raymond C.; Shepperd, Wayne D. 1984.
Silvical characteristics of subalpine fir. Gen. Tech. Rep. RM-115. Fort
Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky
Mountain Forest and Range Experiment Station. 29 p. [7479]
12. Arno, Stephen F. 1980. Forest fire history in the northern Rockies.
Journal of Forestry. 78(8): 460-465. [11990]
13. Arno, Stephen F.; Hammerly, Ramona P. 1977. Northwest trees. Seattle,
WA: The Mountaineers. 222 p. [4208]
14. Arno, Stephen F.; Petersen, Terry D. 1983. Variation in estimates of
fire intervals: a closer look at fire history on the Bitterroot National
Forest. Res. Pap. INT-301. Ogden, UT: U.S. Department of Agriculture,
Forest Service, Intermountain Forest and Range Experiment Station. 8 p.
[10336]
15. Baker, William L. 1989. Classification of the riparian vegetation of the
montane and subalpine zones in western Colorado. Great Basin Naturalist.
49(2): 214-228. [7985]
16. Barth, Richard C. 1970. Revegetation after a subalpine wildfire. Fort
Collins, CO: Colorado State University. 142 p. Thesis. [12458]
17. Bernard, Stephen R.; Brown, Kenneth F. 1977. Distribution of mammals,
reptiles, and amphibians by BLM physiographic regions and A.W. Kuchler's
associations for the eleven western states. Tech. Note 301. Denver, CO:
U.S. Department of the Interior, Bureau of Land Management. 169 p.
[434]
18. Billings, W. D. 1969. Vegetational pattern near alpine timberline as
affected by fire-snowdrift interactions. Vegetatio. 19: 192-207.
[12824]
19. Blanchard, Bonnie M. 1980. Grizzly bear - habitat relationships in the
Yellowstone area. Int. Conf. Bear Research and Management. 5: 118-123.
[8386]
20. Boggs, Keith; Hansen, Paul; Pfister, Robert; Joy, John. 1990.
Classification and management of riparian and wetland sites in
northwestern Montana. Missoula, MT: University of Montana, School of
Forestry, Montana Forest and Conservation Experiment Station, Montana
Riparian Association. 217 p. Draft Version 1. [8447]
21. Carlson, Clinton E.; Fellin, David G.; Schmidt, Wyman C. 1983. The
western spruce budworm in northern Rocky Mountain forests: a review of
ecology, past insecticidal treatments and silvicultural practices. In:
O'Loughlin, Jennifer; Pfister, Robert D., eds. Management of
second-growth forests: The state of knowledge and research needs:
Proceedings of a symposium; 1982 May 14; Missoula, MT. Missoula, MT:
University of Montana, School of Forestry, Montana Forest and
Conservation Experiment Station: 76-103. [7097]
22. Cole, David N. 1982. Vegetation of two drainages in Eagle Cap
Wilderness, Wallowa Mountains, Oregon. Res. Pap. INT-288. Ogden, UT:
U.S. Department of Agriculture, Forest Service, Intermountain Forest and
Range Experiment Station. 42 p. [658]
23. Cooper, Stephen V.; Neiman, Kenneth E.; Steele, Robert; Roberts, David
W. 1987. Forest habitat types of northern Idaho: a second approximation.
Gen. Tech. Rep. INT-236. Ogden, UT: U.S. Department of Agriculture,
Forest Service, Intermountain Research Station. 135 p. [867]
24. Corns, I. G. W. 1983. Forest community types of west-central Alberta in
relation to selected environmental factors. Canadian Journal of Forest
Research. 13: 995-1010. [691]
25. Cronquist, Arthur; Holmgren, Arthur H.; Holmgren, Noel H.; Reveal, James
L. 1972. Intermountain flora: Vascular plants of the Intermountain West,
U.S.A. Vol. 1. New York: Hafner Publishing Company, Inc. 270 p. [717]
26. Daubenmire, Rexford F.; Daubenmire, Jean B. 1968. Forest vegetation of
eastern Washington and northern Idaho. Technical Bulletin 60. Pullman,
WA: Washington State University, Agricultural Experiment Station. 104 p.
[749]
27. DeVelice, Robert L.; Ludwig, John A. 1983. Climax forest series of
northern New Mexico and southern Colorado. In: Moir, W. H.; Hendzel,
Leonard, tech. coords. Proceedings of the workshop on Southwestern
habitat types; 1983 April 6-8; Albuquerque, NM. Albuquerque, NM: U.S.
Department of Agriculture, Forest Service, Southwestern Region: 45-53.
[779]
28. DeVelice, Robert L.; Ludwig, John A. 1983. Forest habitat types south of
the Mogollon Rim, Arizona and New Mexico. Final Report. Cooperative
Agreement No. 28-K2-240 between U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station and New
Mexico State University. Las Cruces, NM: New Mexico State University. 47
p. [780]
29. DeVelice, Robert L.; Ludwig, John A.; Moir, William H.; Ronco, Frank,
Jr. 1986. A classification of forest habitat types of northern New
Mexico and southern Colorado. Gen. Tech. Rep. RM-131. Fort Collins, CO:
U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest
and Range Experiment Station. 59 p. [781]
30. Dittberner, Phillip L.; Olson, Michael R. 1983. The plant information
network (PIN) data base: Colorado, Montana, North Dakota, Utah, and
Wyoming. FWS/OBS-83/86. Washington, DC: U.S. Department of the Interior,
Fish and Wildlife Service. 786 p. [806]
31. Douglas, George W.; Ballard, T. M. 1971. Effects of fire on alpine plant
communities in the North Cascades, Washington. Ecology. 52(6):
1058-1064. [6738]
32. Edwards, D. G. W. 1982. Collection, processing, testing, and storage of
true fir seeds--a review. In: Oliver, Chadwick Dearing; Kenady, Reid M.,
eds. Proceedings of the biology and management of true fir in the
Pacific Northwest symposium; 1981 February 24-26; Seattle-Tacoma, WA.
Contribution No. 45. Seattle, WA: University of Washington, College of
Forest Resources; Portland, OR: U.S. Department of Agriculture, Forest
Service, Pacific Northwest Forest and Range Experiment Station: 113-137.
[11894]
33. Eyre, F. H., ed. 1980. Forest cover types of the United States and
Canada. Washington, DC: Society of American Foresters. 148 p. [905]
34. Fahnestock, George R. 1976. Fires, fuels, and flora as factors in
wilderness management: the Pasayten Case. In: Proceedings, Tall Timbers
Fire Ecology Conference; 1974 October 16-17; Portland, OR. Number 15.
Tallahassee, FL: Tall Timbers Research Station: 33-69. [6453]
35. Fiedler, Carl E.; McCaughey, Ward W.; Schmidt, Wyman C. 1985. Natural
regeneration in Intermountain spruce-fir forests--a gradual process.
Res. Pap. INT-343. Ogden, UT: U.S. Department of Agriculture, Forest
Service, Intermountain Forest and Range Experiment Station. 12 p.
[7482]
36. Fitzhugh, E. Lee; Moir, William H.; Ludwig, John A.; Ronco, Frank, Jr.
1987. Forest habitat types in the Apache, Gila, and part of the Cibola
National Forests, Arizona and New Mexico. Gen. Tech. Rep. RM-145. Fort
Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky
Mountain Forest and Range Experiment Station. 116 p. [4206]
37. Flint, Howard R. 1925. Fire resistance of northern Rocky Mountain
conifers. Idaho Forester. 7: 7-10, 41-43. [4700]
38. Fonda, R. W.; Bliss, L. C. 1969. Forest vegetation of the montane and
subalpine zones, Olympic Mountains, Washington. Ecological Monographs.
39(3): 271-301. [12909]
39. Fowells, H. A., compiler. 1965. Silvics of forest trees of the United
States. Agric. Handb. 271. Washington, DC: U.S. Department of
Agriculture, Forest Service. 762 p. [12442]
40. Franklin, J. F. 1968. Cone production by upper slope conifers. Research
Paper PNW-60. Portland, OR: U.S. Department of Agriculture, Forest
Service, Pacific Northwest Forest and Range Experiment Station. 21 p.
[12912]
41. Franklin, Jerry F. 1974. Abies Mill. fir. In: Schopmeyer, C. S.,
technical coordinator. Seeds of woody plants in the United States.
Agric. Handb. 450. Washington, DC: U.S. Department of Agriculture,
Forest Service: 168-183. [7566]
42. Franklin, Jerry F.; Dyrness, C. T. 1973. Natural vegetation of Oregon
and Washington. Gen. Tech. Rep. PNW-8. Portland, OR: U.S. Department of
Agriculture, Forest Service, Pacific Northwest Forest and Range
Experiment Station. 417 p. [961]
43. Franklin, Jerry F.; Mitchell, Russel G. 1967. Successional status of
subalpine fir in the Cascade Range. Research Paper PNW-46. Portland, OR:
U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest
and Range Experiment Station. 15 p. [963]
44. Franklin, Jerry F.; Moir, William H.; Hemstrom, Miles A.; [and others].
1988. The forest communities of Mount Rainier National Park. Scientific
Monograph Series No 19. Washington, DC: U.S. Department of the Interior,
National Park Service. 194 p. [12392]
45. Franklin, Jerry F.; Ritchie, Gary A. 1970. Phenology of cone and shoot
development of noble fir and some associated true firs. Forest Science.
16: 356-364. [12911]
46. Garrison, George A.; Bjugstad, Ardell J.; Duncan, Don A.; [and others].
1977. Vegetation and environmental features of forest and range
ecosystems. Agric. Handb. 475. Washington, DC: U.S. Department of
Agriculture, Forest Service. 68 p. [998]
47. Haeussler, S.; Pojar, J.; Geisler, B. M.; [and others]. 1985. A guide to
the interior cedar-hemlock zone, northwestern transitional subzone
(ICHg), in the Prince Rupert Forest Region, British Columbia. Land
Management Report Number 26; ISSN 0702-9861. Victoria, BC: British
Columbia, Ministry of Forests. 263 p. [6930]
48. Hall, Frederick C. 1973. Plant communities of the Blue Mountains in
eastern Oregon and southeastern Washington. R6-Area Guide 3-1. Portland,
OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest
Region. 82 p. [1059]
49. Halvorson, Curtis H. 1986. Influence of vertebrates on conifer seed
production. In: Shearer, Raymond C., compiler. Proceedings--conifer tree
seed in the Inland Mountain West symposium; 1985 August 5-6; Missoula,
MT. Gen. Tech. Rep. INT-203. Ogden, UT: U.S. Department of Agriculture,
Forest Service, Intermountain Research Station: 201-222. [13115]
50. Hansen, Paul; Boggs, Keith; Pfister, Robert; Joy, John. 1990.
Classification and management of riparian and wetland sites in central
and eastern Montana. Missoula, MT: University of Montana, School of
Forestry, Montana Forest and Conservation Experiment Station, Montana
Riparian Association. 279 p. [12477]
51. Hansen, Paul L.; Chadde, Steve W.; Pfister, Robert D. 1988. Riparian
dominance types of Montana. Misc. Publ. No. 49. Missoula, MT: University
of Montana, School of Forestry, Montana Forest and Conservation
Experiment Station. 411 p. [5660]
52. Hansen, Paul; Pfister, Robert; Joy, John; [and others]. 1989.
Classification and management of riparian sites in southwestern Montana.
Missoula, MT: University of Montana, School of Forestry, Montana
Riparian Association. 292 p. Draft Version 2. [8900]
53. Hart, J. 1976. Montana--native plants and early peoples. Helena, MT:
Montana Historical Society. 75 p. [9979]
54. Henderson, Jan A. 1982. Ecology of subalpine fir. In: Oliver, Chadwick
Dearing; Kenady, Reid M., eds. Proceedings of the biology and management
of true fir in the Pacific Northwest symposium; 1981 February 24-26;
Seattle-Tacoma, WA. Contribution No. 45. Seattle, WA: University of
Washington, College of Forest Resources: 53-58. [6759]
55. Hess, Karl; Alexander, Robert R. 1986. Forest vegetation of the Arapaho
and Roosevelt National Forests in central Colorado: a habitat type
classification. Res. Pap. RM-266. Fort Collins, CO: U.S. Department of
Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment
Station. 48 p. [1141]
56. Hess, Karl; Wasser, Clinton H. 1982. Grassland, shrubland, and
forestland habitat types of the White River-Arapaho National Forest.
Final Report. Fort Collins, CO: U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station. 335 p.
[1142]
57. Hitchcock, C. Leo; Cronquist, Arthur; Ownbey, Marion. 1969. Vascular
plants of the Pacific Northwest. Part 1: Vascular cryptograms,
gymnosperms, and monocotyledons. Seattle, WA: University of Washington
Press. 914 p. [1169]
58. Hoffman, George R.; Alexander, Robert R. 1976. Forest vegetation of the
Bighorn Mountains, Wyoming: a habitat type classification. Res. Pap.
RM-170. Fort Collins, CO: U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station. 38 p.
[1180]
59. Hoffman, George R.; Alexander, Robert R. 1980. Forest vegetation of the
Routt National Forest in northwestern Colorado: a habitat
classification. Res. Pap. RM-221. Fort Collins, CO: U.S. Department of
Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment
Station. 41 p. [1179]
60. Hoffman, George R.; Alexander, Robert R. 1983. Forest vegetation of the
White River National Forest in western Colorado: a habitat type
classification. Res. Pap. RM-249. Fort Collins, CO: U.S. Department of
Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment
Station. 36 p. [1178]
61. Houston, Douglas B. 1968. The Shiras Moose in Jackson Hole, Wyoming.
Tech. Bull. No. 1. [Place of publication unknown]: The Grand Teton
Natural History Association. 110 p. [7824]
62. Huff, Mark. 1988. Mount Rainier: fire and ice. Park Science. 8(3):
22-23. [6567]
63. Johnson, Charles G., Jr.; Simon, Steven A. 1987. Plant associations of
the Wallowa-Snake Province: Wallowa-Whitman National Forest.
R6-ECOL-TP-255A-86. Baker, OR: U.S. Department of Agriculture, Forest
Service, Pacific Northwest Region, Wallowa-Whitman National Forest. 399
p. [9600]
64. Johnston, Barry C. 1987. Plant associations of Region Two: Potential
plant communities of Wyoming, South Dakota, Nebraska, Colorado, and
Kansas. 4th ed. R2-ECOL-87-2. Lakewood, CO: U.S. Department of
Agriculture, Forest Service, Rocky Mountain Region. 429 p. [3519]
65. Kartesz, John T.; Kartesz, Rosemarie. 1980. A synonymized checklist of
the vascular flora of the United States, Canada, and Greenland. Volume
II: The biota of North America. Chapel Hill, NC: The University of North
Carolina Press; in confederation with Anne H. Lindsey and C. Richie
Bell, North Carolina Botanical Garden. 500 p. [6954]
66. Kearney, Thomas H.; Peebles, Robert H.; Howell, John Thomas; McClintock,
Elizabeth. 1960. Arizona flora. 2d ed. Berkeley, CA: University of
California Press. 1085 p. [6563]
67. Knapp, Alan K.; Smith, William K. 1982. Factors influencing understory
seedling establishment of Engelmann spruce and subalpine fir in
southeast Wyoming. Canadian Journal of Botany. 60(753): 2753-2761.
[12913]
68. Komarkova, Vera; Alexander, Robert R.; Johnston, Barry C. 1988. Forest
vegetation of the Gunnison and parts of the Uncompahgre National
Forests: a preliminary habitat type classification. Gen. Tech. Rep.
RM-163. Fort Collins, CO: U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station. 65 p.
[5798]
69. Kruckeberg, A. R. 1982. Gardening with native plants of the Pacific
Northwest. Seattle: University of Washington Press. 252 p. [9980]
70. Kuchler, A. W. 1964. Manual to accompany the map of potential vegetation
of the conterminous United States. Special Publication No. 36. New York:
American Geographical Society. 77 p. [1384]
71. Lanner, Ronald M. 1983. Trees of the Great Basin: A natural history.
Reno, NV: University of Nevada Press. 215 p. [1401]
72. Larson, Milo; Moir, W. H. 1987. Forest and woodland habitat types (plant
associations) of northern New Mexico and northern Arizona. 2d ed.
Albuquerque, NM: U.S. Department of Agriculture, Forest Service,
Southwestern Region. 90 p. [8947]
73. Layser, Earle F.; Schubert, Gilbert H. 1979. Preliminary classification
for the coniferous forest and woodland series of Arizona and New Mexico.
Res. Pap. RM-208. Fort Collins, CO: U.S. Department of Agriculture,
Forest Service, Rocky Mountain Forest and Range Experiment Station. 27
p. [1428]
74. Leadem, C. L. 1989. Stratification and quality assessment of Abies
lasiocarpa seeds. Victoria, BC: B. C. Ministry of Forests, Research
Branch. 18 p. [12004]
75. Little, Elbert L., Jr. 1979. Checklist of United States trees (native
and naturalized). Agric. Handb. 541. Washington, DC: U.S. Department of
Agriculture, Forest Service. 375 p. [2952]
76. Lyon, L. Jack; Stickney, Peter F. 1976. Early vegetal succession
following large northern Rocky Mountain wildfires. In: Proceedings, Tall
Timbers fire ecology conference and Intermountain Fire Research Council
fire and land management symposium; 1974 October 8-10; Missoula, MT. No.
14. Tallahassee, FL: Tall Timbers Research Station: 355-373. [1496]
77. Martin, Alexander C.; Zim, Herbert S.; Nelson, Arnold L. 1951. American
wildlife and plants. New York: McGraw-Hill Book Company, Inc. 500 p.
[4021]
78. Mauk, Ronald L.; Henderson, Jan A. 1984. Coniferous forest habitat types
of northern Utah. Gen. Tech. Rep. INT-170. Ogden, UT: U.S. Department of
Agriculture, Forest Service, Intermountain Forest and Range Experiment
Station. 89 p. [1553]
79. McLean, Alastair. 1970. Plant communities of the Similkameen Valley,
British Columbia. Ecological Monographs. 40(4): 403-424. [1620]
80. Moir, William H.; Ludwig, John A. 1979. A classification of spruce-fir
and mixed conifer habitat types of Arizona and New Mexico. Res. Pap.
RM-207. Fort Collins, CO: U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station. 47 p.
[1677]
81. Moss, E. H. 1955. The vegetation of Alberta. Botanical Review. 21(9):
493-567. [6878]
82. Mueggler, Walter F. 1988. Aspen community types of the Intermountain
Region. Gen. Tech. Rep. INT-250. Ogden, UT: U.S. Department of
Agriculture, Forest Service, Intermountain Research Station. 135 p.
[5902]
83. Mueggler, Walter F.; Campbell, Robert B., Jr. 1982. Aspen community
types on the Caribou and Targhee National Forests in southeastern Idaho.
Res. Pap. INT-294. Ogden, UT: U.S. Department of Agriculture, Forest
Service, Intermountain Forest and Range Experiment Station. 32 p.
[1713]
84. Mueggler, Walter F.; Campbell, Robert B., Jr. 1986. Aspen community
types of Utah. Res. Pap. INT-362. Ogden, UT: U.S. Department of
Agriculture, Forest Service, Intermountain Research Station. 69 p.
[1714]
85. Muri, Glen. 1955. The effect of simulated slash burning on germination,
primary survival and top-root ratios of Engelmann spruce and alpine fir.
Res. Note. 14. Vancouver, BC: University of British Colubmia, Forest
Club. 7 p. [4027]
86. Noble, Daniel L.; Ronco, Frank, Jr. 1978. Seedfall and establishment of
Engelmann spruce and subalpine fir in clearcut openings in Colorado.
RM-200. Fort Collins, CO: U.S. Department of Agriculture, Forest
Service, Rocky Mountain Forest and Range Experiment Station. 12 p.
[7481]
87. Parminter, John. 1983. Fire-ecological relationships for the
biogeoclimatic zones of the Cassiar Timber Supply Area: summary report.
In: Northern Fire Ecology Project, Cassiar Timber Supply Area. Victoria,
BC: Province of British Columbia, Ministry of Forests. 64 p. [9201]
88. Parminter, John. 1984. Fire-ecological relationships for the
biogeoclimatic zones of the northern portion of the Mackenzie Timber
Supply Area: summary report. In: Northern Fire Ecology Project: Northern
Mackenzie Timber Supply Area. Victoria, BC: Province of British
Columbia, Ministry of Forests. 59 p. [9205]
89. Peek, James M. 1974. Intial response of moose to a forest fire in
northeastern Minnesota. American Midland Naturalist. 91(2): 435-438.
[16531]
90. Peet, Robert K. 1981. Forest vegetation of the Colorado Front Range:
composition and dynamics. Vegetatio. 45: 3-75; 1981. [1867]
91. Pfister, Robert D.; Kovalchik, Bernard L.; Arno, Stephen F.; Presby,
Richard C. 1977. Forest habitat types of Montana. Gen. Tech. Rep.
INT-34. Ogden, UT: U.S. Department of Agriculture, Forest Service,
Intermountain Forest and Range Experiment Station. 174 p. [1878]
92. Plummer, A. Perry. 1977. Revegetation of disturbed Intermountain area
sites. In: Thames, J. C., ed. Reclamation and use of disturbed lands of
the Southwest. Tucson, AZ: University of Arizona Press: 302-337. [171]
93. Pojar, J.; Trowbridge, R.; Coates, D. 1984. Ecosystem classification and
interpretation of the sub-boreal spruce zone, Prince Rupert Forest
Region, British Columbia. Land Management Report No. 17. Victoria, BC:
Province of British Columbia, Ministry of Forests. 319 p. [6929]
94. Raunkiaer, C. 1934. The life forms of plants and statistical plant
geography. Oxford: Clarendon Press. 632 p. [2843]
95. Reed, Robert M. 1976. Coniferous forest habitat types of the Wind River
Mountains, Wyoming. American Midland Naturalist. 95(1): 159-173. [1950]
96. Ritchie, Brent W. 1978. Ecology of moose in Fremont County, Idaho.
Wildlife Bulletin No. 7. Boise, ID: Idaho Department of Fish and Game.
33 p. [4482]
97. Rominger, James M.; Paulik, Laurie A. 1983. A floristic inventory of the
plant communities of the San Francisco Peaks Research Natural Area. Gen.
Tech. Rep. RM-96. Fort Collins, CO: U.S. Department of Agriculture,
Forest Service, Rocky Mountain Forest and Range Experiment Station. 9 p.
[2023]
98. Romme, William H. 1982. Fire and landscape diversity in subalpine
forests of Yellowstone National Park. Ecological Monographs. 52(2):
199-221. [9696]
99. Saunders, Jack K., Jr. 1955. Food habits and range use of the Rocky
Mountain goat in the Crazy Mountains, Montana. Journal of Wildlife
Management. 19(4): 429-437. [484]
100. Schlatterer, Edward F. 1972. A preliminary description of plant
communities found on the Sawtooth, White Cloud, Boulder and Pioneer
Mountains. Unpublished report. Ogden, UT: U.S. Department of
Agriculture, Forest Service, Intermountain Region. 111 p. [2076]
101. Schmidt, Wyman C.; Lotan, James E. 1980. Phenology of common forest
flora of the northern Rockies--1928 to 1937. Res. Pap. INT-259. Ogden,
UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest
and Range Experiment Station. 20 p. [2082]
102. Schmitt, Craig L. 1989. Diagnosis of annosus root disease in mixed
conifer forests in the northwestern United States. In: Otrosina, William
J.; Scharpf, Robert F., technical coordinators. Proceedings of the
symposium on research and management of annosus root disease
(Heterobasidion annosum) in western North America; 1989 April 18-21;
Monterey, CA. Gen. Tech. Rep. PSW-116. Berkeley, CA: U.S. Department of
Agriculture, Forest Service, Pacific Southwest Forest and Range
Experiment Station: 40-42. [11320]
103. Schroeder, Richard L. 1984. Habitat suitability index models: blue
grouse. FWS/OBS-82/10.81. Washington, DC: U.S. Department of the
Interior, Fish and Wildlife Service. 19 p. [11718]
104. Scott, Virgil E.; Crouch, Glenn L.; Whelan, Jill A. 1982. Responses of
birds and small mammals to clearcutting in a subalpine forest in central
Colorado. Res. Note RM-422. Fort Collins, CO: U.S. Department of
Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment
Station. 6 p. [4494]
105. Scott, Virgil E.; Whelan, Jill A.; Alexander Robert R. 1978. Dead trees
used by cavity-nesting birds on the Fraser Experimental Forest: a case
history. Res. Note RM-422. Fort Collins, CO: U.S. Department of
Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment
Station. 6 p. [4539]
106. Shearer, Raymond C. 1976. Early establishment of conifers following
prescribed broadcast burning in western larch/Douglas-fir forests. In:
Proceedings, Tall Timbers fire ecology conference and fire and land
management symposium; 1974 October 8-10; Missoula, MT. No. 14.
Tallahassee, FL:Tall Timbers Research Station: 481-500. [12499]
107. Shearer, Raymond C. 1984. Effects of prescribed burning and wildfire on
regeneration in a larch forest in northwest Montana. In: New forests for
a changing world; Proceedings, Society of American Foresters convention;
1983; Portland, OR. Washington, DC: Society of American Foresters:
266-270. [6730]
108. Smith, Ramsay. 1982. Utilization of true firs. In: Oliver, Chadwick
Dearing; Kenady, Reid M., eds. Proceedings of the biology and management
of true fir in the Pacific Northwest symposium; 1981 February 24-26;
Seattle-Tacoma, WA. Contribution No. 45. Seattle, WA: University of
Washington, College of Forest Resources: 239-242. [6869]
109. Stahelin, R. 1943. Factors influencing the natural restocking of high
altitude burns by coniferous trees in the central Rocky Mountains.
Ecology. 24(1): 19-30. [12910]
110. Stark, N. 1983. The nutrient content of Rocky Mountain vegetation: a
handbook for estimating nutrients lost through harvest and burning.
Misc. Publ. 14. Missoula, MT: University of Montana, School of Forestry,
Montana Forest and Conservation Experiment Station. 81 p. [8617]
111. Starker, T. J. 1934. Fire resistance in the forest. Journal of Forestry.
32: 462-467. [82]
112. Steele, Robert; Cooper, Stephen V.; Ondov, David M.; [and others]. 1983.
Forest habitat types of eastern Idaho-western Wyoming. Gen. Tech. Rep.
INT-144. Ogden, UT: U.S. Department of Agriculture, Forest Service,
Intermountain Forest and Range Experiment Station. 122 p. [2230]
113. Steele, Robert; Pfister, Robert D.; Ryker, Russell A.; Kittams, Jay A.
1981. Forest habitat types of central Idaho. Gen. Tech. Rep. INT-114.
Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain
Forest and Range Experiment Station. 138 p. [2231]
114. Sutton, Richard F.; Johnson, Craig W. 1974. Landscape plants from Utah's
mountains. EC-368. Logan, UT: Utah State University, Cooperative
Extension Service. 135 p. [49]
115. Tanaka, Yasuomi. 1982. Biology of Abies seed production. In: Oliver,
Chadwick Dearing; Kenady, Reid M., eds. Proceedings of the biology and
management of true fir in the Pacific Northwest symposium; 1981 February
24-26; Seattle-Tacoma, WA. Contribution No. 45. Seattle, WA: University
of Washington, College of Forest Resources: 103-111. [6768]
116. Taylor, K. L.; Fonda, R. W. 1990. Woody fuel structure and fire in
subalpine fir forests, Olympic National Park, Washington. Canadian
Journal of Forestry Research. 20: 193-199. [11518]
117. USDA Natural Resources Conservation Service. 2018. PLANTS Database, [Online].
U.S. Department of Agriculture, Natural Resources Conservation Service
(Producer). Available: https://plants.usda.gov/ [34262]
118. Viereck, L. A.; Dyrness, C. T. 1979. Ecological effects of the
Wickersham Dome Fire near Fairbanks, Alaska. Gen. Tech. Rep. PNW-90.
Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific
Northwest Forest and Range Experiment Station. 71 p. [6392]
119. Wasser, C. H.; Hess, Karl. 1982. The habitat types of Region II, U.S.
Forest Service: a synthesis. Final Report Cooperative Agreement No.
16-845-CA. Lakewood, CO: U.S. Department of Agriculture, Forest Service,
Region 2. 140 p. [5594]
120. Watson, L. E.; Parker, R. W.; Polster, D. F. 1980. Manual of plant
species suitablity for reclamation in Alberta. Vol. 2. Forbs, shrubs and
trees. Edmonton, AB: Land Conservation and Reclamation Council. 537 p.
[8855]
121. Welsh, Stanley L.; Atwood, N. Duane; Goodrich, Sherel; Higgins, Larry
C., eds. 1987. A Utah flora. Great Basin Naturalist Memoir No. 9. Provo,
UT: Brigham Young University. 894 p. [2944]
122. Williams, Clinton K.; Lillybridge, Terry R. 1983. Forested plant
associations of the Okanogan National Forest. R6-Ecol-132b. Portland,
OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest
Region. 116 p. [2566]
123. Williams, Ralph E. 1989. Distribution and impacts of annosus root
disease in forests of the northern Rocky Mountains. In: Otrosina,
William J.; Scharpf, Robert F., technical coordinators. Proceedings of
the symposium on research and management of annosus root disease
(Heterobasidion annosum) in western North America; 1989 April 18-21;
Monterey, CA. Gen. Tech. Rep. PSW-116. Berkeley, CA: U.S. Department of
Agriculture, Forest Service, Pacific Southwest Forest and Range
Experiment Station: 51-56. [11322]
124. Woodard, Paul Michael. 1977. Effects of prescribed burning on two
different-aged high-elevation plant communities in eastern Washington.
Seattle, WA: University of Washington. 228 p. Dissertation. [5350]
125. Youngblood, Andrew P.; Mauk, Ronald L. 1985. Coniferous forest habitat
types of central and southern Utah. Gen. Tech. Rep. INT-187. Ogden, UT:
U.S. Department of Agriculture, Forest Service, Intermountain Research
Station. 89 p. [2684]
126. Youngblood, Andrew P.; Mueggler, Walter F. 1981. Aspen community types
on the Bridger-Teton National Forest in western Wyoming. Res. Pap.
INT-272. Ogden, UT: U.S. Department of Agriculture, Forest Service,
Intermountain Forest and Range Experiment Station. 34 p. [2685]
127. Contreras, Glen P.; Evans, Keith E., compilers. 1986.
Proceedings--grizzly bear habitat symposium. General Technical Report
INT-207. Ogden, UT: U.S. Department of Agriculture, Forest Service,
Intermountain Research Station. 252 p. [672]
128. Day, Robert J. 1972. Stand structure, succession, and use of southern
Alberta's Rocky Mountain forest. Ecology. 53(3): 472-478. [12976]
129. Sneck, Kathleen M. Davis. 1970. The fire history Coram Experimental
Forest. Missoula, MT: University of Montana. 134 p. Thesis. [7441]
130. Vogl, Richard J.; Ryder, Calvin. 1969. Effects of slash burning on
conifer reproduction in Montana's Mission Range. Northwest Science.
43(3): 135-147. [8546]
FEIS Home Page