| FEIS Home Page |
 |
||
| A white alder stand in San Diego County, CA. Photo © 2006 Deborah Leonard. |
AUTHORSHIP AND CITATION:
Fryer, Janet L. 2014. Alnus rhombifolia.
In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service,
Rocky Mountain Research Station, Missoula Fire Sciences Laboratory (Producer).
Available: https://www.fs.usda.gov
/database/feis/plants/tree/alnrho/all.html
[].
FEIS ABBREVIATION:
ALNRHO
COMMON NAMES:
white alder
California alder
Sierra alder
TAXONOMY:
The scientific name of white alder is Alnus rhombifolia Nutt. (Betulaceae) [8,39,47,56,58,64,74]. White alder hybridizes with thinleaf alder in southwestern Idaho [86].
Common names are used throughout this Species Review. See Appendix B for a complete list of common and scientific names of plant species discussed in this review and links to other FEIS Species Reviews.
SYNONYMS: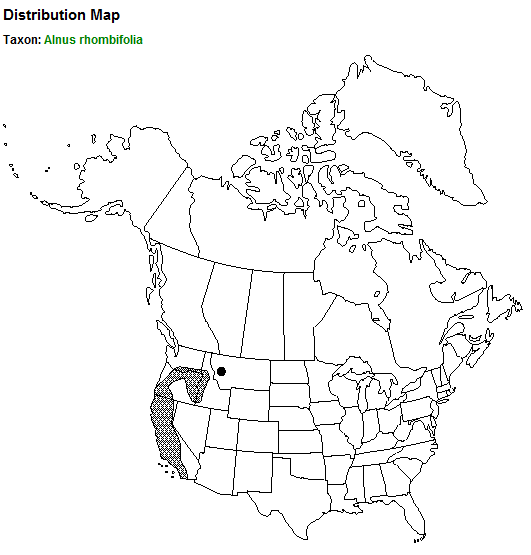 |
| Map courtesy of the Flora of North America Association [39]. (2014, August 8). |
White alder is native to the western United States; it is the most common alder in the mediterranean region of the West. It is distributed from San Diego County, California, north to Chelan County, Washington, and east to Idaho County, Idaho [73,137]. Its core distribution lies in the foothills of northern California and southwestern Oregon [73]. Isolated populations occur in extreme western Nevada [65,73] and western Montana [39]. Some report that its southern distribution probably extends to northwestern Baja California Norte [42,63], but this is undocumented and refuted by others [84].
States [137]:
CA, ID, MT, NV, OR, WA
White alder is most common along rivers and third-order streams [52,151]. In the Siskiyou Mountains of California and Oregon, it was most frequent (90%) on the most mesic sites by large streams [151]. It grows along small streams in the Coast Ranges and the Sierra Nevada [130], however. For example, white alder is most common on upper stream reaches in the Central Coast Ranges above Carmel Valley [50].
White alder is classified as a facultative riparian species [137], although it may be an obligate riparian species [51,88] in dry locations such Hell's Canyon, Idaho [82]. Anatomy of its wood vessels makes water transpiration difficult on seasonally dry sites [29]. Mature white alders are classified as very tolerant of flooding [145]. White alder also tolerates coastal fog [57].
Soils: White alder grows in sands [57], gravels [57,134], and other alluvial soils [125]. Soils are often rocky [58]. In Sequoia National Park, for example, white alder grows on alluvial soils over granodiorite bedrock [4].
White alder grows in soils derived from various parent materials. In Oregon and California, it grows in mafic soils and tolerates ultramafic soils. For example, it grows in gabbro soils in the Sierra Nevada [154]. In the Siskiyou Mountains, it grows in soils derived from olivine gabbro and diorite [151]. In the Klamath Mountains, it occurs in Port-Orford-cedar forests on ultramafics [92]. White alder is also common on nonmafic soils. On Matilija Creek in Ventura County, California, a white alder-California sycamore-coast live oak/willow-hollyleaf cherry riparian forest occurs on marine sandstones and shales [35]. In the No Business Creek Research Natural Area, Idaho, white alder grows in limestone soils. The area is a watershed of the lower Salmon River. On drainages of the Snake River, Idaho, white alder grows in soils derived from basalt and other volcanics [62].
Topography: White alder generally occupies low positions in riparian zones. It is more common on banks just above floodplains than on floodplains or upper terraces [23,118]. In the Eel River Basin of northwestern California, white alder was common on banks and low terraces but uncommon on upper terraces. White alders on upper terraces were associated with springs or tributaries [134]. In 1900, Sudworth [126] reported white alders in the Lake Tahoe area grew in "wet boggy creek bottoms", occasionally in stands as large as 0.25 acre (0.1 ha). In the Sacramento and San Joaquin deltas, white alder grows above the tidal zone [149,152].
White alder communities often occupy steep canyons, so the riparian corridor is narrow [57,62,86]. These canyons are subject to frequent slippage and erosion [86].
In southwestern Oregon, white alder is most common in riparian zones of interior valleys, especially the Willamette Valley [94]. In the Umpqua River valley, white alder is codominant in willow/field horsetail communities. These are tall-shrub, seral communities on low floodplains [131].
Elevation: White alder is most common in low-elevation montane zones [63,112,158]. At its northernmost limits, it is reported from 1,200 feet (370 m) along the Clearwater River of Idaho [63]. In southwestern Oregon, surveys on the Biscuit Burn (postfire year 4) and Bear Butte and Booth Complex Burn (postfire year 2) found white alder presence was positively associated with low elevations, low-gradient topography, large streams, well-drained organic soils, and low prefire cover of deciduous trees (P<0.05 for all variables) [52]. A study in Oregon's North Coast Ranges found white alder was positively associated with relatively low elevations, summer moisture stress, and high temperatures (P<0.01 for all variables). Elevational ranges across white alder's distribution are as follows:
| Region | Elevation (feet) |
| California | 0-8,000 [39,51]; mostly below 5,000 [57,87] |
| Idaho | 900-2,800 [62,86] |
| Nevada | 5,000-6,000 [65] |
| Oregon | 100-4,500 [63]; most common around 1,600 [112] |
| Intermountain region | typically 2,460-5,090 but up to 7,870 [58] |
| Pacific Northwest | 0-5,000 [146] |
| Siskiyou Mountains | 1,500-5,500; most common from 1,500-2,500 [151] |
Plant communities: White alder grows primarily in riparian conifer forests [48,63] and riparian oak woodlands along major streams and rivers [48]. It also grows in riparian zones of bunchgrass, sagebrush-grass [63], and chaparral communities [63,145].
Riparian communities with white alder are often diverse and historically, they were sometimes dense [9,31,91] and difficult to navigate. In Castle Crags State Park, the white alder-coastal Douglas-fir-incense-cedar/Himalayan blackberry association had highest plant species richness (70 species) and second-highest basal area (48 m²/ha) among 17 plant associations. It occurred on warm alluvial soils along Castle Creek and the Sacramento River. The white alder-bigleaf maple/vine maple association occurred along small streams; these sites were cooler and had lower species richness (37 species) [125]. Historical accounts describe California's pristine riparian forests as "jungle-like", with "impenetrable walls of vegetation" near the water [9]. Riparian associations with white alder are habitat for a wide variety of wildlife (see Importance to Wildlife and Livestock).
Many white alder communities are infested with nonnative plant species. See Other Management Considerations for further details.
Discussions of white alder communities by state follow. See Appendix B for a complete list of common and scientific names of plant species discussed in this Species Review and links to other FEIS Species Reviews. See the Fire Regime Table for a list of plant communities in which white alder may occur and information on the fire regimes associated with those communities.
California: White alder is most common in chaparral, oak woodland, and Pacific ponderosa pine zones [87]; it typically dominates riparian vegetation in these zones. In the Santa Monica Mountains, white alder-California sycamore riparian associations occur within coastal sage scrub [13]. Roberts and others [104] list white alder as uncommon in riparian forests of the North and Central Coast ranges but common in the South Coast Ranges.
White alder is an understory dominant or important component of riparian conifer and mixed-evergreen forests [14,31], including Pacific ponderosa pine, coastal Douglas-fir, mixed-conifer, and forests with more restricted geographical ranges [31]. In the southern Cascade Range and northern Sierra Nevada, it dominates the midstory of coastal Douglas-fir/white alder/Indian rhubarb forests [38]. White alder riparian forests intergrade into redwood forests in the North Coast Ranges [22] and giant sequoia forests in the Sierra Nevada [4,106,107]. In Monterey County, white alder is a subcanopy component of redwood/western bracken fern-giant chainfern streamside forests [20]. It is sometimes important in bigcone Douglas-fir-canyon live oak woodlands [78]. In late succession, incense-cedar may dominate many riparian forests with white alder [14,31].
White alder dominates or codominates many riparian hardwood forests [27,51,59,111] on the Coast [33,51,57], Transverse, and Peninsular ranges [57] and the western slope of the Sierra Nevada [33,51]. Across white alder's range in California, codominant or commonly associated tree species include Fremont cottonwood, valley oak, California sycamore, Pacific willow [9,12], bigleaf maple, California sycamore, and/or Oregon ash. Shrub and herb layers of white alder communities are sparse to well-developed [91]. California wildrose, snowberry, and Pacific poison-oak typically grow in shrub layers. White alder is common or codominant California bay woodlands and forests; these are rare communities in the Coast Ranges [19,50,97,109]. In the Santa Ana and Santa Monica mountains, white alder codominates with California sycamore and red willow in large-stream canyon bottoms. These communities occur on west slopes at midelevations (2,000-7,000 feet (600-2,000 m)) [83,144]. In the Transverse Ranges, Fremont cottonwood/white alder communities were associated with high stream flows, high elevations (~3,300 feet (1,000 m)), and relatively long times since fire (54-65 years). Willow communities with chaparral shrubs were associated with low elevations and shorter times since fire (P≤0.05) [15] (see Plant response to fire).
Extent of riparian hardwood communities generally thins in arid southern California. In the South Coast [57], Transverse, and Peninsular ranges, white alder, Fremont cottonwood, and California sycamore form narrow gallery forests along streams [85]. At low elevations in the South Coast Ranges, these riparian woodlands intergrade to arroyo willow types; white alder is characteristic in arroyo willow riparian forests [57]. In Mojave Desert riparian communities, white alder may codominate with blue palo verde and honey mesquite as well as Fremont cottonwood and California sycamore [145]. In Anza-Borrego Desert State Park, San Diego County, white alder is a component of narrowleaf willow-Fremont cottonwood-California sycamore riparian forests. These forest tracts run along Coyote Creek, which is the only perennial stream in that part of the Colorado Desert [140].
Although it prefers streambanks and riverbanks, white alder also grows on springs and seeps [31]. On the East Bay Hills, it grows in spring communities dominated by coast live oak, interior live oak, valley oak, and blue oak [2].
Pacific Northwest: Red alder is more common than white alder in most of the Pacific Northwest. White alder tends to replace red alder in southerly valleys between the North Coast and Cascade ranges in Oregon and Washington [40]. Where the 2 species cooccur in Oregon, white alder tends to grow at lower elevations [77]. In southern Oregon, white alder is a common component of mixed-conifer forests of coastal Douglas-fir, white fir, and incense-cedar [81,150]. In the Willamette Valley, white alder codominates forested floodplains with bigleaf maple, and it is characteristic in black cottonwood/scouringrush horsetail and Oregon ash/common snowberry/stinging nettle floodplain communities [77].
Idaho: In southwestern Idaho, white alder codominates riparian forests with water birch and shrubs, including Lewis' mockorange and redosier dogwood. These forests are particularly common along the lower Salmon and Snake rivers and their tributaries. They are surrounded by sagebrush steppe, mountain shrubland, and/or bluebunch wheatgrass-Idaho fescue palouse prairie [62,86]. White alder is sometimes common or codominant with netleaf hackberry in shrublands by the lower South Fork of the Salmon River [135]. It is also common or dominant along the Snake River and its tributaries. In Hell's Canyon, it grows in gallery forests that have wide gradients of elevation, channel width, and channel stability. Pure white alder stands occur on unstable channels [82]. In 1917, Weaver [148] described white alder as a "numerous secondary tree" in willow-Douglas hawthorn associations of west-central Idaho and eastern Washington.White alder is a small to medium-sized deciduous tree, usually ranging from 16 to 82 feet (5-25 m) tall [56,58]. The champion tree is 91 feet (28 m) tall and grows in Polk County, Oregon [3]. In California, white alders commonly reach 11 inches (28 cm) DBH but may reach 21 inches (53 cm) DBH [19]. The crown is conical, spreading, and open [39]; a mature tree may have one to several stems [42,63]. The bark is thin [58]. Leaves are broadly ovate; water-stressed trees produce smaller and fewer leaves than trees on moist sites [33]. The inflorescences are catkins. Staminate catkins grow in clusters of 3 to 7; pistillate catkins may be solitary or grow in racemose [58] clusters of 2 to 6 [39]. After fertilization, pistillate catkins become cone-shaped and woody [115]. The fruit is a leathery, irregularly-shaped samara [39,58] containing a small nut [53,58,157]. Samara wings are narrow, and the samara is light-weight [53].
 |
 |
| White alder staminate catkins, pistillate catkins, and seeds. Photos by C. Stubler, M. Ritter, W. Mark, and J. Reimer,
courtesy of selectree.calpoly.edu. |
|
Roots of mature white alders can withstand inundation by most large stream flows [72]. Scouring flows may expose the roots. In Humboldt County, California, white alders were rooted ≤1.3 feet (0.4 m) above the low-water line of streambanks. Roots of white alder, willows, and black cottonwood were intertwined and dense [72].
 |
| Exposed white alder roots. Photo by Sevenoaks Native Nursery. |
White alder's root nodules are infected with nitrogen-fixing bacteria in the genus Frankia [93,98], so white alder may increase soil nitrogen levels [21,53].
Stand structure: White alder communities vary from open woodlands to closed forests. In foothills of the Sierra Nevada, crown cover is typically continuous [91]. The canopy may thin toward white alder's southern distribution [57]; however, some southern locations support dense white alder forests. In the Eel River Basin of northwestern California, a white alder population showed normal distribution of size classes, so the authors suggested that most white alders established around the same time. Most trees (10%-38% frequency) were 10 to 20 inches (30-50 cm) DBH. Density averaged 0.15 tree/m of stream channel segment [134]. By the Nacimiento River of coastal central California, white alder ranged from 75% to 95% cover along narrow channels and had good representation in all size classes [118]. In Cleghorn Canyon on the San Bernardino National Forest, white alder forests had a closed canopy with all size classes well represented. The canopy was "well over" 9 feet (3 m) tall [31].
Raunkiaer [101]
life form:
Phanerophyte
Geophyte
SEASONAL DEVELOPMENT:
White alder forms catkins the summer prior to flowering [39,56,58]. Flowering occurs before spring stem and leaf growth starts [58]; white alder sometimes flowers in December on the Central Coast Ranges [50]. The conelike fruits dry, open, and disperse most seed in summer and fall, although some seed may remain in "cones" through winter [53]. Pistillate cones are retained through winter. Leaves drop before winter in most of white alder's range, although they are retained through most of the winter in southern California [42,115].
| Phenology of white alder across its range | ||
| Region | Event | Period |
| California | flowers | January-April [8,87] |
| Nevada | flowers | February-April [65] |
| Oregon | flowers | March [53] |
| fruits | late September-early October [53] | |
| Intermountain region | flowers | April-May [58] |
| fruits | late May [58] | |
| Pacific Northwest | flowers | January-April [56] |
REGENERATION PROCESSES:
White alder regenerates from seeds and sprouts [110,118]. Seeds are important for colonizing new areas such as sand- and gravelbars, and established plants may sprout after bole damage or top-kill [110,118]. By the Nacimiento River, white alder showed good regeneration from both seeds and sprouts [118].
Pollination and breeding system: White alder is wind-pollinated [115] and monoecious [42,115,157].
Seed production: Seed production is generally good [65].
Seed dispersal: Wind [26,53,157], water [26,66], and seed-eating birds [53] disperse white alder seeds.
Seed banking: White alder has a transient soil seed bank, lasting a few months [37]. High flows carry away much seed deposited on banks [66]. Undispersed seed retained in crowns may remain viable though winter. Seed viability is highest in the upper third of crowns [53].
Germination: Seeds germinate rapidly on sunny, wet mineral soils exposed or deposited by receding floodwaters [26]. Fresh seed is immediately germinable [37,157], and seeds may remain viable for a few months [37]. Three-month stratification increases germination rates in the laboratory [37,53], but seed viability drops from fall through spring in the field [53].
When damming and flood control disrupt natural seed dispersal and time of flooding, so tamarisk, tree-of-heaven, and Russian-olive often germinate more successfully than white alder and cottonwoods [91].
Seedling establishment and plant growth: Alder seeds have small cotyledons and lack endosperm [53], so germination and establishment must proceed quickly or germinants will die.
Seedling establishment appears restricted to sites with a continuously moist substrate. White alder seedlings apparently cannot survive on sites that dry out during the summer [26]. White alder usually establishes on banks after floodwaters recede, typically just above active bars [72]. For stands to develop, white alder requires a stable substrate above the scour zone and a shallow rooting depth to year-round moisture. Once established, stands can survive most spring inundations [72]. On 4 sites across western Oregon, most (10% frequency) white alder seedling establishment was on streamside plots, and almost none (~1% frequency) was on upland plots. Density on streamside plots ranged from 109 to 11,227 seedlings/ha. Scouring was the most important factor deterring establishment, with presence of white alder seedlings positively associated with low-scour channels (R²=0.08) [113]. In a common garden study, nearly all seedlings subject to moderate water stress (stomal closure and loss of turgor after 5 days without water) survived, while over half of those subject to severe water stress (leaf loss after 10 days without water) died [33].
Drought [112] and plant removal by channel scouring [75] are the main causes of seedling and sapling mortality. Although mature white alders can survive inundation [112], high flows may remove white alders of any age [75,109]. In the channel of Dry Creek in Sonoma County, California, high flows from storms in winter 1981-1982 eroded a gravelbar into an island, flushing white alder saplings downstream. A few adult trees on the channel margin also fell into the stream. On intact gravelbars, however, survivorship of 2- to 5-year old and 5+-year-old saplings was 100% from June 1981 to June 1982 [75]. In the San Bernardino Mountains, winter floods in 1969 removed entire white alder stands [83].
Mule deer browsing can reduce growth or cause mortality in all age classes. On second- and third-order streams in Mendocino County, California, white alder saplings had greater mean density in deer exclosure plots (0.5 sapling/m²) than in open control plots (0.05 sapling/m²). Saplings in exclosures were also taller (65% were >3 feet (1 m)) than saplings in controls (97% were <3 feet) [95].
White alder generally grows quickly. However, growth is slow and plants are small on sites with fluctuating soil moisture in summer. In Long Canyon on the Sequoia National Forest, white alders by streams with sparse summer flows grew no more than 20 feet (6 m) tall [31].
Vegetative regeneration: White alder sprouts from the root crown [42,110], and/or roots [110]. It is apparently better able to sprout after flooding [17] or top-killing frost [42] than after fire [17,35] (see Plant response to fire).
White alder also layers, although layering is rarely documented. A study by Dry Creek found that white alders growing on alluvial sediments reproduced mostly by layering. Trees on terraces did not layer [76].
SUCCESSIONAL STATUS:White alder establishes during both primary [26,72,76] and secondary [157] succession. In primary succession, it colonizes sandbars and other fresh alluvium exposed or deposited by receding floodwaters [26,72,76]. Such succession typically happens on only small patches of a watercourse [91]. White alder's ability to fix nitrogen helps it establish on young, nutrient-poor soils [53].
In secondary succession, white alder is noted on slumps [31] and burns (see Plant response to fire). However, it is sensitive to interference from other sprouting hardwood trees and may not establish well after fire if other hardwoods were present before fire [17]. On the Biscuit Burn and the Bear Butte and Booth Complex Burn, white alder presence in early postfire communities (postfire years 4 and 2, respectively, for each burn) was positively associated with low prefire cover of deciduous trees (P<0.05) [52]. |
||
| White alder snags 13 months after the Wolf Wildfire. The wildfire killed most white alders, but coast live oaks sprouted. Photo courtesy of Jacob Bendix, Syracuse University. |
Immediate fire effects on plant:
Severe fire kills white alder. Some trees may survive low-severity fires [35]. Exposed
roots
leave white alder especially vulnerable to fire kill.
Postfire regeneration strategy [124]:
Tree with adventitious buds,
a sprouting root crown,
and root sprouts
Tall shrub, adventitious buds, a sprouting root crown, and root sprouts
Geophyte, growing points deep in soil
Ground residual colonizer (on site, initial community)
Crown residual colonizer (on site, initial community)
Initial off-site colonizer (off site, initial community)
Secondary colonizer (on- or off-site seed sources)
Fire adaptations and plant response to fire:
In the Sespe Wilderness, California, white alder established after a stand-replacement fire on the Sespe Wilderness, California, but not after another stand-replacement fire 70 years later. There was some white alder recruitment from 1935-1940, following the 1932 Matilija Wildfire. There was "continuous, but irregular" recruitment from 1966-2000. Whether these waves of recruitment were from sprouts or seeds could not be determined. The 2002 Wolf Wildfire killed white alders on 11 study sites, and white alder had not reestablished as of 2002. Peak flows on the 11 sites were low from 1933-1968, with 6 large floods from 1969-1983. The largest floods were in 1969, 1978, and 1983. White alder showed good recruitment in the 1970s, following the 1969 flood, and steady recruitment in the 1980s and 1990s. The authors suggested that white alder will likely reestablish on the burned sites in decades to come. See Fire regimes for further information on this study [18].
The Regeneration Processes section provides detailed information and documentation on white alder regeneration.
Plant response to fire: White alder plants are not fire resistant [35]. They apparently have sparse postfire seedling establishment [139] and a weak postfire sprouting response. White alder may require decades to recover after severe fire [17,18,35], especially if fire has killed parent trees [35]. In 3 riparian communities on the Lassen National Forest, California, white alder dominance was positively associated with time since fire (P=0.009) [108]. However, white alder escapes many fires because it is restricted to stream- and riverbanks that burn less frequently than surrounding uplands [128]. Along the San Gabriel River in California, Brothers [26] observed no evidence of fire damage on large white alder trees, which were older than the last major fire in the area.
One author stated that white alder is top-killed or survives fire, although no examples or studies were cited [66].
Limited studies show that associated hardwoods have stronger ability to sprout after fire [12,17,18,35], outcompeting white alder for space and light on streambanks. In the Transverse Ranges, the 2002 Wolf Wildfire crowned in 2 white alder-coast live oak/Eastwood's manzanita galleries [16,17]. Overstory trees included white alder, coastal sage scrub oak, and Fremont cottonwood. Most white alders died; the other hardwoods were mostly top-killed. Surveys in postfire year 3 found few white alders sprouting, and coast live oak sprouts dominated the postfire community. White alder had the weakest sprouting response of all overstory trees, and the authors suggested there was little opportunity for white alder to establish by seed due to prolific postfire sprouting of associated hardwoods [17].
| Relative abundance of dominant overstory trees before and 3 years after the 2002 Wolf Wildfire in the Transverse Ranges [17] | ||||||||
| Species | ||||||||
| Relative basal area (%) | Relative density (%) | Relative frequency (%) | Importance value** | Relative basal area (%) | Relative density (%) | Relative frequency (%) | Importance value | |
| White alder | 56.5 | 29.4 | 29.9 | 38.6 | 0.2 | 4.4 | 10.0 | 4.8 |
| Coast live oak | 18.2 | 3.0 | 5.7 | 9.0 | 91.2 | 26.1 | 30.0 | 49.1 |
| *Prefire stand structure determined from postfire snags. **Mean of relative basal area, relative density, and relative frequency. |
||||||||
Three years after the high-severity Wheeler Gorge Wildfire in summer 1985, surveys found that about 97% of white alder trees on the upper Santa Ynez River basin had died. The prefire plant community was white alder-California sycamore-Fremont cottonwood-coast live oak/currant-rose. There was no correlation between white alder diameter and fire-caused mortality [12].
| Effects of the 1985 Wheeler Gorge Wildfire on white alders >10 cm in diameter as of postfire year 3 [11] | ||||
| Canyon | Total # trees sampled | # alders (% of total) |
Dead alders (%) |
Dead fallen alders (%) |
| Jancal | 73 | 49 (67) | 96 | 18 |
| Upper North Fork Matilija | 125 | 71 (57) | 97 | 37 |
In contrast to white alder's poor sprouting response, most associated oaks, California sycamores, and Fremont cottonwoods had only been top-killed and were sprouting. The few white alders that were only top-killed sprouted several months after the other hardwoods ([12], Barro 1989 personal communication [11]). By spring of postfire year two, 7% percent of burned white alders were sprouting compared to 83% of California sycamores and 70% of coast live oaks [35]. Abundance of white alder and other hardwoods in postfire year 3 was [12]:
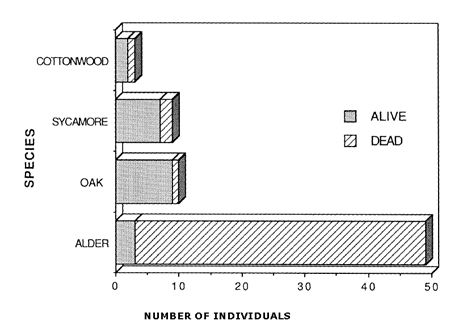 |
| Abundance of overstory tree species on the upper Santa Ynez River basin, 3 years after the 1985 Wheeler Gorge Wildfire [12]. |
Coring of white alder snags on the Wheeler Gorge Burn showed a cohort of white alders that established after a 1969 flood, but none that established after fire (Barro 1989 personal communication [11]). White alder seed rain was low through postfire year 3. There were no white alder seedlings in burned plots by then, and <1% of white alder seeds collected on burned plots were viable. However, there were white alder seedlings in unburned plots just downstream. Some of those seedlings were 10 feet (3 m) tall by postfire year 3, but others had been uprooted by postfire winter storms. The authors suggested that due to the scarcity of viable seed, white alder will likely take decades to fully recover from the wildfire. They also suggested that the postfire storms were typical, not severe, and that the amount of channel erosion was typical of early postfire chaparral ecosystems. A storm on 30-31 January 1986 had produced a streamflow of 2.1 m³/second in the study reach. A second storm on 13 to 15 February 1986 produced a stream flow of 2.5 m³/second. These storms widened the stream channel by 1.6 feet (0.5 m) and deepened it by 2.3 feet (0.7 m) [35].
As that study shows [35], fire can have indirect effects on white alder and other riparian species by altering fluvial processes on watersheds. Increased water flow, erosion, debris, sediment, and nutrient loads are common after fire above or within riparian zones ([141], review by [36]), and these effects may last from several postfire months to 4 years in mediterranean climates [141]. Postfire erosion of stream channels can remove white alders that survived a fire. The prefire white alder-California sycamore-coast live oak riparian forest that burned in the Wheeler Gorge Fire was surrounded by chaparral. After the 2 storms in January and February 1986, 8 of 23 surviving white alders were uprooted, and 2 more were uprooted in the next 2 years [35].
Postfire browsing can reduce white alder seedling establishment and sprout growth (see Mule deer browsing).
FUELS AND FIRE REGIMES:On riparian rangelands, livestock grazing removes many fine fuels [102]. Oak woodlands with annual grass ground layers are often used for cattle grazing, and cattle often seek out white alder communities surrounded by oak woodlands. On an oak rangeland in Tehama County, California, most surface fuels were grasses. By Dye Creek, crews measured litter loads, woody debris loads, and stand structure of an interior live oak-valley oak-white alder/California wild grape-Pacific poison oak-Himalayan blackberry riparian forest. The area was used for cattle grazing; mule deer, feral pigs, and black-tailed jackrabbits were other important foragers. Results of the survey were [132]:
| Fuel and stand structure characteristics of a riparian forest in Tehama County, California. Data were collected in spring (SD) [132]. | |
| Litter depth (cm) | 1.3 (1.3) |
| Canopy height (m) | 4.1 (3.8) |
| Canopy closure (%) | 25.2 (18.7) |
| Basal area (m²/ha) | 7.8 (7.4) |
| Vertical strata cover (%) | |
| Ground level (0.0-0.5 m) | |
water |
0.7 (1.6) |
rock |
16.9 (11.4) |
bare soil |
10.3 (6.4) |
litter |
18.5 (11.2) |
dead wood |
1.4 (1.8) |
moss |
1.2 (2.8) |
grass |
26.3 (18.1) |
forbs & ferns |
18.5 (9.8) |
live wood |
0.6 (0.7) |
evergreen broadleaf foliage |
0.6 (0.9) |
deciduous broadleaf foliage |
5.2 (7.2) |
| Low strata (2-5 m above ground) | |
dead wood |
0.1 (0.3) |
live wood |
1.1 (1.2) |
evergreen broadleaf foliage |
3.4 (4.5) |
deciduous broadleaf foliage |
4.8 (4.4) |
| Mid strata (10-15 m above ground) | |
dead wood |
<0.1 (0.2) |
live wood |
0.5 (1.0) |
evergreen broadleaf foliage |
0.2 (0.5) |
deciduous broadleaf foliage |
3.0 (4.5) |
| Subcanopy strata (20-30 m above ground) | |
dead wood |
0.0 (0.0) |
live wood |
0.1 (0.4) |
evergreen broadleaf foliage |
0.0 (0.0) |
deciduous broadleaf foliage |
0.2 (1.1) |
| Total live plant cover | 83.9 (27.5) |
Brooks and others [25] report that in desert riparian communities, surface fuel loads and fuel continuity can by "very high". This facilitates spread of surface fire; however, ladder and crown fuels are often discontinuous, impeding spread of crown fire [25].
Several fuel metrics are available for white alder. Fuelwood characteristics—including energy output, specific gravity, and moisture content—are provided in Wilson and others (review [155]). Specific leaf area can be used to calculate foliar fuel loads [116]. Studies in California found the specific leaf area of white alder averaged 10.33 m²/kg [143].
Fire regimes: Moderate fire-return intervals (50-70 years) favor white alder [15,17]. Although white alder occurs in the riparian zones of many plant communities that have short (<50 years) fire-return intervals—such as palouse prairie [5], chaparral [96], oak woodlands [1], and Pacific ponderosa pine [5]—riparian areas with white alder have longer fire-return intervals than surrounding uplands in most years [102,120]. These riparian zones are often either too moist to burn ([68], review by [41]) or burn at lower severity than uplands. Some estimate that fire-return intervals in western riparian zones were historically several times longer than those in surrounding uplands ([119], review [41]). Riparian areas by first- and second-order streams may burn at intervals similar to those of surrounding uplands [30], although white alder is less prevalent by small streams (see Site Characteristics).
Historical fire severity in riparian zones is not well known. Some suggest that typically, fires that burned at high or moderate severity in uplands burned at moderate or low severity in riparian zones [17,68]. Other suggest that when riparian areas burned, fires were probably of moderate to high severity [102]. A pattern of frequent fires of at least moderate severity is emerging in some riparian areas. Landsat data from Yosemite National Park showed that from 1984 to 2003, fire severity was moderate to high in 75% of black cottonwood-white alder-bigleaf maple riparian communities that burned; 904 acres (366 ha) or 5.1% of the Park's total riparian area burned during that time. Some riparian communities burned 2 or 3 times in 20 years. The authors placed median severity of riparian fires "at the high end of the moderate category" [130].
Riparian communities can have severe, stand-replacement fires in dry years [17]. Wind funneling down water channels can sustain and increase fire intensity in drought-stressed riparian zones [117]. During droughts, fire severity in riparian zones is strongly influenced by fire severity in upland zones [17]. In the Transverse Ranges, the 2002 Wolf Wildfire was stand-replacing in adjacent upland chamise-manzanita stands. The fire remained severe and stand-replacing when it burned into the white alder-coast live oak community on the floodplain of Piedra Blanca Creek [17].
Discussions and documentation of fire regimes by state follow. See the Fire Regime Table for further information on fire regimes of vegetation communities in which white alder may occur. Find further fire regime information for the plant communities in which this species may occur by entering the species name in the FEIS home page under "Find Fire Regimes".
California: Little is known of historical fire regimes of California's riparian areas [102,117,120], although historical fire-return intervals are thought to be longer in California's riparian zones than in upland zones [102]. Presently, fire-return intervals in riparian zones of the Klamath Mountains and the southern Cascade Range are twice those of adjacent uplands [117,120]. Fires miss riparian areas surrounded by highly flammable vegetation in some years. In the Santa Monica Mountains, the 1970 Maliby Wildfire and the 1973 Mugu and Topanga-Tuna Canyon wildfires were stand-replacing in coastal sage scrub and chaparral vegetation. However, the wildfires did not burn white alder stands in deep canyons [114].
In the Transverse Ranges, median fire-return interval of riparian communities was 60 years [15,17]. White alder communities had not burned for about 54 to 65 years. Willow communities with chaparral shrubs were associated with shorter times since fire (P≤0.05). Because white alder is fire-sensitive (see Plant response to fire), the author concluded that short fire-return intervals excluded white alder from willow-chaparral communities [15]. In chaparral areas near Santa Barbara, return intervals for large fires historically ranged from 30 to 65 years [28]. It is uncertain how often these large fires burned into riparian areas. In the Sespe Wilderness, 11 riparian sites on the Sespe Creek Watershed burned in large wildfires in 1932 [18] (Matilija Wildfire, 220,000 acres (89,100 ha)) [67] and 2002 (Wolf Wildfire, 21,600 acres (8,760 ha)). Two of the 11 sites also burned in a smaller fire in 1975 (in Potero John Creek watershed, ~100 acres (43 ha)). Because this study was conducted by small streams (< 35 km²) drainage area) and only 29 white alders were cored to determine stand ages, the authors cautioned that these results cannot be extrapolated across all white alder communities [18].
Skinner and others [120] stated that California's riparian areas were important topographic features affecting fire spread. In most years, riparian forests were probably barriers to spread of low-severity fires. Relatively moist conditions probably stopped spread of some moderate-severity fires as well. In many years, this barrier effect likely reduced fire severity or occurrence beyond the riparian zone [120].
Oregon: As of 2014, little documentation was available on fire regimes of Oregon's riparian zones containing white alder. A fire history study in in Rouge River Basin showed a regime of mixed-severity fires at moderate intervals. The mixed-conifer forests investigated were coastal Douglas-fir-white fir-incense-cedar/bigleaf maple, with white alder "a major overstory tree" on upland sites (67% frequency) and an important component of the forest in interior valley sites (23% frequency). Recruitment of overstory trees occurred in pulses, with cohorts establishing every 50 to 60 years. The oldest coastal Douglas-firs were 200+ years old. Fire scars on these trees recorded 4 fires on uplands from approximately 1750 to 1910 and 6 fires in interior valleys from 1850 to 1925 [81].
Idaho: Little documentation was available on fire regimes of Idaho's riparian zones containing white alder. In Hell's Canyon, fires are frequent in low-elevation bluebunch wheatgrass-Idaho fescue palouse prairies and adjacent riparian zones [86] where white alder may be present [82,86].
FIRE MANAGEMENT CONSIDERATIONS:Late October prescribed fires on the Blodgett Forest Research Station, California, burned at low to moderate severity and had little effect on riparian trees. Study sites were mixed-conifer forests with white alder and Pacific yew adjacent to first- and second-order streams. In postfire year 1, litter and woody debris loads were about 20% less than a year before burning. Overall mortality of canopy trees was low (4.4%), so the fires did not significantly reduce canopy cover. Data for white alder alone were not provided [14].
Riparian deciduous forests are important buffer strips. In postfire environments, they help protect aquatic zones by capturing sediment flows and nutrients from burned uplands [36]. To protect fisheries, Neary and others [90] recommended that prescribed fires in riparian zones be of low severity, with flame lengths ≤2 feet (0.6 m).
As an important member of riparian woodland communities, white alder contributes to structural diversity and provides cover and food—all important habitat requirements for many bird species. In California, deciduous riparian woodlands support a higher diversity and density of breeding birds than other habitats [43,49,55,71]. For example, an 8-year survey on the Sierra National Forest, California, found that 13 bird species, including Cooper's hawk and several woodpecker, hummingbird, and flycatcher species, used white alder as nest trees [100]. Redpolls, siskins, and goldfinches eat alder seeds [136].
Riparian forests with white alder are important habitat for threatened and rare species [23] including the least Bells' vireo [49] and mountain yellow-legged frog [123]. In Anza-Borrego Desert State Park, riparian forests provide critical habitat for the least Bell's vireo and peninsular bighorn sheep [140]. Foothill riparian hardwoods are a minor habitat type for California spotted owls [142]. Southwestern willow flycatchers use riparian areas as habitat and white alders for nesting [139]. The valley elderberry longhorn beetle is an obligate feeder on elderberries, which are common in riparian zones. The beetle is most abundant in diverse riparian communities with mature overstories and dense understories; white alder is often a component of such communities [138].
Caddisfly larvae feed on white alder litter in streams [45].
Palatability and nutritional value: Ungulate use of white alder browse is rated poor to fair for mule deer and domestic sheep, unused to poor for cattle, and unused for horses [110].
Cover value: When growing with other riparian trees, white alder contributes to structural diversity for many wildlife species and provides cover important for numerous perching birds [49]. Livestock seek riparian zones for shade and resting [127].
Nesting birds use living and snag white alders. Cavity nesters such as pileated woodpeckers [54] and red-breasted nuthatches [7] sometimes select white alders. On 54- to ~70-year-old burns on the Sierra National Forest, woodpecker, nuthatch, and owl species apparently nested in white alder snags as much as expected based on availability [7].
White alder provides shade for fish and other aquatic animals [36,121].
VALUE FOR REHABILITATION OF DISTURBED SITES:For information on artificial regeneration and seedling care of white alder, see these sources: [53,122,157]. Commercial plant sources are available [44].
OTHER USES:California Indians used white alder medicinally and to make dyes [34,129,133]. They used the wood for making utensils and as firewood [133].
OTHER MANAGEMENT CONSIDERATIONS:Many extant riparian systems have been greatly altered, especially where placer mining was conducted. Stream channeling and reduction of American beaver populations have also altered water courses. Historically, stream channels were broader and more braided. Shaffer and others [117] suggest that although the fire-return interval may not have changed due to altered stream flows, fires probably move through valley bottoms and low-gradient riparian zones differently than they did historically.
Invasive nonnative plant species may reduce cover of white alder and other native riparian species and lower overall diversity of native riparian plant species. Nonnative plants common in many riparian communities with white alder include giant reed [158], Himalayan blackberry [125], tamarisk, Russian-olive, tree-of-heaven [91,103], and black locust [99]. In Hell's Canyon, some white alder/Lewis' mockorange communities are infested with Himalayan blackberry and/or bittersweet nightshade [86]. In the Santa Monica Mountains of southern California, nonnative invasive annual and perennial herbs such as black mustard, ripgut brome, and horehound dominated the ground layer of a white alder-California bay woodland in Solstice Canyon. Residual native annuals included caterpillar phacelia and hollowleaf annual lupine [97].
Climate change modeling predicts that white alder's climatic niche will shift 9.7° northward in latitude. Among 25 tree species in the United States, it was the only western tree with such a large predicted change in distribution [79].| Fire regime information on vegetation communities in which white alder may occur. This information is taken from the LANDFIRE Rapid Assessment Vegetation Models [70], which were developed by local experts using available literature, local data, and/or expert estimates. This table summarizes fire regime characteristics for each plant community listed. The PDF file linked from each plant community name describes the model and synthesizes the knowledge available on vegetation composition, structure, and dynamics in that community. Cells are blank where information is not available in the Rapid Assessment Vegetation Model. In many cases, white alder riparian zones are imbedded in the vegetation communities listed below, and fire regime characteristics in these riparian zones may differ from those described for upland models. | ||||||||
|
||||||||
| Pacific Northwest | ||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | ||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
|||||
| Pacific Northwest Grassland | ||||||||
| Bluebunch wheatgrass | Replacement | 47% | 18 | 5 | 20 | |||
| Mixed | 53% | 16 | 5 | 20 | ||||
| Idaho fescue grasslands | Replacement | 76% | 40 | |||||
| Mixed | 24% | 125 | ||||||
| Pacific Northwest Shrubland | ||||||||
| Low sagebrush | Replacement | 41% | 180 | |||||
| Mixed | 59% | 125 | ||||||
| Mountain big sagebrush (cool sagebrush) | Replacement | 100% | 20 | 10 | 40 | |||
| Wyoming big sagebrush steppe | Replacement | 89% | 92 | 30 | 120 | |||
| Mixed | 11% | 714 | 120 | |||||
| Pacific Northwest Woodland | ||||||||
| Oregon white oak | Replacement | 3% | 275 | |||||
| Mixed | 19% | 50 | ||||||
| Surface or low | 78% | 12.5 | ||||||
| Oregon white oak-ponderosa pine | Replacement | 16% | 125 | 100 | 300 | |||
| Mixed | 2% | 900 | 50 | |||||
| Surface or low | 81% | 25 | 5 | 30 | ||||
| Ponderosa pine | Replacement | 5% | 200 | |||||
| Mixed | 17% | 60 | ||||||
| Surface or low | 78% | 13 | ||||||
| Subalpine woodland | Replacement | 21% | 300 | 200 | 400 | |||
| Mixed | 79% | 80 | 35 | 120 | ||||
| Pacific Northwest Forested | ||||||||
| California mixed evergreen (northern California and southern Oregon) | Replacement | 6% | 150 | 100 | 200 | |||
| Mixed | 29% | 33 | 15 | 50 | ||||
| Surface or low | 64% | 15 | 5 | 30 | ||||
| Douglas-fir (Willamette Valley foothills) | Replacement | 18% | 150 | 100 | 400 | |||
| Mixed | 29% | 90 | 40 | 150 | ||||
| Surface or low | 53% | 50 | 20 | 80 | ||||
| Douglas-fir-western hemlock (dry mesic) | Replacement | 25% | 300 | 250 | 500 | |||
| Mixed | 75% | 100 | 50 | 150 | ||||
| Douglas-fir-western hemlock (wet mesic) | Replacement | 71% | 400 | |||||
| Mixed | 29% | >1,000 | ||||||
| Mixed conifer (eastside mesic) | Replacement | 35% | 200 | |||||
| Mixed | 47% | 150 | ||||||
| Surface or low | 18% | 400 | ||||||
| Mixed conifer (southwestern Oregon) | Replacement | 4% | 400 | |||||
| Mixed | 29% | 50 | ||||||
| Surface or low | 67% | 22 | ||||||
| Oregon coastal tanoak | Replacement | 10% | 250 | |||||
| Mixed | 90% | 28 | 15 | 40 | ||||
| Ponderosa pine, (mesic) | Replacement | 5% | 125 | |||||
| Mixed | 13% | 50 | ||||||
| Surface or low | 82% | 8 | ||||||
| California | ||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | ||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
|||||
| California Grassland | ||||||||
| California grassland | Replacement | 100% | 2 | 1 | 3 | |||
| California Shrubland | ||||||||
| Chaparral | Replacement | 100% | 50 | 30 | 125 | |||
| Coastal sage scrub | Replacement | 100% | 50 | 20 | 150 | |||
| Coastal sage scrub-coastal prairie | Replacement | 8% | 40 | 8 | 900 | |||
| Mixed | 31% | 10 | 1 | 900 | ||||
| Surface or low | 62% | 5 | 1 | 6 | ||||
| Montane chaparral | Replacement | 34% | 95 | |||||
| Mixed | 66% | 50 | ||||||
| California Woodland | ||||||||
| California oak woodlands | Replacement | 8% | 120 | |||||
| Mixed | 2% | 500 | ||||||
| Surface or low | 91% | 10 | ||||||
| Ponderosa pine | Replacement | 5% | 200 | |||||
| Mixed | 17% | 60 | ||||||
| Surface or low | 78% | 13 | ||||||
| California Forested | ||||||||
| Aspen with conifer | Replacement | 24% | 155 | 50 | 300 | |||
| Mixed | 15% | 240 | ||||||
| Surface or low | 61% | 60 | ||||||
| California mixed evergreen | Replacement | 10% | 140 | 65 | 700 | |||
| Mixed | 58% | 25 | 10 | 33 | ||||
| Surface or low | 32% | 45 | 7 | |||||
| Coast redwood | Replacement | 2% | ≥1,000 | |||||
| Surface or low | 98% | 20 | ||||||
| Jeffrey pine | Replacement | 9% | 250 | |||||
| Mixed | 17% | 130 | ||||||
| Surface or low | 74% | 30 | ||||||
| Interior white fir (northeastern California) | Replacement | 47% | 145 | |||||
| Mixed | 32% | 210 | ||||||
| Surface or low | 21% | 325 | ||||||
| Mixed conifer (north slopes) | Replacement | 5% | 250 | |||||
| Mixed | 7% | 200 | ||||||
| Surface or low | 88% | 15 | 10 | 40 | ||||
| Mixed conifer (south slopes) | Replacement | 4% | 200 | |||||
| Mixed | 16% | 50 | ||||||
| Surface or low | 80% | 10 | ||||||
| Mixed evergreen-bigcone Douglas-fir (southern coastal) | Replacement | 29% | 250 | |||||
| Mixed | 71% | 100 | ||||||
| Red fir-white fir | Replacement | 13% | 200 | 125 | 500 | |||
| Mixed | 36% | 70 | ||||||
| Surface or low | 51% | 50 | 15 | 50 | ||||
| Sierra Nevada lodgepole pine (cold wet upper montane) | Replacement | 23% | 150 | 37 | 764 | |||
| Mixed | 70% | 50 | ||||||
| Surface or low | 7% | 500 | ||||||
| Northern and Central Rockies | ||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | ||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
|||||
| Northern and Central Rockies Grassland | ||||||||
| Mountain grassland | Replacement | 60% | 20 | 10 | ||||
| Mixed | 40% | 30 | ||||||
| Northern and Central Rockies Shrubland | ||||||||
| Mountain big sagebrush steppe and shrubland | Replacement | 100% | 70 | 30 | 200 | |||
| Mountain shrub, nonsagebrush | Replacement | 80% | 100 | 20 | 150 | |||
| Mixed | 20% | 400 | ||||||
| *Fire Severities— Replacement: Any fire that causes greater than 75% top removal of a vegetation-fuel type, resulting in general replacement of existing vegetation; may or may not cause a lethal effect on the plants. Mixed: Any fire burning more than 5% of an area that does not qualify as a replacement, surface, or low-severity fire; includes mosaic and other fires that are intermediate in effects. Surface or low: Any fire that causes less than 25% upper layer replacement and/or removal in a vegetation-fuel class but burns 5% or more of the area [10,69] |
| APPENDIX B: Common and scientific names of plant species | |
| Trees | |
| alder | Alnus spp. |
| bigcone Douglas-fir | Pseudotsuga macrocarpa |
| bigleaf maple | Acer macrophyllum |
| black cottonwood | Populus balsamifera subsp. trichocarpa |
| black locust | Robinia pseudoacacia |
| blue oak | Quercus douglasii |
| blue palo verde | Parkinsonia florida |
| California bay | Umbellularia californica |
| California sycamore | Platanus racemosa |
| canyon live oak | Quercus chrysolepis |
| coastal Douglas-fir | Pseudotsuga menziesii var. menziesii |
| coastal sage scrub oak | Quercus dumosa |
| coast live oak | Quercus agrifolia |
| cottonwood | Populus spp. |
| Fremont cottonwood | Populus fremontii |
| giant sequoia | Sequoiadendron giganteum |
| honey mesquite | Prosopis glandulosa |
| incense-cedar | Calocedrus decurrens |
| interior live oak | Quercus wislizeni |
| narrowleaf willow | Salix exigua |
| oak | Quercus spp. |
| Oregon ash | Fraxinus latifolia |
| Pacific ponderosa pine | Pinus ponderosa var. ponderosa |
| Pacific yew | Taxus brevifolia |
| Port-Orford-cedar | Chamaecyparis lawsoniana |
| red alder | Alnus rubra |
| red willow | Salix laevigata |
| redwood | Sequoia sempervirens |
| Russian-olive | Elaeagnus angustifolia |
| tamarisk | Tamarix spp. |
| thinleaf alder | Alnus incana subsp. tenuifolia |
| tree-of-heaven | Ailanthus altissima |
| valley oak | Quercus lobata |
| water birch | Betula occidentalis |
| white fir | Abies concolor |
| willow | Salix spp. |
| Shrubs | |
| arroyo willow | Salix lasiolepis |
| bittersweet nightshade | Solanum dulcamara |
| California wildrose | Rosa californica |
| common snowberry | Symphoricarpos albus |
| currant | Ribes spp. |
| Douglas hawthorn | Crataegus douglasii |
| Eastwood's manzanita | Arctostaphylos glandulosa |
| elderberries | Sambucus spp. |
| Himalayan blackberry | Rubus discolor |
| hollyleaf cherry | Prunus ilicifolia |
| Lewis' mockorange | Philadelphus lewisii |
| netleaf hackberry | Celtis reticulata |
| Pacific poison-oak | Toxicodendron diversilobum |
| Pacific willow | Salix lasiolepis |
| red elderberry | Sambucus racemosa |
| redosier dogwood | Cornus sericea |
| rose | Rosa spp. |
| sagebrush | Artemisia spp. |
| snowberry | Symphoricarpos spp. |
| vine maple | Acer circinatum |
| Forbs | |
| black mustard | Brassica nigra |
| caterpillar phacelia | Phacelia cicutaria |
| Indian rhubarb | Darmera peltata |
| hollowleaf annual lupine | Lupinus succulentus |
| horehound | Marrubium vulgare |
| stinging nettle | Urtica dioica |
| Graminoids | |
| bluebunch wheatgrass | Pseudoroegneria spicata |
| giant reed | Arundo donax |
| Idaho fescue | Festuca idahoensis |
| ripgut brome | Bromus diandrus |
| Ferns and fern allies | |
| field horsetail | Equisetum arvense |
| giant chainfern | Woodwardia fimbriata |
| scouringrush horsetail | Equisetum hyemale |
| western bracken fern | Pteridium aquilinum |
1. Agee, James K. 1990. The historical role of fire in Pacific Northwest forests. In: Walstad, John D.; Radosevich, Steven R.; Sandberg, David V., eds. Natural and prescribed fire in Pacific Northwest forests. Corvallis, OR: Oregon State University Press: 25-38. [46954]
2. Allen-Diaz, Barbara; Jackson, Randall D.; Phillips, Catherine. 2001. Spring-fed plant communities of California's East Bay Hills oak woodlands. Madrono. 48(2): 98-111. [40591]
3. American Forests. 2014. White alder: Alnus rhombifolia. In: National register of big trees, [Online]. Washington, DC: American Forests (Producer). Available: http://www.americanforests.org/resources/bigtrees/ [2014, August 8]. [88185]
4. Anderson, R. Scott. 1994. Paleohistory of a giant sequoia grove: the record from Log Meadow, Sequoia National Park. In: Aune, Philip S., technical coordinator. Proceedings of the symposium on giant sequoias: their place in the ecosystem and society; 1992 June 23-25; Visalia, CA. Gen. Tech. Rep. PSW-GTR-151. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 49-55. [24749]
5. Arno, Stephen F. 2000. Fire in western forest ecosystems. In: Brown, James K.; Smith, Jane Kapler, eds. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42-vol. 2. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 97-120. [36984]
6. Arno, Stephen F.; Hammerly, Ramona P. 1977. Northwest trees. Seattle, WA: The Mountaineers. 222 p. [4208]
7. Bagne, Karen E.; Purcell, Kathryn L.; Rotenberry, John T. 2008. Prescribed fire, snag population dynamics, and avian nest site selection. Forest Ecology and Management. 255(1): 99-105. [69698]
8. Baldwin, Bruce G.; Goldman, Douglas H.; Keil, David J.; Patterson, Robert; Rosatti, Thomas J.; Wilken, Dieter H., eds. 2012. The Jepson manual. Vascular plants of California, second edition. Berkeley, CA: University of California Press. 1568 p. [86254]
9. Barbour, Michael G. 1987. Community ecology and distribution of California hardwood forests and woodlands. In: Plumb, Timothy R.; Pillsbury, Norman H., technical coordinators. Proceedings of the symposium on multiple-use management of California's hardwood resources; 1986 November 12-14; San Luis Obispo, CA. Gen. Tech. Rep. PSW-100. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 18-25. [5356]
10. Barrett, S.; Havlina, D.; Jones, J.; Hann, W.; Frame, C.; Hamilton, D.; Schon, K.; Demeo, T.; Hutter, L.; Menakis, J. 2010. Interagency Fire Regime Condition Class Guidebook. Version 3.0, [Online]. In: Interagency Fire Regime Condition Class (FRCC). U.S. Department of Agriculture, Forest Service; U.S. Department of the Interior; The Nature Conservancy (Producers). Available: http://www.frcc.gov/ [2013, May 13]. [85876]
11. Barro, Sue. 1989. [Email to Bill Fischer]. April 29. Regarding riparian vegetation after fire - a case study. Riverside, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station, Forest Fire Laboratory. On file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [6815]
12. Barro, Susan C.; Wohlgemuth, Peter M.; Campbell, Allan G. 1989. Post-fire interactions between riparian vegetation and channel morphology and the implications for stream channel rehabilitation choices. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 51-53. [21778]
13. Bauer, Harry L. 1936. Moisture relations in the chaparral of the Santa Monica Mountains, California. Ecological Monographs. 6(3): 409-454. [10528]
14. Beche, Leah A.; Stephens, Scott L.; Resh, Vincent H. 2005. Effects of prescribed fire on a Sierra Nevada (California, USA) stream and its riparian zone. Forest Ecology and Management. 218(1-3): 37-59. [55791]
15. Bendix, Jacob. 1994. Among-site variation in riparian vegetation of the southern California Transverse Range. The American Midland Naturalist. 132(1): 136-151. [83377]
16. Bendix, Jacob. 2014. [Personal communication to Janet Fryer]. 18 September. Regarding fire behavior and postfire responses of Fremont cottonwood, bigberry manzanita, and white alder to the 2012 Wolf Creek Wildfire. On file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [88298]
17. Bendix, Jacob; Cowell, C. Mark. 2010. Impacts of wildfire on the composition and structure of riparian forests in southern California. Ecosystems. 13(1): 99-107. [81845]
18. Bendix, Jacob; Cowell, C. Mark. 2013. Disturbance and riparian tree establishment in the Sespe Wilderness, California, USA. Physical Geography. 34(2): 149-158. [88318]
19. Bolsinger, Charles L. 1988. The hardwoods of California's timberlands, woodlands, and savannas. Resour. Bull. PNW-RB-148. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 148 p. [5291]
20. Borchert, Mark; Segotta, Daniel; Purser, Michael D. 1988. Coast redwood ecological types of southern Monterey County, California. Gen. Tech. Rep. PSW-107. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 27 p. [10225]
21. Bormann, Bernard T. 1988. A masterful scheme: Symbiotic nitrogen-fixing plants of the Pacific Northwest. University of Washington Arboretum Bulletin. 51(2): 10-14. [6796]
22. Bowcutt, Frederica S. 1999. A floristic study of Sugarloaf Ridge State Park, Sonoma County, California. Aliso. 18(1): 19-34. [40636]
23. Bowler, Peter A. 1990. Riparian woodland: an endangered habitat in southern California. In: Schoenherr, Allan A., ed. Endangered plant communities of southern California: Proceedings, 15th annual symposium; 1989 October 28; Fullerton, CA. Special Publication No. 3. Claremont, CA: Southern California Botanists: 80-97. [21321]
24. Brode, John M.; Bury, R. Bruce. 1984. The importance of riparian systems to amphibians and reptiles. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of the conference. 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 30-36. [5823]
25. Brooks, Matthew L.; Minnich, Richard A. 2006. Southeastern deserts bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 391-414. [65559]
26. Brothers, Timothy S. 1985. Riparian species distributions in relation to stream dynamics, San Gabriel River, California. Los Angeles, CA: University of California. 120 p. Dissertation. [8971]
27. Brown, David E. 1979. Southwestern wetlands - their classification and characteristics. In: Johnson, R. Roy; McCormick, J. Frank, technical coordinators. Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium; 1978 December 11-13; Callaway Gardens, GA. Gen. Tech. Rep. WO-12. Washington, DC: U.S. Department of Agriculture, Forest Service: 269-282. [4366]
28. Byrne, Roger; Michaelsen, Joel; Soutar, Andrew. 1977. Fossil charcoal as a measure of wildfire frequency in southern California: a preliminary analysis. In: Mooney, Harold A.; Conrad, C. Eugene, technical coordinators. Proceedings of the symposium on the environmental consequences of fire and fuel management in Mediterranean ecosystems; 1977 August 1-5; Palo Alto, CA. Gen. Tech. Rep. WO-3. Washington, DC: U.S. Department of Agriculture, Forest Service: 361-367. [4848]
29. Carlquist, Sherwin. 1989. Adaptive wood anatomy of chaparral shrubs. In: Keeley, Sterling C., ed. The California chaparral: Paradigms reexamined. No. 34: Science Series. Los Angeles, CA: Natural History Museum of Los Angeles County: 25-35. [84194]
30. Charron, I.; Johnson, E. A. 2006. The importance of fires and floods on tree ages along mountainous gravel-bed streams. Ecological Applications. 16(5): 1757-1770. [64733]
31. Cheng, Sheauchi, ed. 2004. Forest Service Research Natural Areas in California. Gen. Tech. Rep. PSW-GTR-188. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station. 338 p. [88290]
32. Conard, Susan G.; MacDonald, Rod L.; Holland, Robert F. 1980. Riparian vegetation and flora of the Sacramento Valley. In: Sands, Anne, editor. Riparian forests in California: Their ecology and conservation: Symposium proceedings; 1977 May 14; Davis, CA. Davis, CA: University of California, Division of Agricultural Sciences: 47-55. [5285]
33. Dains, Virginia I. 1989. Water relations of white alder. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 375-380. [13769]
34. Dale, Nancy. 1986. Flowering plants: The Santa Monica Mountains, coastal and chaparral regions of southern California. Santa Barbara, CA: Capra Press. 239 p. In cooperation with: The California Native Plant Society. [7605]
35. Davis, Frank W.; Keller, Edward A.; Parikh, Anuja; Florsheim, Joan. 1989. Recovery of the chaparral riparian zone after wildfire. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 194-203. [13883]
36. DeBano, Leonard F.; Neary, Daniel G. 1996. Effects of fire on riparian systems. In: Ffolliott, Peter F.; DeBano, Leonard F.; Baker, Malchus B., Jr.; Gottfried, Gerald J.; Solis-Garza, Gilberto; Edminster, Carleton B.; Neary, Daniel G.; Allen, Larry S.; Hamre, R. H., tech. coords. Effects of fire on Madrean Province ecosystems: a symposium proceedings; 1996 March 11-15; Tucson, AZ. Gen. Tech. Rep. RM-GTR-289. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 69-76. [28064]
37. Dirr, Michael A.; Heuser, Charles W., Jr. 1987. The reference manual of woody plant propagation: From seed to tissue culture. Athens, GA: Varsity Press. 239 p. [16999]
38. Fites, Jo Ann. 1993. Ecological guide to mixed conifer plant associations--northern Sierra Nevada and southern Cascades: Lassen, Plumas, Tahoe, and El Dorado National Forests. R5-ECOL-TP-001. Vallejo, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Region. 120 p. [82332]
39. Flora of North America Editorial Committee, eds. 2014. Flora of North America north of Mexico, [Online]. Flora of North America Association (Producer). Available: http://www.efloras.org/flora_page.aspx?flora_id=1. [36990]
40. Franklin, Jerry F.; Dyrness, C. T. 1973. Natural vegetation of Oregon and Washington. Gen. Tech. Rep. PNW-8. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 417 p. [961]
41. Frost, Evan J.; Sweeney, Rob. 2000. Fire regimes, fire history and forest conditions in the Klamath-Siskiyou region: an overview and synthesis of knowledge, [Online]. In: Klamath Siskiyou Wildland Center--Fire ecology and policy. Ashland, OR: Wildwood Environmental Consulting (Producer). Available: http://kswild.org/fire/fire_report.pdf [2011, November 15]. [69111]
42. Furlow, John J. 1979. The systematics of the American species of Alnus (Betulaceae)--I. Rhodora. 81(825): 1-121. [6195]
43. Gaines, David A. 1980. The valley riparian forests of California: their importance to bird populations. In: Sands, Anne, editor. Riparian forests in California: Their ecology and conservation: Symposium proceedings; 1977 May 14; Davis, CA. Davis, CA: University of California, Division of Agricultural Sciences: 57-85. [5286]
44. Germplasm Resources Information Network. 2014. National plant germplasm system, [Online]. Beltsville, MD: U.S. Department of Agriculture, Agricultural Research Service, Germplasm Resources Information Network (Producer). Available: http://www.ars-grin.gov/npgs/index.html [69523]
45. Going, Barbara M.; Dudley, Tom L. 2008. Invasive riparian plant litter alters aquatic insect growth. Biological Invasions. 10(7): 1041-1051. [72888]
46. Goldner, Bernard H. 1984. Riparian restoration efforts associated with structurally modified flood control channels. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of the conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 445-451. [5852]
47. Govaerts, Rafael; Frodin, David G. 1998. World checklist and bibliography of Fagales (Betulaceae, Corylaceae, Fagaceae and Ticodendraceae). Kew, UK: The Royal Botanic Gardens. 497 p. [60947]
48. Graves, George W. 1932. Ecological relationships of Pinus sabiniana. Botanical Gazette. 94(1): 106-133. [63160]
49. Gray, M. Violet; Greaves, James M. 1984. Riparian forest as habitat for the least Bell's vireo. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 605-611. [5862]
50. Griffin, James R. 1974. Notes on environment, vegetation and flora: Hastings Natural History Reservation. Carmel Valley, CA: University of California, Hastings Natural History Reservation. Unpublished paper on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 90 p. [10531]
51. Griffin, James R.; Critchfield, William B. 1972. The distribution of forest trees in California. Res. Pap. PSW-82. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 118 p. [1041]
52. Halofsky, Jessica E.; Hibbs, David E. 2009. Controls on early post-fire woody plant colonization in riparian areas. Forest Ecology and Management. 258(7): 1350-1358. [77249]
53. Harrington, Constance A.; Brodie, Leslie Chandler; DeBell, Dean S.; Schopmeyer, C. S. 2008. Alnus P. Mill.: alder. In: Bonner, Franklin T.; Karrfalt, Robert P., eds. Woody plant seed manual. Agric. Handbook No. 727. Washington, DC: U.S. Department of Agriculture, Forest Service: 232-242. [79017]
54. Harris, Roger D. 1983. Decay characteristics of pileated woodpecker nest trees. In: Davis, Jerry W.; Goodwin, Gregory A.; Ockenfeis, Richard A., technical coordinators. Snag habitat management: proceedings of the symposium; 1983 June 7-9; Flagstaff, AZ. Gen. Tech. Rep. RM-99. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 125-129. [17826]
55. Hehnke, Merlin; Stone, Charles P. 1979. Value of riparian vegetation to avian populations along the Sacramento River System. In: Johnson, R. Roy; McCormick, J. Frank, technical coordinators. Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium; 1978 December 11-13; Callaway Gardens, GA. General Technical Report WO-12. Washington, DC: U.S. Department of Agriculture, Forest Service: 228-235. [4363]
56. Hitchcock, C. Leo; Cronquist, Arthur. 1964. Vascular plants of the Pacific Northwest. Part 2: Salicaceae to Saxifragaceae. Seattle, WA: University of Washington Press. 597 p. [1166]
57. Holland, Robert F. 1986. Preliminary descriptions of the terrestrial natural communities of California. Sacramento, CA: California Department of Fish and Game. 156 p. [12756]
58. Holmgren, Noel H.; Holmgren, Patricia K.; Reveal, James L. 2012. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Vol. 2, Part A: Subclass Magnoliidae-Caryophyllidae. New York: The New York Botanical Garden Press. 731 p. [88110]
59. Holstein, Glen. 1984. California riparian forests: deciduous islands in an evergreen sea. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 2-22. [5830]
60. Horton, Jerome S.; Wright, John T. 1944. The wood rat as an ecological factor in southern California watersheds. Ecology. 25(3): 341-351. [10682]
61. Jahn, Laurence R. 1979. Values of riparian habitats to natural systems. In: Johnson, R. Roy; McCormick, J. Frank, technical coordinators. Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium; 1978 December 11-13; Callaway Gardens, GA. Gen. Tech. Rep. WO-12. Washington, DC: U.S. Department of Agriculture, Forest Service: 157-160. [88300]
62. Jankovsky-Jones, Mabel; Rust, Steven K.; Moseley, Robert K. 1999. Riparian reference areas in Idaho: a catalog of plant associations and conservation sites. Gen. Tech. Rep. RMRS-GTR-20. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 141 p. [29900]
63. Johnson, Frederic D. 1968. Taxonomy and distribution of northwestern alders. In: Trappe, J. M.; Franklin, J. F.; Tarrant, R. F.; Hansen, G. M., ed. Biology of alder; 1967 April 14-15; Pullman, WA. Portland, OR: U. S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: 9-22. [6187]
64. Kartesz, J. T.; The Biota of North America Program (BONAP). 2014. North American plant atlas, [Online]. Chapel Hill, NC: The Biota of North America Program (Producer). Available: http://bonap.org/. [Maps generated from Kartesz, J. T. 2010. Floristic synthesis of North America, Version 1.0. Biota of North America Program (BONAP). [In press]. [84789]
65. Kartesz, John Thomas. 1988. A flora of Nevada. Reno, NV: University of Nevada. 1729 p. Dissertation. [In 2 volumes]. [42426]
66. Keeley, Jon E. 2006. South Coast bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 350-390. [65557]
67. Keeley, Jon E.; Zedler, Paul H. 2009. Large, high-intensity fire events in southern California shrublands: debunking the fine-grain age patch model. Ecological Applications. 19(1): 69-94. [74209]
68. Kobziar, Leda N.; McBride, Joe R. 2006. Wildfire burn patterns and riparian vegetation response along two northern Sierra Nevada streams. Forest Ecology and Management. 222(1-3): 254-265. [79123]
69. LANDFIRE Rapid Assessment. 2005. Reference condition modeling manual (Version 2.1). Cooperative Agreement 04-CA-11132543-189. Boulder, CO: The Nature Conservancy; U.S. Department of Agriculture, Forest Service; U.S. Department of the Interior. 72 p. On file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. [66741]
70. LANDFIRE Rapid Assessment. 2007. Rapid assessment potential natural vegetation groups (PNVGs): associated vegetation descriptions and geographic distributions. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Lab; U.S. Geological Survey; Arlington, VA: The Nature Conservancy. 84 p. [66533]
71. Laymon, Stephen A. 1984. Riparian bird community structure and dynamics: Dog Island, Red Bluff, California. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 587-597. [5860]
72. Lisle, Thomas E. 1989. Channel-dynamic control on the establishment of riparian trees after large floods in northwestern California. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 9-13. [13508]
73. Little, Elbert L., Jr. 1976. Atlas of United States trees. Volume 3. Minor western hardwoods. Misc. Publ. 1314. Washington, DC: U.S. Department of Agriculture, Forest Service. 13 p. [+ 290 maps]. [10430]
74. Little, Elbert L., Jr. 1979. Checklist of United States trees (native and naturalized). Agric. Handb. 541. Washington, DC: U.S. Department of Agriculture, Forest Service. 375 p. [2952]
75. McBride, Joe R.; Strahan, Jan. 1984. Establishment and survival of woody riparian species on gravel bars of an intermittent stream. The American Midland Naturalist. 112(2): 235-245. [9675]
76. McBride, Joe R.; Strahan, Jan. 1984. Fluvial processes and woodland succession along Dry Creek, Sonoma County, California. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 110-119. [5832]
77. McCain, Cindy; Christy, John A. 2005. Field guide to riparian plant communities in northwestern Oregon. Tech. Pap. R6-NR-ECOL-TP-01-05. [Portland, OR]: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 357 p. [63114]
78. McDonald, Philip M.; Littrell, Edward E. 1976. The bigcone Douglas-fir--canyon live oak community in southern California. Madrono. 23(6): 310-320. [10662]
79. McKenney, Daniel W.; Pedlar, John H.; Lawrence, Kevin; Campbell, Kathy; Hutchinson, Michael F. 2007. Potential impacts of climate change on the distribution of North American trees. BioScience. 57(11): 939-948. [70374]
80. Medin, Dean E.; Clary, Warren P. 1991. Small mammals of a beaver pond ecosystem and adjacent riparian habitat in Idaho. Res. Pap. INT-445. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 4 p. [16003]
81. Messier, Michael S.; Shatford, Jeff P. A.; Hibbs, David E. 2012. Fire exclusion effects on riparian forest dynamics in southwestern Oregon. Forest Ecology and Management. 264: 60-71. [84153]
82. Miller, Thomas B.; Johnson, Frederic D. 1986. Sampling and data analyses of narrow, variable-width gallery forests over environmental gradients. Tropical Ecology. 27: 132-142. [12310]
83. Minnich, Richard A. 1976. Vegetation of the San Bernardino Mountains. In: Latting, June, ed. Symposium proceedings: Plant communities of southern California; 1974 May 4; Fullerton, CA. Special Publication No. 2. Berkeley, CA: California Native Plant Society: 99-124. [4232]
84. Minnich, Richard A.; Franco-Vizcaino, Ernesto. 1997. Mediterranean vegetation of northern Baja California. Fremontia. 25(3): 3-12. [40196]
85. Minnich, Richard A.; Franco-Vizcaino, Ernesto. 2009. A probabilistic view of chaparral and forest fire regimes in southern California and northern Baja California. In: Bytnerowicz, Andrzej; Arbaugh, Michael J.; Riebau, Allen R.; Andersen, Christian, eds. Wildland fires and air pollution. Developments in Environmental Science: Volume 8. Amsterdam, The Netherlands: Elsevier B. V: 339-363. [81235]
86. Moseley, Robert K. 1998. Riparian and wetland community inventory of 14 reference areas in southwestern Idaho. Technical Bulletin No. 98-5. Boise, Idaho: U.S. Department of the Interior, Bureau of Land Management, Boise State Office. 52 p. [75569]
87. Munz, Philip A.; Keck, David D. 1973. A California flora and supplement. Berkeley, CA: University of California Press. 1905 p. [6155]
88. Nachlinger, Janet L.; Smith, Stanley D.; Risser, Roland J. 1989. Riparian plant water relations along the North Fork Kings River, California. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 366-369. [88200]
89. NatureServe. 2014. NatureServe Explorer: An online encyclopedia of life, [Online]. Version 7.1. Arlington, VA: NatureServe (Producer). Available http://www.natureserve.org/explorer. [69873]
90. Neary, D. G.; Comeford, N. B.; Swift, L. W., Jr. 1993. Land and riparian interactions with sediment in the southern United States. In: Proceedings of a technical workshop on sediments; 1992 February 3-7; Corvallis, OR. Washington, DC: Terrene Institute: 51-60. [88202]
91. Nelson, Duane; Lux, Linda; Thompson, Steve; Brochert, Mark; Gifford, Dan; Brandow, Clay; Levin, Maxine; Balen, Barbara; Barbour, Michael; Pronos, John; Wenz, John; Stover, Paul; Husari, Sue. 1994. Foothill riparian: Ecological Support Team report. In: Toth, Edward; Laboa, Jane; Nelson, Duane; Hermit, Raymond; Andrews, Richard S., eds. Ecological Support Team workshop proceedings for the California spotted owl Environmental Impact Statement; 1993 August 16; Sacramento, CA. San Francisco, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Region. 19 p. [68114]
92. O'Geen, Anthony T.; Dahlgren, Randy A.; Sanchez-Mata, Daniel. 2007. California soils and examples of ultramafic vegetation. In: Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. Terrestrial vegetation of California. 3rd ed. Berkeley, CA: University of California Press: 71-106. [82692]
93. Oakley, Brian; North, Malcolm; Franklin, Jerry F.; Hedlund, Brian P.; Staley, James T. 2004. Diversity and distribution of Frankia strains symbiotic with Ceanothus in California. Applied and Environmental Microbiology. 70(11): 6444-6452. [60444]
94. Ohmann, Janet L.; Gregory, Matthew J.; Spies, Thomas A. 2007. Influence of environment, disturbance, and ownership on forest vegetation of coastal Oregon. Ecological Applications. 17(1): 18-33. [83690]
95. Opperman, Jeff J.; Merenlender, Adina M. 2000. Deer herbivory as an ecological constraint to restoration of degraded riparian corridors. Restoration Ecology. 8(1): 41-47. [39021]
96. Paysen, Timothy E.; Ansley, R. James; Brown, James K.; Gottfried, Gerald J.; Haase, Sally M.; Harrington, Michael G.; Narog, Marcia G.; Sackett, Stephen S.; Wilson, Ruth C. 2000. Fire in western shrubland, woodland, and grassland ecosystems. In: Brown, James K.; Smith, Jane Kapler, eds. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42-vol. 2. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 121-159. [36978]
97. Perala, C.; Hoover, D. A. 1990. Hand-removal of exotics and planting of natives key to restoration of riparian forest understory. Restoration and Management Notes. 8(2): 118. [13791]
98. Pokharel, Anita; Mirza, Babur S.; Dawson, Jeffrey O.; Hahn, Dittmar. 2011. Frankia populations in soil and root nodules of sympatrically grown Alnus taxa. Microbial Ecology. 61(1): 92-100. [88122]
99. Pratt, R. B.; Black, R. A. 2006. Do invasive trees have a hydraulic advantage over native trees? Biological Invasions. 8(6): 1331-1341. [71478]
100. Purcell, Kathryn, L.; Drynan, Douglas A. 2008. Use of hardwoods by birds nesting in ponderosa pine forests. In: Merenlender, Adina; McCreary, Douglas; Purcell, Kathryn L., tech. eds. Proceedings of the 6th symposium on oak woodlands: today's challenges, tomorrow's opportunities; 2006 October 9-12; Rohnert Park, CA. Gen. Tech. Rep. PSW-GTR-217. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 417-431. [80945]
101. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press. 632 p. [2843]
102. Riegel, Gregg M.; Miller, Richard F.; Skinner, Carl N.; Smith, Sydney E. 2006. Northeastern Plateaus bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 225-263. [65541]
103. Ringold, Paul L.; Magee, Teresa K.; Peck, David V. 2008. Twelve invasive plant taxa in US western riparian ecosystems. Journal of the North American Benthological Society. 27(4): 949-966. [73064]
104. Roberts, Warren G.; Howe, J. Greg; Major, Jack. 1980. A survey of riparian forest flora and fauna in California. In: Sands, Anne, ed. Riparian forests in California: Their ecology and conservation: Symposium proceedings; 1977 May 14; Davis, CA. Institute of Ecology Publication No. 15. Davis, CA: University of California, Division of Agricultural Sciences: 3-19. [5271]
105. Robinson, Cyril S. 1937. Plants eaten by California mule deer on the Los Padres National Forest. Journal of Forestry. 35(3): 285-292. [51853]
106. Rundel, Philip W. 1971. Community structure and stability in the giant sequoia groves of the Sierra Nevada, California. The American Midland Naturalist. 85(2): 478-492. [10504]
107. Rundel, Philip Wilson. 1969. The distribution and ecology of the giant sequoia ecosystem in the Sierra Nevada, California. Durham, NC: Duke University. 205 p. Dissertation. [37436]
108. Russell, William H.; McBride, Joe R. 2001. The relative importance of fire and watercourse proximity in determining stand composition in mixed conifer riparian forests. Forest Ecology and Management. 150(3): 259-265. [39300]
109. Safford, Hugh Deforest. 1995. Woody vegetation and succession in the Garin Woods, Hayward Hills, Alameda County, California. Madrono. 42(4): 470-489. [40868]
110. Sampson, Arthur W.; Jespersen, Beryl S. 1963. California range brushlands and browse plants. Berkeley, CA: University of California, Division of Agricultural Sciences; California Agricultural Experiment Station, Extension Service. 162 p. [3240]
111. Sands, Anne. 1979. Public involvement in riparian habitat protection: A California case history. In: Johnson, R. Roy; McCormick, J. Frank, technical coordinators. Strategies for protection and management of floodplain wetlands and other riparian ecosystems: Proceedings of the symposium; 1978 December 11-13; Callaway Gardens, GA. General Technical Report WO-12. Washington, DC: U.S. Department of Agriculture, Forest Service: 216-227. [4362]
112. Sarr, D. A.; Hibbs, D. E. 2007. Woody riparian plant distributions in western Oregon, USA: comparing landscape and local scale factors. Plant Ecology. 190(2): 291-311. [84241]
113. Sarr, Daniel A.; Hibbs, David E.; Shatford, Jeffrey P. A.; Momsen, Rick. 2011. Influences of life history, environmental gradients, and disturbance on riparian tree regeneration in Western Oregon. Forest Ecology and Management. 261(7): 1241-1253. [86637]
114. Sauer, Jonathan D. 1977. Fire history, environmental patterns, and species patterns in Santa Monica Mountain chaparral. In: Mooney, Harold A.; Conrad, C. Eugene, technical coordinators. Proceedings of the symposium of the environmental consequences of fire and fuel management in Mediterranean ecosystems; 1977 August 1-5; Palo Alto, CA. Gen. Tech. Rep. WO-3. Washington, DC: U.S. Department of Agriculture, Forest Service: 383-386. [4866]
115. Schopmeyer, C. S. 1974. Alnus B. Ehrh. alder. In: Schopmeyer, C. S., technical coordinator. Seeds of woody plants in the United States. Agric. Handb. 450. Washington, DC: U.S. Department of Agriculture, Forest Service: 206-211. [7460]
116. Scott, Joe H.; Reinhardt, Elizabeth D. 2001. Assessing crown fire potential by linking models of surface and crown fire behavior. Res. Pap. RMRS-RP-29. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 59 p. [88259]
117. Shaffer, Kevin E.; Laudenslayer, William F., Jr. 2006. Fire and animal interactions. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 118-144. [65536]
118. Shanfield, Allan N. 1984. Alder, cottonwood, and sycamore distribution and regeneration along the Nacimiento River, California. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 196-202. [5834]
119. Skinner, Carl N. 2002. Fire history in riparian reserves of the Klamath Mountains. In: Sugihara, Neil G.; Morales, Maria; Morales, Tony, eds. Fire in California ecosystems: integrating ecology, prevention and management: Proceedings of the symposium; 1997 November 17-20; San Diego, CA. Misc. Pub. No. 1. [Berkeley, CA]: Association for Fire Ecology: 164-169. [46203]
120. Skinner, Carl N.; Taylor, Alan H.; Agee, James K. 2006. Klamath Mountains bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 170-194. [65539]
121. Spina, Anthony P.; Tormey, Daniel R. 2000. Postfire sediment deposition in geographically restricted steelhead habitat. American Journal of Fisheries Management. 20(2): 562-569. [40230]
122. Stanley, John T.; Silva, L. R.; Appleton, H. C.; Marangio, M. S.; Lapaz, W. J.; Goldner, B. H. 1989. Coyote Creek (Santa Clara County) pilot revegetation project. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 447-454. [88201]
123. Stewart, Glenn R.; Jennings, Mark R.; Goodman, Robert H., Jr. 2005. Sensitive species of snakes, frogs, and salamanders in southern California conifer forest areas: status and management. In: Kus, Barbara E.; Beyers, Jan L., tech. coords. Planning for biodiversity: Bringing research and management together: Proceedings of a symposium for the South Coast ecoregion; 2000 February 29 - March 2; Pomona, CA. Gen. Tech. Rep. PSW-GTR-195. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 165-197. [64510]
124. Stickney, Peter F. 1989. Seral origin of species comprising secondary plant succession in northern Rocky Mountain forests. FEIS workshop: Postfire regeneration. Unpublished draft on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 10 p. [20090]
125. Stuart, John D.; Worley, Tom; Buell, Ann C. 1996. Plant associations of Castle Crags State Park, Shasta County, California. Madrono. 43(2): 273-291. [64661]
126. Sudworth, George B. 1900. Stanislaus and Lake Tahoe Forest Reserves, California, and adjacent territory. In: Annual reports of the Department of Interior, 21st annual report of the U.S. Geological Survey. Part 5. Washington, DC: Government Printing Office: 505-561. [65699]
127. Svejcar, Tony. 1997. Riparian zones: 2) History and human impacts. Rangelands. 19(4): 8-12. [88332]
128. Swanson, John; Johnson, Robert C.; Merrifield, Dave; Dennestan, Alan. 1982. Fire management plan: Lassen fire management planning area: Lassen Volcanic National Park-Caribou Wilderness Unit: Implementation plan. Mineral, CA: U.S. Department of the Interior, National Park Service, Lassen Volcanic National Park; Susanville, CA: U.S. Department of Agriculture, Forest Service, Lassen National Forest. 66 p. [21407]
129. Sweet, Muriel. 1962. Common edible and useful plants of the West. Healdsburg, CA: Naturegraph Company. 64 p. [54095]
130. Thode, Andrea E.; van Wagtendonk, Jan W.; Miller, Jay D.; Quinn, James F. 2011. Quantifying the fire regime distributions for severity in Yosemite National Park, California, USA. International Journal of Wildland Fire. 20(2): 223-239. [84640]
131. Thompson, Ralph L. 2001. Botanical survey of Myrtle Island Research Natural Area, Oregon. Gen. Tech. Rep. PNW-GTR-507. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 27 p. [43785]
132. Tietje, William D.; Barrett, Reginald H.; Kleinfelter, Eric B.; Carre, Brett T. 1991. Wildlife diversity in valley-foothill riparian habitat: North central vs. central coast California. In: Standiford, Richard B., technical coordinator. Proceedings of the symposium on oak woodlands and hardwood rangeland management; 1990 October 31 - November 2; Davis, CA. Gen. Tech. Rep. PSW-126. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 120-125. [18898]
133. Timbrook, Jan. 1990. Ethnobotany of Chumash Indians, California, based on collections by John P. Harrington. Economic Botany. 44(2): 236-253. [13777]
134. Trush, William J.; Connor, Edward C.; Knight, Allen W. 1989. Alder establishment and channel dynamics in a tributary of the South Fork Eel River, Mendocino County, California. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 14-21. [13509]
135. Tuhy, Joel S.; Jensen, Sherman. 1982. Riparian classification for the Upper Salmon/Middle Fork Salmon River drainages, Idaho. Final report. Smithfield, UT: White Horse Associates. Unpublished report on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station Fire Sciences Laboratory, Missoula, MT. 153 p. [8380]
136. U.S. Department of Agriculture, Forest Service. 1937. Range plant handbook. Washington, DC: U.S. Department of Agriculture, Forest Service. 532 p. [2387]
137. U.S. Department of Agriculture, Natural Resources Conservation Service. 2014. PLANTS Database, [Online]. Available: https://plants.usda.gov /. [34262]
138. U.S. Department of the Interior, Fish and Wildlife Service, Sacramento Fish and Wildlife Office. 1999. Conservation guidelines for the valley elderberry longhorn beetle. Sacramento, CA: U.S. Department of the Interior, Fish and Wildlife Service, Sacramento Fish and Wildlife Office. Available: http://www.fws.gov/sfbaydelta/documents/velb_conservation.pdf [2013, January 31]. [86474]
139. U.S. Fish and Wildlife Service, Region 2. 2002. Final recovery plan: Southwestern willow flycatcher (Empidonax traillii extimus), [Online]. Albuquerque, NM: Southwestern Willow Flycatcher Recovery Team (Producer). Available: http://www.treesearch.fs.usda.gov/pubs/35557 [2014, October 16]. [44503]
140. Van Cleve, David H.; Comrack, Lyann A.; Wier, Harold A. 1989. Coyote Creek (San Diego County) managment and restoration at Anza-Borrego Desert State Park. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 149-153. [88196]
141. Verkaik, Iraima; Rieradevall, Maria; Cooper, Scott D.; Melack, John M; Dudley, Tom L.; Prat, Narcis. 2013. Fire as a disturbance in mediterranean climate streams. Hydrobiologia. 719(1): 353-382. [88326]
142. Verner, Jared; McKelvey, Kevin S.; Noon, Barry R; Gutierrez, R. J.; Gould, Gordon I., Jr.; Beck, Thomas W. 1992. Assessment of the current status of the California spotted owl, with recommendations for management. In: Verner, Jared; McKelvey, Kevin S.; Noon, Barry R.; Gutierrez, R. J.; Gould, Gordon I., Jr.; Beck, Thomas W., tech. coords. The California spotted owl: a technical assessment of its current status. Gen. Tech. Rep. PSW-GTR-133. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 3-26. [28195]
143. Villar, R.; Merino, J. 2001. Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems. New Phytologist. 151(1): 213-226. [80064]
144. Vogl, Richard J. 1976. An introduction to the plant communities of the Santa Ana and San Jacinto Mountains. In: Latting, June, ed. Symposium proceedings: plant communities of southern California; 1974 May 4; Fullerton, CA. Special Publication No. 2. Berkeley, CA: California Native Plant Society: 77-98. [4230]
145. Walters, M. Alice; Teskey, Robert O.; Hinckley, Thomas M. 1980. Impact of water level changes on woody riparian and wetland communities. Volume 7: Mediterranean region, western arid and semi-arid region. FWS/OBS-78/93. Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service, Biological Services Program. 84 p. [52899]
146. Walters, M. Alice; Teskey, Robert O.; Hinckley, Thomas M. 1980. Impact of water level changes on woody riparian and wetland communities. Volume 8. Pacific Northwest and Rocky Mountain regions. Final report FWS/OBS 78/94. Kearneysville, WV: U.S. Department of Interior, Fish and Wildlife Service, Biological Services Program. 47 p. [85052]
147. Warner, Richard E. 1984. Structural, floristic, and condition inventory of Central Valley riparian systems. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 356-374. [5840]
148. Weaver, J. E. 1917. A study of the vegetation of southeastern Washington and adjacent Idaho. Nebraska University Studies. 17(1): 1-133. [7153]
149. Whitlow, Thomas H.; Harris, Richard W.; Leiser, Andrew T. 1984. Experimenting with levee vegetation: some unexpected findings. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 558-565. [5858]
150. Whittaker, R. H. 1953. A consideration of climax theory: the climax as a population and pattern. Ecological Monographs. 23(1): 41-78. [64492]
151. Whittaker, R. H. 1960. Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monographs. 30(3): 279-338. [6836]
152. Willoughby, John W.; Davilla, William. 1984. Plant species composition and life form spectra of tidal streambanks and adjacent riparian woodlands along the lower Sacramento River. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 642-651. [5866]
153. Wills, Robin. 2006. Central Valley bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 295-320. [65547]
154. Wilson, James L.; Ayres, Debra R.; Steinmaus, Scott; Baad, Michael. 2009. Vegetation and flora of a biodiversity hotspot: Pine Hill, El Dorado County, California, USA. Madrono. 56(4): 246-278. [80220]
155. Wilson, Pamela L.; Funck, James W.; Avery, Robert B. 2010. Fuelwood characteristics of northwestern conifers and hardwoods. PNW-GTR-810 [Updated]. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 50 p. In cooperation with: Oregon State University. [Updated by Parrent, Daniel J.; Funck, James W.; Reeb, James; Brackley, Allen M]. [81905]
156. Wolfe, Douglas. 1988. Recreating a "natural" riparian environment, or getting the creek out of the culvert. In: Rieger, John P.; Williams, Bradford K., eds. Proceedings of the second native plant revegetation symposium; 1987 April 15-18; San Diego, CA. Madison, WI: University of Wisconsin Arboretum, Society for Ecological Restoration & Management: 193-197. [4114]
157. Young, James A.; Young, Cheryl G. 1992. Seeds of woody plants in North America. [Revised and enlarged edition]. Portland, OR: Dioscorides Press. 407 p. [72640]
158. Zembal, Richard. 1998. Habitat for threatened habitat and endangered species--quarantine areas or control exotic weeds? In: Bell, Carl E., ed. Arundo and saltcedar: the deadly duo: Proceedings of a workshop on combating the threat from Arundo and saltcedar; 1998 June 17; Ontario, CA. Holtville, CA: University of California, Cooperative Extension: 15-20. [47121]