| FEIS Home Page |
 |
|
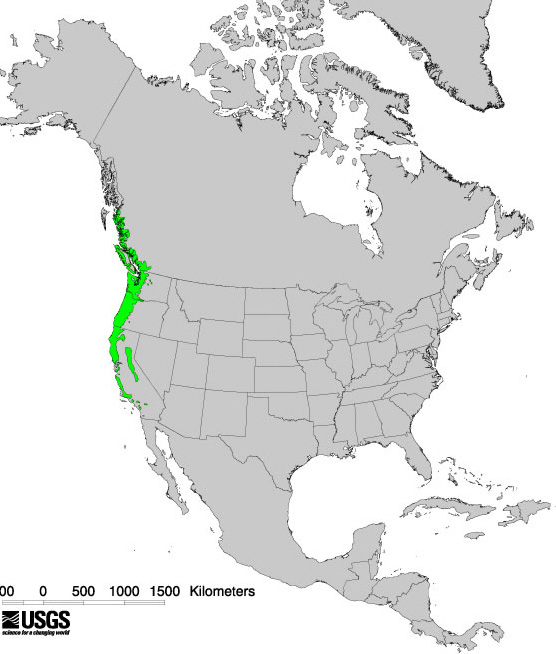 |
Bigleaf maple grows in mountainous regions [138]. It is widespread in the Coast Ranges, the Klamath-Siskiyou Mountains, and the foothills of the Cascade Range and the northern Sierra Nevada [75,81,99,102], obtaining best development in southern Oregon [13]. Some authors place bigleaf maple's distribution as far north as the Alaska panhandle [13,99,138]. Isolated bigleaf maple populations may occur in Idaho [102].
States and provinces (as of 2011 [211]):
United States: CA, OR, WA
Canada: BC
Moisture regime: Bigleaf maple prefers moist to mesic habitats and is described as "moisture demanding" [40]. In the Siskiyou Mountains of southern Oregon and northern California, bigleaf maple frequency increased with increasing soil moisture [219]. Bigleaf maple is most common in upland riparian zones [1] but also grows in low riparian zones and in mesic upland forests of the Pacific Northwest. In riparian habitats, it is often found on upper terraces of 5th- and 6th-order streams [1,219], although it also grows near smaller streams [158,214]. Franklin and Dyrness [63] consider bigleaf maple characteristic of old land surfaces—particularly upper riparian terraces—of the Olympic rain forest. Bigleaf maple is confined to riparian zones, moist canyons [13,81], and other sites with permanent water [151] in central and southern California [20,81,173]. It is occasionally reported on dry sites. It is a minor species, for example, on the Dead Indian Plateau of southwestern Oregon. The site has hot, dry summers, late spring frosts, and is a notoriously difficult site on which to regenerate commercial conifer species [149,193].
Soils: Bigleaf maple often grows in alluvial soils [40,93,105], but it is occasionally found on dry upland soils [17]. Soils supporting bigleaf maple are commonly coarse or gravelly in texture [105,151], although it also grows in fine-textured soils [139]. On the Gulf Islands of British Columbia, bigleaf maple grows on alluvial floodplains, outwash sandy loams, eroded rocky shores, shallow to deep glacial tills, poorly drained marine clays, and wet organic soils [54,55]. In the Pacific Northwest, bigleaf maple thrives in moist gravels [151]. In Sitka spruce-western hemlock (Picea sitchensis-Tsuga heterophylla) forests of Washington and Oregon, shallow, stony soils support bigleaf maple groves [63]. A bigleaf maple/California hazelnut (Corylus cornuta subsp. californica) community on the Suislaw National Forest, Oregon, occurs on loams and silt loams [139]. On the Willamette National Forest, Oregon, bigleaf maple grows in a rock garden community. This community is situated on exposed ridges with patches of shallow soil lodged in rock. The community is highly diverse, with no clear species dominants [97]. Bigleaf maple dominates some talus slopes in the Coast Ranges of Oregon and Washington [63]. On the Tillamook Burn of Oregon, the bigleaf maple/creeping snowberry (Symphoricarpos mollis) vegetation type occurs on steep talus slopes and basalt cliff faces [17,100]. Elevations range from 1,000 to 1,600 feet (300-500 m), and most sites exceed 100% slope. In contrast to bigleaf maple's usual habitat, these sites are xeric [17]. Thirteen years following the last reburn in 1945, groundlayer vegetation in this type was sparse, and soil was still eroding [100].
Quartz diorite is the parent material of soils supporting bigleaf maple in the Siskiyou Mountains [219], and gabbro is the parent material in the Pine Hills of El Dorado County, California [221]. Bigleaf maple grows in ultramafic soils in Port-Orford cedar (Chamaecyparis lawsoniana) riparian forests of the Klamath-Siskiyou region of Oregon and California [156].
Bigleaf maple is not nutrient-demanding [151], although best growth occurs on rich bottomland soils [13].
In the Pacific Northwest, soil pH beneath bigleaf maple averaged 5.5 [200].
Elevation and topography: Bigleaf maple is found from sea level to about 3,000 feet (900 m) in Washington and Oregon [13]. It does not grow much above 1,150 feet (350 m) in British Columbia [83,130,151]. In the Siskiyou Mountains, bigleaf maple frequency was greatest at 1,500 to 2,500 feet (460-760 m) elevation. It decreased with increasing elevation, and bigleaf maple was not found above 5,500 feet (1,700 m) [219]. Bigleaf maple's elevational range extends to about 6,000 feet (1,800 m) in most of California [40,99,138]. It may be found up to 7,000 feet (2,000 m) in southern California [28,151], often growing near montane headwater streams [28].
Topography where bigleaf maple grows is highly variable. Bigleaf maple frequently grows in flat, interior valleys in Washington and Oregon but is also found on steep, rocky slopes and cliffs on the Coast Ranges of southwestern Oregon [151]. In a survey of unmanaged riparian forests in western Oregon, bigleaf maple cover was greatest on floodplains, moderate on stream terraces, and least on low hillslopes on the Coast Ranges. In the Klamath Mountains, it was not found on floodplains, and it had greater cover on terraces than on lower hillslopes [158]. In a study of watersheds near Coos Bay, Oregon, bigleaf maple made up less than 10% of stands near 0-order (headwater) streams. On these sites, its basal area was greatest on downstream slopes, moderate at headwaters, and least in valleys (4.2, 3.5, and 3.1 m²/ha, respectively) [183]. In the Columbia River Gorge, bigleaf maple is associated with south slopes and ravines [223].
Plant communities: Bigleaf maple is common in some conifer, mixed-evergreen, and hardwood communities and in seral brushfields. These communities are often diverse in species composition and structure [162]. Bigleaf maple is associated with Pacific madrone (Arbutus menziesii), Pacific dogwood (Cornus nuttallii), and prince's-pine (Chimaphila) throughout most of its range [151]. Chapters within Barbour and others [21] provide details of other over- and understory species commonly associated with bigleaf maple. In addition to the many vascular plant species with which bigleaf maple is associated [21], many arboreal moss and lichen species use bigleaf maple as a substrate ([105,164], review by [117]). The table below lists plant communities in which bigleaf maple is most common. Among these, bigleaf maple reaches greatest frequency in mixed-evergreen forests of southwestern Oregon and northern California [13,206]. Descriptions of these and other plant communities with bigleaf maple follow. See the Fire Regime Table for further information about these plant communities.
| Plant communities in which bigleaf maple is important | |
| Community type | Area |
| coast Douglas-fir (Pseudotsuga menziesii var. menziesii) | British Columbia, Pacific Northwest, California [4,53,62,63,105,133,151,166,220] |
| grand fir (Abies grandis) | Pacific Northwest [29,133,207] |
| mixed-conifer forests | southern Oregon and California [20] |
| mixed-evergreen forests | Washington, Oregon, and northern California [206] |
| Pacific ponderosa pine (Pinus ponderosa var. ponderosa) | southern Oregon and California [4,143,151,179] |
| Port-Orford cedar (Chamaecyparis lawsoniana) | Oregon, northern California [151,191] |
| redwood | California [151,172,206] |
| Sitka spruce | British Columbia, Pacific Northwest [63,90] |
| western hemlock | British Columbia, Pacific Northwest [62,105,133,150,151,166] |
| white fir (Abies concolor) | southern Oregon and northern California [151] |
| western redcedar (Thuja plicata) | Washington, Oregon [62,105,151,151] |
| black cottonwood (Populus balsamifera subsp. trichocarpa) | Pacific Northwest, California [20,62,93,105,117,151] |
| California bay (Umbellularia californica) | Oregon, California [176] |
| coast live oak (Q. agrifolia) | California [103] |
| Oregon white oak (Quercus garryana) | Oregon, northern California [63,151,176,192] |
| red alder (Alnus rubra) | Pacific Northwest, California [20,93,117,147] |
| white alder (A. rhombifolia) | California [103] |
Conifer communities: Bigleaf maple often occurs in scattered patches within or on the streamside edges of conifer-dominated riparian communities [117]. In surveys near rivers of the Puget Sound area of Washington, bigleaf maple was most important in areas just adjacent to waterways, while western redcedar and Douglas-fir tended to dominate upland riparian zones [41]. Within western hemlock-Sitka spruce forests, bigleaf maple grows mostly as scattered individuals or in groves of large trees (≥30 inches (76 cm) DBH) within the conifer forest matrix. It is not restricted to riparian sites in these mesic forest types [63]. Bigleaf maple is a minor species in Pacific silver fir (Abies amabilis) forests of the Olympic National Forest, Washington [98], and in bristlecone fir (A. bracteata) forests of California [20]. In Monterey County, California, the redwood-bigleaf maple/California polypody (Polypodium californicum) community occurs on gently sloping alluvial terraces near streams with boulder or very rocky substrates [139]. It is a rare type within the area's redwood ecosystem [27]. In the Santa Ana [26] and San Gabriel [86] mountains of southern California, bigleaf maple grows in bigcone Douglas-fir (Pseudotsuga macrocarpa) communities.
 |
Mixed-evergreen forests: Bigleaf maple is particularly abundant in mixed-evergreen forests, which are dominated by a mix of Douglas-fir and the evergreen broadleaved species tanoak (Lithocarpus densiflorus), California bay, and/or Pacific madrone. Bigleaf maple is very common in tanoak-Douglas-fir communities in the Klamath-Siskiyou region [15,25]. In southern Oregon and northern California, Oregon white oak or other deciduous species may codominate in mixed-evergreen forests [206].
Hardwood communities: Bigleaf maple is an important to dominant component of many riparian hardwood and some upland oak (Quercus spp.) communities. It occurs in a few other, minor hardwood communities; these are typically in upland wetlands.
Bigleaf maple dominates small stretches of some streams and is associated with many riparian hardwood species. Black cottonwood, red alder, and/or white alder are bigleaf maple's most common associates or codominants in hardwood riparian systems. In coastal California, black cottonwood-bigleaf maple-California bay woodlands are interspersed with mixed-oak, redwood, and Douglas-fir forests [217]. An inventory on the Tillamook Burn revealed a bigleaf maple-red alder association that occurred at 700 to 900 feet (200-275 m) elevation on flat bottomlands of large streams [17]. See the Bigleaf maple communities section of Fire Regimes for more information on this community. Bigleaf maple is a characteristic species in sycamore (Platanus racemosa) riparian forests of the Central Coast Ranges of California [103]. Quaking aspen (Populus tremuloides) often codominates with bigleaf maple near streams in the Sierra Nevada [20]. On the Blodgett Forest Research Station, California, bigleaf maple dominated a few riparian sites. Incense-cedar (Calocedrus decurrens) dominated most riparian areas surveyed [23].
In central and southern California, bigleaf maple occurs only in riparian woodlands or sheltered, mesic sites such as canyon bottoms [148,160]. Codominant or associated species may include boxelder (Acer negundo), Fremont cottonwood (Populus fremontii), and/or white alder [80,86,206]. In the San Gabriel Mountains, a white alder-bigleaf maple vegetation type occupied <5% of the landscape surveyed [196].
Bigleaf maple is a codominant or common component of several hardwood communities that are not exclusively riparian. It occurs in California bay [176], Oregon white oak [176], and canyon live oak (Q. chrysolepis) [43,137] woodlands of the Pacific Northwest and California. Bigleaf maple occurs in canyon live oak-Coulter pine (Pinus coulteri) woodlands of the San Bernardino Mountains [20]. Bigleaf maple is sometimes codominant in California hazelnut communities of the Willamette Valley, Oregon. Bigleaf maple-California hazelnut communities may be successional to western hemlock, but some occur on cobbly floodplains that flood frequently enough to prevent further succession [139].
In southern California, bigleaf maple may dominate or occur in hardwood communities in canyons, other mesic sites, or riparian areas that are surrounded by montane chaparral or coastal sage scrub. California bay and white alder often codominate in these sheltered communities [178]. In Orange County, bigleaf maple dominates shaded aspects of Coal Canyon amidst a mosaic of mixed chaparral, stands of bigcone Douglas-fir, and stands of the northernmost population of Tecate cypress (Cupressus forbesii) [187]. In the San Bernardino Mountains, bigleaf maple-coast live oak-California bay communities are a minor riparian type within the coastal sage scrub matrix [104]. On Santa Cruz Island, bigleaf maple stands occur below 1,510 feet (460 m) on the north side of the island and in Diablo Canyon. Coastal sage scrub predominates across the island [114].
Bigleaf maple is important in a few minor woodlands on year-round springs. In the East Bay Area of California, it is important (17% cover) in California bay-coast live oak/poison-oak (Toxicodendron diversilobum) communities on hillside springs [8]. Bigleaf maple dominates some spring-fed communities in Contra Costa and Alameda counties. California bay, willows (Salix spp.), and/or white alder codominate [7].
Brushfields: In seral brushfields of southern Oregon and northern California, bigleaf maple typically associates or is codominant with red alder; less often, it is the seral dominant [51,52,169]. These brushfields typically persist for a few decades, then succeed to Douglas-fir or mixed-evergreen forests. See Successional Status for more information.Some information in this section came from a review [151].
GENERAL BOTANICAL CHARACTERISTICS:Bigleaf maple is a large, deciduous [118] tree. It is typically about 50 feet (15 m) tall at maturity [151] but sometimes grows more than 80 feet (20 m) [102,138], making it the largest maple species in North America [13]. Trees are generally as wide-spreading as they are tall. Open-grown trees usually develop broad, rounded crowns, with branches that often grow low to the ground [105] and trunks from 2 to 5 feet (0.6-2 m) DBH [40]. Shaded trees are usually pyramidal in form, with narrow crowns [105] and clear, straight boles for one-half to two-thirds of their lengths [40,105]. Bigleaf maple is most frequent and reaches best development in southern Oregon [13]. The champion tree as of 2011 was located in Marion, Oregon. It was 88 feet (27 m) tall, 305 inches (775 cm) in circumference, and had a 104-foot (32 m) spread [9]. On cutover sites, bigleaf maple usually grows in shrubby, multistemmed clumps [83,84]. It also assumes a shrubby form in montane chaparral [151].
Bigleaf maple wood is moderately hard, but it is porous and not strong [105]. Branches of mature trees are massive, spreading [105], and steeply inclined at the tips [118]. Bark is thin [146], rarely more than 0.5 inch (1.3 cm) thick [151,164].
Bigleaf maple typically supports many epiphytes. Mosses, liverworts, and ferns hang from its branches or grow in branch crotches [105,164] (see photo in the Plant communities section). Bigleaf maple's moss load is generally the greatest of all tree species in the Pacific Northwest [164]. Arno [13] estimates that in rain forests of the Pacific Northwest, a bigleaf maple tree supports about 1 ton (0.9 t) of mosses.
 |
| Comparing sizes of a human hand and a bigleaf maple leaf. Photo by Joe Nicholson, Nature photographer, Bugwood.org. |
As the common name claims, the leaves of this species are big. Bigleaf maple has the largest leaves of any North American maple [105], ranging from 4 to 10 inches (10-25 cm) across [99,138]. Trees in western Oregon had a mean area of 43 inches² (280 cm²)/leaf [118]. Male and female flowers are clustered on the same raceme [99,138,146]. (See Breeding system for a more detailed explanation of bigleaf maple flowers.) The fruit is a bristly [102,138], biwinged achene [99] or samara [13,83,146,164] bearing one seed/wing [99].
Bigleaf maple is deep-rooted [47,105,151]; hence, it is ranked low in susceptibility to windthrow [47,48].
Bigleaf maples live about 50 to 200 years [13],154,40,48.
Bigleaf maple tolerates short-term flooding [151], surviving periodic flooding in both active and upper stream channels [208]. It does not tolerate sustained flooding. In British Columbia, Brink [31] observed that bigleaf maples suffered mortality following 100-year floods on the Fraser and Columbia rivers in 1948. Bigleaf maples of all age classes die if floods last for 2 months or more [151].
Raunkiaer [165] life form:| Phenology of bigleaf maple | ||
| Area | Event | Period |
| California, southern | flowers | late March |
| Oregon, Coast Ranges | germination complete | April-May [151] |
| Pacific Northwest | flowers and leaves emerge | late April-early May [13] |
| Northern parts of range, high elevations | flowers | June [151] |
| Across range | germination starts | late January-February [151,199] |
| seeds ripe | August-October [151,225] | |
| seeds disperse | October-January [13,151,225]; some seeds remain on tree until March [151,225] | |
Pollination and breeding system: Insects pollinate bigleaf maple [13,83,113,151]; these pollinators include bees, flies, and beetles (Julian-Gordon 1993 personal observation [113]).
Bigleaf maple is heterdichogamous. This mating system is rare; plant species employing it bear 2 types of flowers on individual trees: 1) those that are functionally female but structurally also have vestigial male flower parts and 2) those that are functionally and structurally male [73,113]. Within and among populations, trees may produce female flowers before male flowers (protogynous) or the reverse (protandrous). The ratio of female:male flowers is apparently about 1:1 [113]. Functionally, this mating system is similar to that of monoecious species, and some authors have labeled bigleaf maple as monoecious [99,138].
Genetic tests of 2 populations in western British Columbia showed bigleaf maple was mostly outcrossing, with low levels of population differentiation. This is consistent with a species with wind-dispersed pollen and seed [109] and a heterdichogamous mating system [113].
Seed production: Bigleaf maple first produces seed when about 10 years old [13,83,84,151]. Arno [13] describes bigleaf maple as a "prolific seed producer"; production is abundant in most years [151]. Open-grown trees may produce a good seed crop every year, but shade-grown trees may have sporadic seed production [40].
 |
| Bigleaf maple seed on gravel. Photo by Susan McDougall @ USDA-NRCS PLANTS Database. |
Wind disperses bigleaf maple seed [78,109]. The samaras "descend like little helicopters, which greatly increases their dispersal" [164].
Seed banking: Seeds do not live more than one winter under field conditions [83,151], so bigleaf maple's seed bank is only transient. In the laboratory, bigleaf maple seed viability dropped from about 88% for fresh seed to about 15% after 1 year in storage at 34 °F (1 °C) [226]. In southwestern British Columbia, viable bigleaf maple seeds were most common in undisturbed forest soils and least common in highly disturbed soils. Soil samples were collected from forest floor to 20-inch (5 cm) depths beneath a Douglas-fir/creambush oceanspray (Holodiscus discolor) forest, then germinated in either the greenhouse or the field [144].
Germination: Bigleaf maple seeds overwinter in the soil, germinating at low winter [151,199] or spring [13] temperatures. They require 60 to 120 days of cold stratification [152,225]. Germination occurs on mineral or organic substrates [83,84,151]. In the Pacific Northwest, Arno [13] observed that in spring, "countless seeds from the previous year's crop" germinate on the forest floor. Seeds that overwinter on the tree may begin germinating before they disperse [225]. Germination rates of 30% to 90% are reported in the laboratory [151,152].
Seedling establishment and plant growth: Bigleaf maple may establish on a variety of sites but generally, seedlings survive only if taproots reach moist soil before the dry season [40,151]. On the Coast Ranges and Klamath Mountains of Oregon, bigleaf maple seedling establishment was better on mesic valley floors than on low hillslopes (42% vs. 29% of total tree seedling composition, respectively) [158]. Most report best establishment occurs on clearcuts or other open sites [13,40,83,84], including canopy gaps (review by [83]). In Douglas-fir plantations in western Oregon, bigleaf maple seedlings established best in open canopies [64,65]. There are a few reports of bigleaf maple establishing best under closed canopies [151]. Some suggest that bigleaf maple is a "seedling banker", maintaining stunted seedlings that persist and grow slowly beneath a closed canopy but that grow rapidly once released (review by [83]).
In general, bigleaf maple seedlings may show better initial establishment than some associated species because of bigleaf maple's relatively large seeds. Many factors, including browsing, may impede this initial success, however. In a planting experiment in a western hemlock habitat type on the McDonald-Dunn Research Forest, Oregon, bigleaf maple and vine maple (Acer circinatum)—both large-seeded species—had higher rates of seedling emergence and survival than the smaller-seeded, associated species salmonberry (Rubus spectabilis) and salal (Gaultheria shallon) on sites protected from browsing. However, browsing pressure on the maples was high on unprotected sites, with maple seedling emergence ranging from only 0% to 4%. This was significantly less than emergence rates of salmonberry and salal on unprotected sites (P<0.001). Three years after sowing, bigleaf maple seedling growth (averaged across sites) was fastest on thinned sites, with seedlings averaging 6.3 inches (16 cm) tall. Growth averaged 4.7 inches (12 cm) on clearcuts and 2.4 inches (6 cm) on unthinned sites [199].
Open-grown seedlings in moist soils may grow 3.3 to 6.6 feet (1-2 m) in one growing season; typically, this growth rate is faster than that of bigleaf maple's conifer associates. Bigleaf maple's growth rate may be cut in half on sites with dense vegetation [151].
Growth generally slows at the pole and sawtimber stages [151]. On the southern coast of British Columbia, bigleaf maple grew most rapidly when 5 to 15 years old. On productive sites, 5-year-old trees averaged 4.5 feet (1.4 m) in height. At 100 years old, trees ranged from 40 to 120 feet (10-40 m) tall [186].
Heavy grazing can slow or stop bigleaf maple growth. A study of historical records describing vegetation at Olympic National Park, Washington, from the mid-1800s through the late 1900s revealed few bigleaf maples and black cottonwoods had grown above browse line after a hunting ban was implemented in 1905 and gray wolves became extirpated around 1920. The authors attributed bigleaf maple's lack of growth to intense browsing by elk herds that are increasing in size without population controls [24].
See Hann and Larsen [88,129] for equations to predict diameter growth of bigleaf maple and other overstory trees of southwestern Oregon.
Vegetative regeneration: Bigleaf maple sprouts from the root crown or stump after top-kill by fire [59,120,151,164,171,184,195,214], cutting [40,58,120,131,151,164,171,198], or herbicide use [44,58,120,131,215]. Sprouts may attain 10 feet (3 m) in 1 year [58,164]. Three-year-old sprouts may reach 17 feet (5 m) tall and 21.5 feet (6.5 m) in crown diameter, with as many as 67 sprouts/root crown [151]. All age classes of bigleaf maple sprout, with sprouts of large trees growing more rapidly than sprouts of small trees [162]. Dale and others [48] developed a model to predict the rate of bigleaf maple sprouting. They predicted that trees from 4 to 20 inches (10-50 cm) in DBH will produce 10 to 50 sprouts/plant after top-kill. Their model was developed for bigleaf maples in the Pacific Northwest [47].
Sprouts grow most quickly on open sites, where they may reach 7 to 13 feet (2-4 m) in their first growing season [83,84]. Bigleaf maples growing under the canopy of old growth may die back and sprout repeatedly (Newton 1984 personal communication cited in [83]).
On the McDonald-Dunn Research Forest, clump size of bigleaf maple sprouts was positively correlated with parent tree stump diameter and negatively correlated with amount of bark removed from the parent during logging (P≤0.05). Mule deer browsing had no significant impact on clump size [198].
SUCCESSIONAL STATUS:Bigleaf maple occurs in all stages of succession, being most common in seral forests. It is considered an indicator of young-seral forests of coastal British Columbia [119]. It is sometimes common in late-seral forests of the Pacific Northwest, however [38]. In surveys along the Tahsish and Artlish rivers on Vancouver Island, bigleaf maple occurred mostly in young-seral Douglas-fir forests, although a few bigleaf maples were noted in mature forests [37]. On the Coast Ranges, bigleaf maple may be a seral dominant in western hemlock vegetation types [63]. The often clumped distribution of bigleaf maple trees in Douglas-fir forests suggests that canopy-gap dynamics may be important in bigleaf maple succession [162], but little information was available on bigleaf maple succession in canopy gaps as of 2011.
Early and midseral stages: Bigleaf maple is common in primary succession on riverine sites, and it may establish on other new substrates as well. Bigleaf maple is described as a primary-successional dominant on fragmented, colluvial soils in coastal British Columbia [119]. Bigleaf maple seedlings were noted, at 8% mean frequency, in primary succession on mudflows the year following the 1980 eruption of Mount St Helens [85].
Periodic disturbances such as flooding, fire, or logging apparently help retain bigleaf maple as an important member of seral plant communities. Riparian zones experience flooding frequently [1]. Bigleaf maple grows on both lower and upper stream terraces (see Site characteristics). The upper terraces, where bigleaf maple is common, may experience fire or wind disturbances more frequently than lower terraces. Even so, in most years, upland riparian plant communities are more protected from flooding, fire, and wind than plant communities outside the riparian zone [1]. In the Stehekin Valley of Washington, Douglas-fir-bigleaf maple and Douglas-fir-ponderosa pine/bigleaf maple forests experienced small- to large-scale wildfires, logging, or both every 30 to 375 years [130].
In riparian communities, bigleaf maple often follows red alder and willows successionally. On upland sites, it may replace Pacific madrone and oaks [151]. Along the Hoh River in Washington, Agee [1] reports that bigleaf maple is a midseral species that occupies midlevel terraces. Red alder and Scouler willow (Salix scouleriana) colonize gravel bars, while red alder dominates 1st-level, 80- to 100-year-old terraces. Bigleaf maple, Sitka spruce, and black cottonwood dominate the next-oldest and higher, 2nd terrace. Sitka spruce-western hemlock and western hemlock, respectively, dominate the successively older, 3rd and 4th terraces [1]. Another survey along the Hoh River had similar findings, with red alders dominating floodplains, bigleaf maple dominating communities on 1st and 2nd terraces, and western hemlock dominating uppermost, 3rd terraces [60].
Conifers may replace bigleaf maple successionally in riparian zones. A Sitka spruce-black cottonwood-bigleaf maple forest may replace riparian hardwoods on the Olympic Peninsula (review by [63]). On banks of the Queets River in Olympic National Park, bigleaf maple and black cottonwood occur in midsuccession, after red alder. In turn, they are eventually replaced by Sitka spruce. Turnover rate from red alder to bigleaf maple and black cottonwood dominance ranges from 60 to 80 years; bigleaf maple and Sitka spruce may remain codominant "for centuries" before Sitka spruce overtops the hardwoods [19]. Incense-cedar and other shade-tolerant conifers may replace bigleaf maple in the Sierra Nevada (Fryer 2001 personal observation).
After logging, bigleaf maple and other sprouting species [151,169] occupy clearings that succeed to conifers [63,105,169,224]. Bigleaf maple is common in 2nd-growth forests of southwestern British Columbia, but it is rarely dominant [163]. In western hemlock-Douglas-fir forests in coastal British Columbia, bigleaf maple frequency was 1% on a site logged 13 years previously, 4% on a site logged 42 years previously, and 8% on an unlogged site [116]. Such sites may convert to red alder-bigleaf maple or red alder brushfields if conifer seed sources are lacking [220].
Bigleaf maple increased in cover following logging and slash burning on the H. J. Andrews Experimental Forest in southern Oregon. Western hemlock dominated the overstory prior to logging. Bigleaf maple abundance was [51,52]:
| Mean percent bigleaf maple cover (and frequency) before and after logging and slash burning on 3 sites on the H. J. Andrews Experimental Forest [51,52] | |
| Before logging (1962) | 0.2 (3.3) |
| After logging (1963) | 0.3 (3.3) |
| Postfire year 1 (1964) | 1.1 (4.9) |
| Postfire year 2 (1965) | 1.3 (3.3) |
| Postfire year 3 (1966) | 2.1 (4.9) |
| Postfire year 4 (1967) | 2.2 (6.6) |
| Postfire year 5 (1968) | 2.3 (6.6) |
Studies in the Cascade Range of southwestern Washington and western Oregon showed bigleaf maple was significantly more common in young and mature than in old-growth Douglas-fir forests (P≤ 0.5). However, on the Coast Ranges of Oregon, bigleaf maple was most common in old growth (P<0.1). All stands sampled originated after stand-replacing wildfires. Stand age classes were [188]:
<80 years = young
80 to 195 years = mature
>195 years = old growth
Bigleaf maple patches within a conifer matrix may shrink or wink out without disturbances such as fire or logging. A comparison of 1939 and 1993 landscapes on the Coast Ranges of Oregon showed early- to midsuccessional patches of bigleaf maple and/or red alder had declined in number, size, and total area. Declines were attributed to fire exclusion and reduced logging and grazing [117]. Two surveys made of the same areas in Yosemite Valley—one from 1932 to 1936 and the other from 1988 to 1999—indicated that large-diameter bigleaf maples (≥4 inches (10 cm) DBH) had declined over time, although differences in methods and inability to relocate the original plots made direct comparisons impossible [135].
Plant response to fire provides more information on bigleaf maple postfire succession and growth.
Late-seral and old-growth stages: Bigleaf maple may form part of the canopy in late succession, sometimes sharing dominance with conifers or other hardwoods. Bigleaf maple is a common component of old-growth, mixed-evergreen forests of southwestern Oregon and northern California [25,76,77]. It is the most common deciduous tree in old-growth Douglas-fir forests of Horse Rock Ridge Research Natural Area in Oregon [46]. It also occurs in old-growth redwood forests in California [72].
Bigleaf maple is noted as a late-seral species in the Willamette Valley [175]. Following stand-replacement fire, most vegetation succeeds from a grass stage to an oak woodland, then an oak forest, and last to a Douglas-fir-grand fir-bigleaf maple forest (Franklin and Dyrness 1973 cited in [50]). Comparing plant community composition of the European-American settlement period with that of the early 1960s, Habeck [82] found bigleaf maple was encroaching into Oregon white oak woodlands on mesic sites, while Douglas-fir was encroaching on both mesic and dry sites. Habeck suggested that eventually, bigleaf maple may replace both Oregon white oak and Douglas-fir successionally [82]. Another viewpoint is that Douglas-fir becomes the late-successional dominant on dry sites, while bigleaf maple codominates with Douglas-fir on moist sites (review by [63]).
Immediate fire effect on plant:
Bigleaf maple is relatively thin-barked [146], so it is not fire-resistant [48,151,164]. Fire top-kills bigleaf maple in most size classes [83,184,195,214], although large, mature trees that have developed thick bark may survive [47] moderate-severity fires. Severe fire kills most bigleaf maples ([61,94], review by [162]), but a few may survive severe fire ([61], review by [162]).
Postfire regeneration strategy [194]:
Tree with a sprouting root crown
Crown residual colonizer (on site, initial community)
Initial off-site colonizer (off site, initial community)
Secondary colonizer (on- or off-site seed sources)
Fire adaptations and plant response to fire:
Fire adaptations:
Adventitious buds on the root crown enable bigleaf maple to sprout after most fires [146].
Plant response to fire: Bigleaf maple sprouts from the root crown after top-kill by fire [59,61,94,120,146,151,164,171,184,184,195,195,214,214].
As of 2011, there were no reports of postfire seedling establishment by bigleaf maple, although few studies reported whether bigleaf maple's postfire regeneration was from root crown sprouts or seeds. Given bigleaf maple's ability to establish from seed after logging (see Seedling establishment and Successional Status), bigleaf maple seedlings may establish on new burns if surrounding vegetation is not dense enough to interfere with bigleaf maple seedling establishment and growth. Trees that survive or are missed by fire may provide on-site seed. Because wind readily disperses bigleaf maple seed, off-site trees may provide additional bigleaf maple seed. Since bigleaf maple has only a transient seed bank, it is unlikely that viable seeds remain in the soil after the fire season. However, bigleaf maple disperses seed after the fire season, through late fall and winter, so if a seed source is available, bigleaf maple seedlings may emerge on new burns in the first winter or spring (see Seasonal Development) after fire.
Fire generally favors bigleaf maple [12]. Studies in Washington [3,61], Oregon [136,169], and northern California [95,171] showed postfire establishment of bigleaf maple, even after a severe fire [61].
Red alder and bigleaf maple often dominate early stages of postfire succession in Douglas-fir forests of the Pacific Northwest [12]. Two months after the lightning-ignited Hoh Fire in Olympic National Park, all bigleaf maples on burned plots were top-killed or killed. Before fire, bigleaf maple grew on well-drained, stable terraces above the Hoh River. The forest type on these low-level terraces was Sitka spruce, with a western hemlock-Douglas-fir forest type on upland sites. The wildfire was patchy, with some crown fire, some crown scorch, and some unburned areas; most crowning occurred on upland sites. The fire often extinguished when it reached moist Sitka spruce/bigleaf maple terraces. Bigleaf maple was a minor component in most riparian communities, although after the fire it showed the greatest relative loss of live basal area compared to the dominant conifers [3].
| Bigleaf maple basal area (m²/ha) 3 months after the Hoh Fire in Olympic National Park [3] | ||
| Burned plots | Unburned plots | |
| Live stems | 0 | 1.7 |
| Dead stems | 0.9 | 0 |
Bigleaf maple sprouted after a severe wildfire near Bellingham, Washington. A gas pipeline near Whatcom Creek leaked, exploded, and burned a black cottonwood-red alder-bigleaf maple forest. The burned area was surveyed in postfire year 1. Seventy-two bigleaf maples grew in the area before the fire. Of these, 59 trees (82%) survived. Eighteen of the surviving trees retained a live crown, and 41 were top-killed and sprouted. Bigleaf maples with surviving crowns, and those that died, ranged from 4 to 24 inches (10-60 cm) in diameter. Sprouting trees were 4 to 12 inches (10-30 cm) in diameter [61].
A 38-year study on the H. J. Andrews Experimental Forest showed bigleaf maple abundance peaked in early seral stages following clearcutting and slash burning. Bigleaf maple density peaked in posttreatment years 18 to 21, at around 200 stems/ha. Posttreatment mortality was partially attributed to successional replacement (26.3%) and mechanical damage such as windthrow (13.6%); most mortality was due to unknown causes (58.4%) [136].
On the Oregon Coast Ranges, bigleaf maples had mixed responses to hardwood thinning, spraying, and prescribed fire. Some trees were killed by the treatments, while others were only top-killed and sprouted. Treatments were initiated to reduce hardwood dominance on brushfields that developed after the Sitka spruce-western hemlock-Douglas-fir overstory was logged. Large hardwoods were logged in March and April 1974; 2, 4, 5-T was sprayed on 2 June 1974; and the site was burned on 9 August 1974. The fire was "intense", and most of the site burned [169].
Bigleaf maple sprouts were noted on broadcast-burned, pile-burned, and unburned sites following clearcutting in Douglas-fir-ponderosa pine forests on the Klamath National Forest, California. Across sites, sprouts reached up to 16 feet (5 m) by postfire year 6 [95].
Hardwood sprouts grew rapidly after wildfire or logging in northern California. The Three Creeks Wildfire in Humboldt County, California, started in heavy slash left after a mixed-evergreen forest was logged. The mostly stand-replacement fire burned in August, 1951. Among bigleaf maple, Pacific madrone, Oregon white oak, tanoak, and Pacific dogwood (the 5 most common sprouting hardwoods), bigleaf maple had produced the greatest number of sprouts/root crown by postfire year 2, and its sprouts were the tallest. Bigleaf maple also showed a strong sprouting response after a mixed-evergreen forest in Trinity County, California, was logged. Among the same 5 sprouting hardwoods, bigleaf maple showed the strongest tendency for the numbers of sprouts produced/clump to increase with diameter of the top-killed tree. Height of bigleaf maple sprouts also increased with tree diameter, although this response was greater for Pacific madrone and Oregon white oak than for bigleaf maple. Pooled data, collected from burned or logged sites, for growth of bigleaf maple are shown below [171].
| Pooled growth responses of bigleaf maple on wildfire or logged sites in northern California. n=10 bigleaf maple sprout clumps [171]. | ||||
| Variable | ||||
| mean | range | mean | range | |
| Height of tallest sprout in clump (feet) | 9.8 | 6.8-13.1 | 12.8 | 7.5-17.1 |
| Crown diameter of sprout clump (feet) | 11.5 | 6.8-15.5 | 14.7 | 10.4-21.5 |
| Sprouts/clump | 78 | 14-143 | 37 | 8-67 |
An energy value of about 8,400 BTUs/pound (oven-dry) is reported for bigleaf maple wood ([222], review by [110]).
Several studies provide site-specific information on fuel structure or loads that may apply to similar sites elsewhere. Fierke and others [57] describe stand structure for black cottonwood communities with bigleaf maple. Their data were collected in stands by the Willamette River, Oregon [57,57]. Spies and Franklin [189] provide a description of forest floor, coarse woody debris, litter, and stand structure characteristics of Douglas-fir forests of the Pacific Northwest. Their information is based on 196 sample sites across the Cascade Range in Washington and Oregon, the Northern Coast Ranges of Oregon, and the Siskiyou Mountains of Oregon. Young (<80 years), mature (80-195 years), and old-growth (>195 years) stands were sampled; bigleaf maple was among the dominant deciduous trees in these stands [189]. Rickard [167] gives measurements of litterfall in a 2nd-growth Douglas-fir forest with scattered bigleaf maples and Oregon white oaks. Data were collected on the west bank of the Columbia River [167].
The following citations provide models that help predict fuel loads in plant communities where bigleaf maple is important:Fire regimes: Bigleaf maple experiences a wide variety of fire regimes across its distribution. Western hemlock, Douglas-fir, and mixed-evergreen forests lie on a north-south gradient along the Pacific Coast, with Douglas-fir, then ponderosa pine, attaining increasing dominance to the south. Fire regimes historically reflected this gradient, with long-interval, more severe fire to the north and shorter-interval, less severe fires to the south.
Western hemlock-Sitka spruce forests have stand-replacement [123] and mixed-severity (review by [159]) fires. Douglas-fir and mixed-evergreen forests of the Pacific Northwest and northern California experience a mix of stand-replacing and understory fires. Historically, ponderosa pine and mixed-conifer forests of southern Oregon and northern California also had a mixed-severity fire regime, with a prevalence of low-severity surface fires and some patchy stand-replacement fires. For all these types, most fires occurred in summer and fall (reviews in [197]).
Bigleaf maple often lies within the riparian zones of these forests. Historical fire regimes for riparian zones of the western United States are not well studied [182,185]. In general, fires are less frequent in riparian and wetland areas of the West than in drier areas [14]. At low- to midelevations in the southern Cascade Range and Klamath Mountain regions of California, fire-return intervals in riparian areas overall are about twice those of surrounding areas, but fire intensity is generally greater when the riparian areas burn [182]. Fire-return intervals near intermittent and ephemeral streams are likely similar to those of surrounding areas [185]. In most years, perennial stream communities may serve as firebreaks (review by [182]).
Descriptions of fire regimes in plant communities in which bigleaf maple occurs follow. See the Fire Regime Table for further information on fire regimes of plant communities in which bigleaf maple may occur. Find further fire regime information for the plant communities in which this species may occur by entering the species name in the FEIS home page under "Find Fire Regimes".
Western hemlock-dominated forests: Western hemlock-Sitka spruce forests on the coast of southern British Columbia historically experienced mixed understory-crown and crown fires. Fire-return intervals on the coast ranged from 6 to 130 years, with an average of 75.4 years (review by [159]). Western hemlock-Douglas-fir forests on the Olympic Peninsula have mostly long-interval, stand-replacement fires. These infrequent crown fires occur mostly in drought years, with fires returning about every 500 to 750 years [106].
Douglas-fir-dominated forests: Douglas-fir forests in mainland western British Columbia historically experienced mixed-severity fires, with a prevalence of even-aged stands suggesting a history of mostly stand-replacement fires. Fire-return intervals ranged from 35 to 256 years, averaging 107.5 years (review by [159]). A fire history study on Vancouver Island suggested a regime of mostly low-severity understory fires in a Douglas-fir-grand fir-bigleaf maple forest. The community occurred in a ravine. Most trees were 100 to 150 years old, and the oldest were 350 to 500 years old [142].
A fire history study on Orcas Island, Washington, showed a regime of frequent, low-severity understory fires in Douglas-fir forests with scattered bigleaf maples, western hemlocks, and shore pines (Pinus contorta var. contorta). Fire-return interval averaged 28.3 years from 1773 to 1893, with no fires since then. By the 1990s, stand structure had changed from a primarily open to a closed canopy on most sites. The authors predicted that because fire exclusion led to stand closure, the next fire is likely to be stand-replacing [161].
Along the McKenzie River on the western slope of the Cascade Range, Oregon, bigleaf maple is common in early-seral Douglas-fir-western hemlock-western redcedar forests. These forests had a fire rotation interval of about 162 years in the presettlement period, with at least 77% of the landscape burning from 1525 to 1575. Average fire-return interval during the settlement period (1850-1924) was 40 years [218]. Other fire studies in the southern Cascade Range found fire-return intervals from 18.6 to 26.3 years in Douglas-fir forests; fires were mostly low-severity surface fires (review by [184]). Skinner and Taylor [184] caution that actual fire-return intervals might have been shorter because many old Douglas-firs had fire scars that healed over after low-severity fires. These scars were not apparent until after the area was logged and entire stumps could be examined [184].
A fire history in southwestern Oregon showed bigleaf maple was associated with productive, low-elevation sites that were relatively close to old-growth stands (P=0.15). The area was on the Siouxon Creek Watershed in a Douglas-fir-western hemlock forest. It had experienced both large, "intense" and small, patchy fires. The oldest Douglas-firs in old-growth areas established around 1500. Numerous small, patchy fires occurred in the early 1800s, and a large, mostly stand-replacement fire occurred in 1902, with portions of the 1902 burn apparently reburning 2 or 3 times in the next 80 years. Between 1479 and 1938, 12 separate fires were detected on the 40,000-acre (16,000 ha) watershed [76].
Bigleaf maple has invaded formerly open areas where fire has been excluded in the Willamette Valley. Bigleaf maple, and to a larger extent, Douglas-fir, is encroaching into areas that were historically grasslands or Oregon oak woodlands [82]. Fires set by American Indians and then by European-American settlers maintained the Willamette Valley as grasslands and woodlands [30].
Mixed-evergreen forests: Fire regimes in mixed-evergreen forests region are variable [2,134]. These forests often occupy steep slopes and have abrupt elevation changes and highly variable soils, so vegetation types can shift quickly. Mixed-evergreen forests in the Klamath-Siskiyou region have mostly mixed understory and stand-replacement fires that leave patches of unburned, underburned, and crown-fire burned areas across the landscape [5,76,77,134]. Fire-return intervals vary from 10 years or less for Oregon white oak and canyon live oak woodlands to about 15 years for low-elevation (≤4,000 feet (1,000 m)) Douglas-fir/hardwood forests and about 160 or more years for midelevation (4,000-6,000 feet (1,000-2,000 m)) white fir forests.
The Big Bar (1999) and Quartz (2001) wildfires in the Klamath-Siskiyou region of southern Oregon were mostly low- to moderate-severity understory fires at high elevations and north slopes, especially on sites with large-diameter trees. Low-elevation sites, south-facing slopes, and sites with small-diameter trees and/or ladder fuels were most likely to experience stand-replacement fires. The plant communities that burned were Douglas-fir-tanoak-ponderosa pine at low elevations and white fir at high elevations; elevations ranged from 2,580 to 6,043 feet (785-1,842 m). Bigleaf maple was described as a dominant hardwood in these communities [5].
Fire studies in riparian zones within mixed-evergreen forests are rare (review by [67]). Limited studies show median fire-return intervals of 16 to 42 years in black cottonwood sites near Lake Shasta (Skinner 2000 cited in [67]) and 12 to 19 years in riparian Douglas-fir forests on the Klamath National Forest. From 1627 to 1992, the average area burned in riparian Douglas-fir forests was 860 acres (350 ha), with 16 fires larger than 1,200 acres (500 ha) [201]. Fires burned mostly in midsummer or early fall [202].
Bigleaf maple communities: Bigleaf maple-dominated stands embedded within mixed-evergreen forests comprise a very small portion of the mixed-evergreen ecosystem, and little information is available on fire behavior in these stands. With the Biscuit Wildfire in southwestern Oregon, 82% of the hardwood forest types that burned experienced very low- to low-severity fire. Hardwood forest types were classified as nonstocked (<10% commercial timber species). In contrast, 84% of other nonstocked forest types experienced moderate- to high-severity fire. The bigleaf maple type comprised about 2% of hardwood-dominated stands; tanoak comprised about 63% [16].
Bigleaf maple stands may occur on sites that are too moist to burn in most years. A 1961 to 1962 inventory of the Tillamook Burn found that the bigleaf maple-red alder association was the only type that did not burn during the initial, 1933 wildfire. The association occurred in mesic stream bottomlands. Red alder showed fire scars from the 1939 and 1945 fires, but bigleaf maple was apparently not damaged. At the time of the study, the bigleaf maple overstory was about 90 feet (30 m) tall, with bigleaf maple cover ranging from 1% to 70% [17].
Chaparral: Low-elevation [124] chaparral communities have almost exclusively stand-replacement fires, about every 50 to 125 years. Coastal sage scrub communities also have stand-replacement fires but with a wider range of fire-return intervals, from 20 to 150 years [126]. Montane chaparral communities, which border mixed-conifer ecosystems of the Sierra Nevada, experience both mixed-severity and stand-replacement fires that average 50 and 95 years, respectively [125].
Fire may skip over hardwood communities in ravines and mesic canyons that are surrounded by chaparral. In the Santa Monica Mountains, bigleaf maple-California bay-white alder communities in deep canyons escaped wildfires that burned in adjacent chamise (Adenostoma fasciculatum) or mixed-chaparral communities [178].
FIRE MANAGEMENT CONSIDERATIONS:In the Sierra Nevada, a prescribed fire in riparian zones of the Blodgett Research Station significantly reduced biomass of coarse woody debris and groundlayer vegetation in the short term compared to prefire biomass. The 21-23 October fire was conducted near 1st- and 2nd-order streams dominated by bigleaf maple or incense-cedar. Surveys were conducted at postfire week 2 for fuels and postfire year 2 for vegetation. The fire killed only 8 large-diameter (4.6-15.9 inches (11.7-40.4 cm)) trees, which comprised 4.4% of the total of all large trees. Large, fire-killed trees were not differentiated by species [23].
Bigleaf maple is important as a wildlife tree and for plant community diversity in general [153]. Prescribed fires in Douglas-fir forests may help maintain large Douglas-firs in the overstory and create patches of bigleaf maple, red alder, and other hardwood within the conifer forest. Arno [12] reports that the benefits of using prescribed fire in Douglas-fir forests of the Pacific Northwest and coastal California include removing understory conifer and ladder fuels, retarding succession to late-seral species such as western hemlock, and creating openings "of all sizes" that favor bigleaf maple and other deciduous woody species.Mule deer [151,170], elk [24,151], mountain beavers [45,151], other rodents [151,162], and invertebrates browse bigleaf maple foliage [151]. Mountain beavers on the Tillamook Burn used bigleaf maple as a winter food [151]. Various animals, including Douglas's squirrels [13,32], northern flying squirrels [33], finches, and evening grosbeaks [13] eat the seeds, especially during winter when other foods are scarce [13,32]. In the spring, deer mice were observed eating germinating bigleaf maple seeds in the Sierra Nevada [111].
Early-seral, riparian communities with bigleaf maples are habitat for many small animals. A survey of riparian zones on the Oregon Coast Ranges found that some rodent and amphibian species preferred seral red alder-bigleaf maple brushfields that developed after the Douglas-fir overstory was logged. Numbers of creeping voles, Townsend's chipmunks, western redbacked salamanders, and Pacific giant salamanders captured were greater the year after logging compared to before logging [39]. McComb and McGarigal [141] provide site descriptions and lists of small mammals and amphibians that use Douglas-fir-red alder riparian communities with bigleaf maple as habitat in Lincoln County, Oregon.
Many bird species use riparian habitats in which bigleaf maple is prominent [107]; some depend on a component of deciduous trees in general [107] or are associated with bigleaf maple in particular. Gleaning birds such as brown creepers forage on large bigleaf maples [140]. In studies on the western slope of the Cascade Range in Oregon, dusky and Hammond's flycatchers were positively associated with small- (0.4-4 inches (1-10 cm)) and medium- (4-20 inches (10-50 cm)) diameter bigleaf maples (P<0.04) [71]. Barred owls prefer uneven-aged mosaics of bigleaf maple and other early-successional tree species over mature conifer forests. Allen [6] speculates that increased numbers of barred owls in the Pacific Northwest are partially due to establishment and growth of bigleaf maple and other early-successional trees after logging.
Downed bigleaf maple branches are a component of channel-floodplain woody debris and log jams [41,108]. Such debris slows stream channel flow [68,208], and retained organic matter enhances habitat quality for many aquatic animals [68,122]. Retained debris enhances steelhead [204] and other fisheries [181,204]. In the Pacific Northwest, hardwoods such as bigleaf maple are associated with high biomass of aquatic macroinvertebrates (Piccolo and Wipfli 2002 cited in [117]). In the Queets River, large woody debris of bigleaf maple and other large hardwoods appeared to decay more rapidly than that of Sitka spruce and other conifers [108], so organic matter from decaying hardwoods was likely more readily available to aquatic detritus feeders than organic matter from conifers.
Palatability and nutritional value: Sampson and Jesperson [177] rate the palatability of bigleaf maple foliage as good to poor for mule deer, fair to poor for domestic goats, and poor for domestic sheep, cattle, and horses. Palatability is rated good for Roosevelt elk [42].
Bigleaf maple foliage collected in the Pacific Northwest was fairly high in nitrogen and calcium content. See Tarant and others [200] for a nutritional analysis of bigleaf maple foliage. Bigleaf maple bark is also high in calcium. This probably accounts for the unusually high load of epiphytes that bigleaf maple typically carries [121]. Valachovic and others [213] provide nutritional analyses of bigleaf maple litter.
Cover value: Wild ungulates, rodents, birds, and insects use bigleaf maple as cover, nesting, and/or foraging habitat.
Mule deer select bigleaf maple communities as habitat. On the Tillamook Burn, their density averaged 54 deer/mile² in bigleaf maple/creeping snowberry the community, the highest among 5 communities studied. Mule deer populations were surveyed from 1964 to 1969 [101]; this was 13 to 18 years after the last reburn. Mule deer probably used the bigleaf maple/creeping snowberry community for foraging and bedding in winter because the open slopes were relatively warm. They avoided the community in summer [100]. A 1964 study on the Tillamook Burn found mule deer sightings in bigleaf maple/creeping snowberry communities were high in late winter (15% of total sightings), declined from March to May (2.2%-4.6%), increased through summer (12%), and peaked in October (17.8%). In contrast to the earlier study, mule deer preferred these communities for summer bedding [145].
On the Olympic Peninsula, Roosevelt elk used mature hardwood forests in the Queets River valley floor as spring, summer, and fall habitat. Red alder, black cottonwood, and bigleaf maple dominated these forests [180].
Dusky-footed woodrats may build nests in bigleaf maple [56].
Many bird species nest in bigleaf maple [13], including harlequin ducks [35] and pileated woodpeckers [91,92]. Pileated woodpeckers on Vancouver Island excavated cavities in large bigleaf maples for nesting, although they also selected other large trees (x = 32 inches (82 cm)) DBH) for cavity nesting. Regardless of the tree species selected for nesting, areas around active cavity trees had greater numbers of bigleaf maple than areas without active cavity trees (P=0.05) [92].
Bald eagles use bigleaf maples as roost trees [190].
Many insects use the furrowed, scaly bark of large bigleaf maples as habitat [140].
VALUE FOR REHABILITATION OF DISTURBED SITES:The sap can be rendered into maple syrup [174], although the yield is less than that of sugar maple (Acer saccharum) [151].
American Indians historically harvested bigleaf maple branches and twigs. They used its bark for making rope, the wood for utensils and canoe paddles [13], and the shoots for baskets [10,11,36]. Selected bigleaf maples were burned or pruned frequently to force sprouting [11].
OTHER MANAGEMENT CONSIDERATIONS:Bigleaf maple can interfere with the establishment and growth of commercial conifer species [83,84,131,164,164] because its sprouts usually grow faster than conifer seedlings [83]. These sources provide information controlling bigleaf maple with herbicides: [44,58,120,131,131,151,155,162,198,215]. Norris and others [154] provide a guide for using and some effects of herbicides in riparian zones.
Bigleaf maple can also have positive effects on conifer growth. Litter and debris from bigleaf maple trees and sprout clumps can increase forest floor and soil nutrients and accelerate nutrient and litter turnover in Douglas-fir and western hemlock forests [209]. In western Oregon, soil nitrogen and potassium levels were greater under bigleaf maples than under Douglas-firs. Nutrient content (N, K, P, Ca, Mg) and turnover rate of litter was significantly greater under bigleaf maples, as were turnover rates for forest floor nutrients (P<0.05) [64,66]. Fried [64] concluded that complete removal of bigleaf maple from Douglas-fir plantations could reduce overall nutrient availability. Bigleaf maple may be a nurse tree for western redcedar, which often establishes in the humus layer beneath bigleaf maple [121].
Bigleaf maple is among the many native species that are alternate, non-oak hosts of Phytophthora ramorum, the pathogen causing sudden oak death disease [69,168].
Bigleaf maples burdened with heavy epiphyte loads are more susceptible to windthrow than unencumbered trees [151].| Fire regime information on vegetation communities in which bigleaf maple occurs. This information is taken from the LANDFIRE Rapid Assessment Vegetation Models [128], which were developed by local experts using available literature, local data, and/or expert opinion. This table summarizes fire regime characteristics for each plant community listed. The PDF file linked from each plant community name describes the model and synthesizes the knowledge available on vegetation composition, structure, and dynamics in that community. Cells are blank where information is not available in the Rapid Assessment Vegetation Model. | |||||||||
|
|||||||||
| Pacific Northwest | |||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||
| Northwest Woodland | |||||||||
| Oregon white oak-ponderosa pine | Replacement | 16% | 125 | 100 | 300 | ||||
| Mixed | 2% | 900 | 50 | ||||||
| Surface or low | 81% | 25 | 5 | 30 | |||||
| Pine savannah (ultramafic) | Replacement | 7% | 200 | 100 | 300 | ||||
| Surface or low | 93% | 15 | 10 | 20 | |||||
| Ponderosa pine | Replacement | 5% | 200 | ||||||
| Mixed | 17% | 60 | |||||||
| Surface or low | 78% | 13 | |||||||
| Oregon white oak | Replacement | 3% | 275 | ||||||
| Mixed | 19% | 50 | |||||||
| Surface or low | 78% | 12.5 | |||||||
| Subalpine woodland | Replacement | 21% | 300 | 200 | 400 | ||||
| Mixed | 79% | 80 | 35 | 120 | |||||
| Northwest Forested | |||||||||
| Sitka spruce-western hemlock | Replacement | 100% | 700 | 300 | >1,000 | ||||
| Douglas-fir (Willamette Valley foothills) | Replacement | 18% | 150 | 100 | 400 | ||||
| Mixed | 29% | 90 | 40 | 150 | |||||
| Surface or low | 53% | 50 | 20 | 80 | |||||
| Oregon coastal tanoak | Replacement | 10% | 250 | ||||||
| Mixed | 90% | 28 | 15 | 40 | |||||
| Ponderosa pine (xeric) | Replacement | 37% | 130 | ||||||
| Mixed | 48% | 100 | |||||||
| Surface or low | 16% | 300 | |||||||
| Dry ponderosa pine (mesic) | Replacement | 5% | 125 | ||||||
| Mixed | 13% | 50 | |||||||
| Surface or low | 82% | 8 | |||||||
| Douglas-fir-western hemlock (dry mesic) | Replacement | 25% | 300 | 250 | 500 | ||||
| Mixed | 75% | 100 | 50 | 150 | |||||
| Douglas-fir-western hemlock (wet mesic) | Replacement | 71% | 400 | ||||||
| Mixed | 29% | >1,000 | |||||||
| Mixed conifer (southwestern Oregon) | Replacement | 4% | 400 | ||||||
| Mixed | 29% | 50 | |||||||
| Surface or low | 67% | 22 | |||||||
| California mixed evergreen (northern California and southern Oregon) | Replacement | 6% | 150 | 100 | 200 | ||||
| Mixed | 29% | 33 | 15 | 50 | |||||
| Surface or low | 64% | 15 | 5 | 30 | |||||
| Lodgepole pine (pumice soils) | Replacement | 78% | 125 | 65 | 200 | ||||
| Mixed | 22% | 450 | 45 | 85 | |||||
| Pacific silver fir (low elevation) | Replacement | 46% | 350 | 100 | 800 | ||||
| Mixed | 54% | 300 | 100 | 400 | |||||
| Pacific silver fir (high elevation) | Replacement | 69% | 500 | ||||||
| Mixed | 31% | >1,000 | |||||||
| Spruce-fir | Replacement | 84% | 135 | 80 | 270 | ||||
| Mixed | 16% | 700 | 285 | >1,000 | |||||
| California | |||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | |||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
||||||
| California Shrubland | |||||||||
| Coastal sage scrub | Replacement | 100% | 50 | 20 | 150 | ||||
| Coastal sage scrub-coastal prairie | Replacement | 8% | 40 | 8 | 900 | ||||
| Mixed | 31% | 10 | 1 | 900 | |||||
| Surface or low | 62% | 5 | 1 | 6 | |||||
| Chaparral | Replacement | 100% | 50 | 30 | 125 | ||||
| Montane chaparral | Replacement | 34% | 95 | ||||||
| Mixed | 66% | 50 | |||||||
| California Woodland | |||||||||
| California oak woodlands | Replacement | 8% | 120 | ||||||
| Mixed | 2% | 500 | |||||||
| Surface or low | 91% | 10 | |||||||
| Ponderosa pine | Replacement | 5% | 200 | ||||||
| Mixed | 17% | 60 | |||||||
| Surface or low | 78% | 13 | |||||||
| California Forested | |||||||||
| California mixed evergreen | Replacement | 10% | 140 | 65 | 700 | ||||
| Mixed | 58% | 25 | 10 | 33 | |||||
| Surface or low | 32% | 45 | 7 | ||||||
| Coast redwood | Replacement | 2% | ≥1,000 | ||||||
| Surface or low | 98% | 20 | |||||||
| Mixed conifer (north slopes) | Replacement | 5% | 250 | ||||||
| Mixed | 7% | 200 | |||||||
| Surface or low | 88% | 15 | 10 | 40 | |||||
| Mixed conifer (south slopes) | Replacement | 4% | 200 | ||||||
| Mixed | 16% | 50 | |||||||
| Surface or low | 80% | 10 | |||||||
| Aspen with conifer | Replacement | 24% | 155 | 50 | 300 | ||||
| Mixed | 15% | 240 | |||||||
| Surface or low | 61% | 60 | |||||||
| Jeffrey pine | Replacement | 9% | 250 | ||||||
| Mixed | 17% | 130 | |||||||
| Surface or low | 74% | 30 | |||||||
| Mixed evergreen-bigcone Douglas-fir (southern coastal) | Replacement | 29% | 250 | ||||||
| Mixed | 71% | 100 | |||||||
| Interior white fir (northeastern California) | Replacement | 47% | 145 | ||||||
| Mixed | 32% | 210 | |||||||
| Surface or low | 21% | 325 | |||||||
| *Fire Severities— Replacement: Any fire that causes greater than 75% top removal of a vegetation-fuel type, resulting in general replacement of existing vegetation; may or may not cause a lethal effect on the plants. Mixed: Any fire burning more than 5% of an area that does not qualify as a replacement, surface, or low-severity fire; includes mosaic and other fires that are intermediate in effects. Surface or low: Any fire that causes less than 25% upper layer replacement and/or removal in a vegetation-fuel class but burns 5% or more of the area [89,127]. |
|||||||||
1. Agee, James K. 1988. Successional dynamics in forest riparian zones. In: Raedeke, Kenneth J., ed. Streamside management: riparian wildlife and forestry interactions. Institute of Forest Resources Contribution No. 58. Seattle, WA: University of Washington, College of Forest Resources: 31-43. [7657]
2. Agee, James K. 1993. Fire ecology of Pacific Northwest forests. Washington, DC: Island Press. 493 p. [22247]
3. Agee, James K.; Huff, Mark H. 1980. First year ecological effects of the Hoh Fire, Olympic Mountains, Washington. In: Martin, Robert E.; Edmonds, Robert L.; Faulkner, Donald A.; Harrington, James B.; Fuquay, Donald M.; Stocks, Brian J.; Barr, Sumner, eds. Proceedings, sixth conference on fire and forest meteorology; 1980 April 22-24; Seattle, WA. Washington, DC: Society of American Foresters: 175-181. [10201]
4. Agee, James K.; Kertis, Jane. 1987. Forest types of the North Cascades National Park Service Complex. Canadian Journal of Botany. 65: 1520-1530. [6327]
5. Alexander, John D.; Seavy, Nathaniel E.; Ralph, C. John; Hogoboom, Bill. 2006. Vegetation and topographical correlates of fire severity from two fires in the Klamath-Siskiyou region of Oregon and California. International Journal of Wildland Fire. 15: 237-245. [64044]
6. Allen, Arthur W. 1987. Habitat suitability index models: barred owl. Biol. Rep. 82 (10.143). Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service. 17 p. [11719]
7. Allen-Diaz, Barbara; Bartolome, James W.; McClaran, Mitchell P. 1999. California oak savanna. In: Anderson, Roger; Fralish, James S.; Baskin, Jerry M., eds. Savannas, barrens, and rock outcrop plant communities of North America. Boston, MA: Cambridge University Press: 322-340. [83865]
8. Allen-Diaz, Barbara; Jackson, Randall D.; Phillips, Catherine. 2001. Spring-fed plant communities of California's East Bay Hills oak woodlands. Madrono. 48(2): 98-111. [40591]
9. American Forests. 2011. Bigleaf maple: Acer macrophyllum. In: National register of big trees, [Online]. Available: http://www.americanforests.org/resources/bigtrees/ [2011, August 8]. [83258]
10. Anderson, Kat. 1991. Wild plant management: cross-cultural examples of the small farmers of Jaumave, Mexico, and the southern Miwok of the Yosemite region. Arid Lands Newsletter. Tucson, AZ: The University of Arizona, Office of Arid Lands Studies. 31: 18-23. [17350]
11. Anderson, M. Kat. 1999. The fire, pruning, and coppice management of temperate ecosystems for basketry material by California Indian tribes. Human Ecology. 27(1): 79-113. [35820]
12. Arno, Stephen F. 2000. Fire in western forest ecosystems. In: Brown, James K.; Smith, Jane Kapler, eds. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42-vol. 2. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 97-120. [36984]
13. Arno, Stephen F.; Hammerly, Ramona P. 1977. Northwest trees. Seattle, WA: The Mountaineers. 222 p. [4208]
14. Arno, Stephen F.; Harrington, Michael G. 1998. The Interior West: managing fire-dependent forests by simulating natural disturbance regimes. In: Proceedings: Forest management into the next century: what will make it work; 1997 November 19-21; Spokane, WA. Madison, WI: Forest Products Society: 53-62. [43185]
15. Atzet, Thomas; Wheeler, David L. 1982. Historical and ecological perspectives on fire activity in the Klamath Geological Province of the Rogue River and Siskiyou National Forests. R6-Range-102-1982. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 16 p. [6252]
16. Azuma, David L.; Donnegan, Joseph; Gedney, Donald. 2004. Southwest Oregon Biscuit Fire: an analysis of forest resources and fire severity. Res. Pap. PNW-RP-560. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 32 p. [50121]
17. Bailey, Arthur W.; Poulton, Charles E. 1968. Plant communities and environmental interrelationships in a portion of the Tillamook Burn, northwestern Oregon. Ecology. 49(1): 1-13. [6232]
18. Baker, Frederick S. 1949. A revised tolerance table. Journal of Forestry. 47: 179-181. [20405]
19. Balian, Estelle V.; Naiman, Robert J. 2005. Abundance and production of riparian trees in the lowland floodplain of the Queets River, Washington. Ecosystems. 8(7): 841-861. [83177]
20. Barbour, Michael G. 1988. Californian upland forests and woodlands. In: Barbour, Michael G.; Billings, William Dwight, eds. North American terrestrial vegetation. New York: Cambridge University Press: 131-164. [13880]
21. Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. 2007. Terrestrial vegetation of California. 3rd ed. Berkeley, CA: University of California Press. 712 p. [82605]
22. Barry, W. James. 1988. Some uses of riparian species in the landscape and for revegetation. In: Rieger, John P.; Williams, Bradford K., eds. Proceedings of the second native plant revegetation symposium; 1987 April 15-18; San Diego, CA. Madison, WI: University of Wisconsin Arboretum; Society for Ecological Restoration & Management: 164-168. [4111]
23. Beche, Leah A.; Stephens, Scott L.; Resh, Vincent H. 2005. Effects of prescribed fire on a Sierra Nevada (California, USA) stream and its riparian zone. Forest Ecology and Management. 218(1-3): 37-59. [55791]
24. Beschta, Robert L.; Ripple, William J. 2009. Large predators and trophic cascades in terrestrial ecosystems of the western United States. Biological Conservation. 142(11): 2401-2414. [77264]
25. Bolsinger, Charles L.; Waddell, Karen L. 1993. Area of old-growth forests in California, Oregon, and Washington. Resource Bulletin PNW-RB-197. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 26 p. [24150]
26. Bolton, Robert B., Jr.; Vogl, Richard J. 1969. Ecological requirements of Pseudotsuga macrocarpa in the Santa Ana Mountains, California. Journal of Forestry. 67: 112-116. [10807]
27. Borchert, Mark; Segotta, Daniel; Purser, Michael D. 1988. Coast redwood ecological types of southern Monterey County, California. Gen. Tech. Rep. PSW-107. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 27 p. [10225]
28. Bowler, Peter A. 1990. Riparian woodland: an endangered habitat in southern California. In: Schoenherr, Allan A., ed. Endangered plant communities of southern California: Proceedings, 15th annual symposium; 1989 October 28; Fullerton, CA. Special Publication No. 3. Claremont, CA: Southern California Botanists: 80-97. [21321]
29. Boyd, Jr., Raymond J. 1980. Grand fir. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 94. [50029]
30. Boyd, Robert. 1999. Strategies of Indian burning in the Willamette Valley. In: Boyd, Robert, ed. Indians, fire and the land in the Pacific Northwest. Corvallis, OR: Oregon State University Press: 94-138. [35572]
31. Brink, V. C. 1954. Survival of plants under flood in the lower Fraser River valley, British Columbia. Ecology. 35(1): 94-95. [64483]
32. Carey, Andrew B. 1996. Interactions of Northwest forest canopies and arboreal mammals. Northwest Science. 70: 72-78. [26799]
33. Carey, Andrew B. 2000. Ecology of northern flying squirrels: implications for ecosystem management in the Pacific Northwest, USA. In: Goldingay, Ross L.; Scheibe, John S., eds. Biology of gliding mammals. Firth, Germany: Filander Verlag: 45-63. [47416]
34. Carlson, Jack R. 1992. Selection, production, and use of riparian plant materials for the western United States. In: Landis, Thomas D., technical coordinator. Proceedings, Intermountain Forest Nursery Association; 1991 August 12-16; Park City, UT. Gen. Tech. Rep. RM-211. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 55-67. [20926]
35. Cassirer, E. Francis; Schirato, Greg; Sharpe, Fred; Groves, Craig R.; Anderson, Rusty N. 1993. Cavity nesting by harlequin ducks in the Pacific Northwest. The Wilson Bulletin. 105(4): 691-694. [22778]
36. Chesnut, V. K. 1902. Plants used by the Indians of Mendocino County, California. Contributions from the U.S. National Herbarium. [Washington, DC]: U.S. Department of Agriculture, Division of Botany. 7(3): 295-408. [54917]
37. Clement, C. J. E. 1985. Floodplain succession on the west coast of Vancouver Island. The Canadian Field-Naturalist. 99(1): 34-39. [8928]
38. Cline, Steven P.; Phillips, Charles A. 1983. Coarse woody debris and debris-dependent wildlife in logged and natural riparian zone forests--a western Oregon example. In: Davis, Jerry W.; Goodwin, Gregory A.; Ockenfeis, Richard A., technical coordinators. Snag habitat management: proceedings of the symposium; 1983 June 7-9; Flagstaff, AZ. Gen. Tech. Rep. RM-99. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 33-39. [17816]
39. Cole, Elizabeth C.; McComb, William C.; Newton, Michael; Chambers, Carol L.; Leeming, J. P. 1999. Response of small mammal and amphibian capture rates to clearcutting, burning, and glyphosate application in the Oregon Coast Range. In: Healthy forests for the 21st century: new technologies in integrated vegetation management: Proceedings, 20th annual forest vegetation conference; 1999 January 19-21; Redding, CA. Redding, CA: Forest Vegetation Conference: 103-105. [40672]
40. Collingwood, G. H.; Brush, Warren D. 1964. Knowing your trees. 2nd ed. [Revised edition edited by Devereux Butcher]. Washington, DC: The American Forestry Association. 349 p. [22497]
41. Collins, Brian D.; Montgomery, David R. 2002. Forest development, wood jams, and restoration of floodplain rivers in the Puget Lowland, Washington. Restoration Ecology. 10(2): 237-247. [44625]
42. Cook, John G. 2002. Nutrition and food. In: Toweill, Dale E.; Thomas, Jack Ward, eds. North American elk: ecology and management. Washington, DC: Smithsonian Institution Press: 259-350. [81796]
43. Cooper, William Skinner. 1922. The broad-sclerophyll vegetation of California: An ecological study of the chaparral and its related communities. Publ. No. 319. Washington, DC: The Carnegie Institution of Washington. 145 p. [6716]
44. Crockett, R. P.; Alber, B. 1992. Glyphosate and imazapyr combinations for big leaf maple resprout control in Pacific Northwest conifer production. In: Lym, Rodney G., ed. Proceedings, Western Society of Weed Science; 1992 March 10-12; Salt Lake City, UT. In: Proceedings, Western Society of Weed Science; 45: 72. [20611]
45. Crouch, Glenn L. 1968. Clipping of woody plants by mountain beaver. Journal of Mammalogy. 49(1): 151-152. [64410]
46. Curtis, Alan B. 2003. Horse Rock Ridge Research Natural Area: Guidebook Supplement 27. Gen. Tech. Rep. PNW-GTR-571. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 30 p. [45385]
47. Dale, Virginia H.; Hemstrom, Miles A.; Franklin, Jerry F. 1984. The effect of disturbance frequency on forest succession in the Pacific Northwest. In: New forests for a changing world: Proceedings of the 1983 convention of the Society of American Foresters; 1983 October 16-20; Portland, OR. Bethesda, MD: Society of American Foresters: 300-304. [4781]
48. Dale, Virginia H.; Hemstrom, Miles; Franklin, Jerry. 1986. Modeling the long-term effects of disturbances on forest succession, Olympic Peninsula, Washington. Canadian Journal of Forest Research. 16: 56-57. [4785]
49. Dayton, William A. 1931. Important western browse plants. Misc. Publ. No. 101. Washington, DC: U.S. Department of Agriculture. 214 p. [768]
50. Dewberry, Charley. 1990. Burning issues: fire and the western Oregon landscape. Eugene, OR: University of Oregon, Museum of Natural History. 11 p. [11756]
51. Dyrness, C. T. 1965. The effect of logging and slash burning on understory vegetation in the H. J. Andrews Experimental Forest. Res. Note PNW-31. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 13 p. [4939]
52. Dyrness, C. T. 1973. Early stages of plant succession following logging and burning in the western Cascades of Oregon. Ecology. 54(1): 57-69. [7345]
53. Dyrness, C. T.; Franklin, J. F.; Moir, W. H. 1974. A preliminary classification of forest communities in the central portion of the western Cascades in Oregon. Bulletin No. 4. Seattle, WA: University of Washington, Ecosystem Analysis Studies, Coniferous Forest Biome. 123 p. [8480]
54. Eis, S.; Craigdallie, D. 1980. Gulf Islands of British Columbia, a landscape analysis. BC-X-216. Victoria, BC: Environment Canada, Canadian Forestry Service, Pacific Forest Research Centre. 39. [18774]
55. Eis, S.; Craigdallie, D. 1980. Shore and landscape analysis of the western section of the Capital Regional District of British Columbia. BC-X-208. Victoria, BC: Canadian Forestry Service, Pacific Forest Research Centre. 43 p. [18680]
56. English, Pennoyer F. 1923. The dusky-footed wood rat (Neotoma fuscipes). Journal of Mammalogy. 4(1): 1-9. [65455]
57. Fierke, Melissa K.; Kauffman, J. Boone. 2005. Structural dynamics of riparian forests along a black cottonwood successional gradient. Forest Ecology and Management. 215(1-3): 149-162. [55572]
58. Figueroa, Paul F.; Nishimura, Thomas E. 1992. Bigleaf maple control: basal thin-line applications using imazapyr liquids and ground applications of imazapyr granules. In: Lym, Rodney G., ed. Proceedings, Western Society of Weed Science; 1992 March 10-12; Salt Lake City, UT. In: Western Society of Weed Science; 45: 65-72. [20610]
59. Fites-Kaufman, Joann; Bradley, Anne F.; Merrill, Amy G. 2006. Fire and plant interactions. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 94-117. [65534]
60. Fonda, R. W. 1974. Forest succession in relation to river terrace development in Olympic National Park, Washington. Ecology. 55(5): 927-942. [6746]
61. Fonda, R. W. 2001. Postfire response of red alder, black cottonwood, and bigleaf maple to the Whatcom Creek fire, Bellingham, Washington. Northwest Science. 75(1): 25-36. [38964]
62. Franklin, Jerry F. 1988. Pacific Northwest forests. In: Barbour, Michael G.; Billings, William Dwight, eds. North American terrestrial vegetation. New York: Cambridge University Press: 103-130. [13879]
63. Franklin, Jerry F.; Dyrness, C. T. 1973. Natural vegetation of Oregon and Washington. Gen. Tech. Rep. PNW-8. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 417 p. [961]
64. Fried, Jeremy S. 1987. Big leaf maple in Douglas-fir forests: effects on soils; seedling establishment and early growth. In: Plumb, Timothy R.; Pillsbury, Norman H., technical coordinators. Proceedings of the symposium on multiple-use management of California's hardwood resources; 1986 November 12-14; San Luis Obispo, CA. Gen. Tech. Rep. PSW-100. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 435. [5391]
65. Fried, Jeremy S.; Tappeiner, John C., II; Hibbs, David E. 1988. Bigleaf maple seedling establishment and early growth in Douglas-fir forests. Canadian Journal of Forest Research. 18: 1226-1233. [6189]
66. Fried, Jeremy S; Boyle, James R.; Tappeiner, John C., II; Cromack, Kermit, Jr. 1990. Effects of bigleaf maple on soils in Douglas-fir forests. Canadian Journal of Forest Research. 20: 259-266. [11033]
67. Frost, Evan J.; Sweeney, Rob. 2000. Fire regimes, fire history and forest conditions in the Klamath-Siskiyou region: an overview and synthesis of knowledge, [Online]. In: Klamath Siskiyou Wildland Center--Fire ecology and policy. Ashland, OR: Wildwood Environmental Consulting (Producer). Available: http://kswild.org/fire/fire_report.pdf [2011, November 15]. [69111]
68. Galatowitsch, S. M. 1990. Using the original land survey notes to reconstruct presettlement landscapes in the American West. The Great Basin Naturalist. 50(2): 181-191. [13772]
69. Garbelotto, Matteo; Davidson, Jennifer M.; Ivors, Kelly; Maloney, Patricia E.; Huberli, Daniel; Koike, Steven T.; Rizzo, David M. 2003. Non-oak native plants are main hosts for sudden oak death pathogen in California. California Agriculture. 57(1): 18-23. [50299]
70. Garman, Steven L.; Acker, Steven A.; Ohmann, Janet L.; Spies, Thomas A. 1995. Asymptotic height-diameter equations for twenty-four tree species in western Oregon. Research Contribution 10. Corvallis, OR: Oregon State University, College of Forestry, Forest Research Laboratory. 22 p. [65706]
71. Gilbert, Frederick F.; Allwine, Rochelle. 1991. Spring bird communities in the Oregon Cascade Range. In: Ruggiero, Leonard F.; Aubry, Keith B.; Carey, Andrew B.; Huff, Mark H., technical coordinators. Wildlife and vegetation of unmanaged Douglas-fir forests. Gen. Tech. Rep. PNW-GTR-285. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 145-158. [17311]
72. Giusti, Gregory A. 2007. Structural characteristics of an old-growth coast redwood stand in Mendocino County, California. In: Standiford, Richard B.; Giusti, Gregory A.; Valachovic, Yana; Zielinski, William J.; Furniss, Michale J., tech. eds. Proceedings of the redwood region forest science symposium: What does the future hold; 15-17 March 2004; Rohnert Park, CA. Gen. Tech. Rep. RSW-GTR-194. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 161-168. [71147]
73. Gleiser, Gabriela; Verdu, Miguel. 2005. Repeated evolution of dioecy from androdioecy in Acer. New Phytologist. 165(2): 633-640. [83794]
74. Goldner, Bernard H. 1984. Riparian restoration efforts associated with structurally modified flood control channels. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of the conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 445-451. [5852]
75. Gratkowski, H. 1974. Brushfield reclamation and type conversion. In: Cramer, Owen P., ed. Environmental effects of forest residues management in the Pacific Northwest: A state-of-knowledge compendium. Gen. Tech. Rep. PNW-24. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: I-1 to I-31. [6418]
76. Gray, Andrew N.; Franklin, Jerry F. 1997. Effects of multiple fires on the structure of southwestern Washington forests. Northwest Science. 71(3): 174-185. [27793]
77. Gray, Andrew N.; Monleon, Vicente J.; Spies, Thomas A. 2009. Characteristics of remnant old-growth forests in the northern Coast Range of Oregon and comparison to surrounding landscapes. Gen Tech. Rep. PNW-GTR-790. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 45 p. [81007]
78. Greene, D. F.; Johnson, E. A. 1993. Seed mass and dispersal capacity in wind-dispersed diaspores. Oikos. 67: 69-74. [68013]
79. Grier, Charles C.; Logan, Robert S. 1977. Old-growth Pseudotsuga menziesii communities of a western Oregon watershed: biomass distribution and production budgets. Ecological Monographs. 47: 373-400. [8762]
80. Griffin, James R. 1974. Notes on environment, vegetation and flora: Hastings Natural History Reservation. Carmel Valley, CA: University of California, Hastings Natural History Reservation. Unpublished paper on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 90 p. [10531]
81. Griffin, James R.; Critchfield, William B. 1972. The distribution of forest trees in California. Res. Pap. PSW-82. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 118 p. [1041]
82. Habeck, J. R. 1962. Forest succession in Monmouth Township, Polk County, Oregon since 1850. Proceedings of the Montana Academy of Sciences. 21: 7-17. [9059]
83. Haeussler, S.; Coates, D. 1986. Autecological characteristics of selected species that compete with conifers in British Columbia: a literature review. Land Management Report No. 33. Victoria, BC: Ministry of Forests, Information Services Branch. 180 p. [1055]
84. Haeussler, S.; Coates, D.; Mather, J. 1990. Autecology of common plants in British Columbia: A literature review. Economic and Regional Development Agreement: FRDA Report 158. Victoria, BC: Forestry Canada, Pacific Forestry Centre; British Columbia Ministry of Forests, Research Branch. 272 p. [18033]
85. Halpern, Charles B.; Harmon, Mark E. 1983. Early plant succession on the Muddy River mudflow, Mount St. Helens, Washington. The American Midland Naturalist. 110(1): 97-106. [8870]
86. Hanes, Ted L. 1976. Vegetation types of the San Gabriel Mountains. In: Latting, June, ed. Symposium proceedings: plant communities of southern California; 1974 May 4; Fullerton, CA. Special Publication No. 2. Berkeley, CA: California Native Plant Society: 65-76. [4227]
87. Hann, David W. 1997. Equations for predicting the largest crown width of stand-grown trees in western Oregon. Research Contribution 17. Corvallis, OR: Oregon State University, College of Forestry, Forest Research Laboratory. 14 p. [65649]
88. Hann, David W.; Larsen, David R. 1991. Diameter growth equations for fourteen tree species in southwest Oregon. Res. Bull. 69. Corvallis, OR: Oregon State University, College of Forestry, Forest Research Laboratory. 18 p. [65650]
89. Hann, Wendel; Havlina, Doug; Shlisky, Ayn; [and others]. 2010. Interagency fire regime condition class (FRCC) guidebook, [Online]. Version 3.0. In: FRAMES (Fire Research and Management Exchange System). National Interagency Fuels, Fire & Vegetation Technology Transfer (NIFTT) (Producer). Available: http://www.fire.org/niftt/released/FRCC_Guidebook_2010_final.pdf. [81749]
90. Harris, A. S. 1980. Western hemlock. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 102-103. [50038]
91. Harris, Roger D. 1983. Decay characteristics of pileated woodpecker nest trees. In: Davis, Jerry W.; Goodwin, Gregory A.; Ockenfeis, Richard A., technical coordinators. Snag habitat management: proceedings of the symposium; 1983 June 7-9; Flagstaff, AZ. Gen. Tech. Rep. RM-99. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 125-129. [17826]
92. Hartwig, C. L.; Eastman, D. S.; Harestad, A. S. 2004. Characteristics of pileated woodpecker (Dryocopus pileatus) cavity trees and their patchiness on southeastern Vancouver Island, British Columbia, Canada. Forest Ecology and Management. 187: 225-234. [47058]
93. Hawk, G. M.; Zobel, D. B. 1974. Forest succession on alluvial landforms of the McKenzie River Valley, Oregon. Northwest Science. 48(4): 245-265. [9686]
94. Hawkes, B. C.; Feller, M. C.; Meehan, D. 1990. Site preparation: fire. In: Lavender, D. P.; Parish, R.; Johnson, C. M.; Montgomery, G.; Vyse, A.; Willis, R. A.; Winston, D., eds. Regenerating British Columbia's forests. Vancouver, BC: University of British Columbia Press: 131-149. [10712]
95. Heavilin, Danny. 1977. Conifer regeneration on burned and unburned clearcuts on granitic soils of the Klamath National Forest. Res. Note PSW-321. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 3 p. [4981]
96. Hemstrom, Miles A.; Logan, Sheila E. 1986. Plant association and management guide: Siuslaw National Forest. R6-Ecol 220-1986a. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 121 p. [10321]
97. Hemstrom, Miles A.; Logan, Sheila E.; Pavlat, Warren. 1987. Plant association and management guide: Willamette National Forest. R6-Ecol 257-B-86. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 312 p. [13402]
98. Henderson, Jan A.; Peter, David H.; Lesher, Robin D.; Shaw, David C. 1989. Forested plant associations of the Olympic National Forest. R6-ECOL-TP 001-88. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 502 p. [23405]
99. Hickman, James C., ed. 1993. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 1400 p. [21992]
100. Hines, William W. 1973. Black-tailed deer populations and Douglas-fir reforestation in the Tillamook Burn, Oregon. Game Research Report No. 3. Final report: Federal Aid to Wildlife Restoration--Project W-51-R. Corvallis, OR: Oregon State Game Commission, Research Division. 59 p. [8431]
101. Hines, William W.; Land, Charles E. 1974. Black-tailed deer and Douglas-fir regeneration in the Coast Range of Oregon. In: Black, Hugh C., ed. Wildlife and forest management in the Pacific Northwest: Proceedings of a symposium; 1973 September 11-12; Corvallis, OR. Corvallis, OR: Oregon State University, School of Forestry, Forest Research Laboratory: 121-132. [7999]
102. Hitchcock, C. Leo; Cronquist, Arthur. 1973. Flora of the Pacific Northwest. Seattle, WA: University of Washington Press. 730 p. [1168]
103. Holland, Robert F. 1986. Preliminary descriptions of the terrestrial natural communities of California. Sacramento, CA: California Department of Fish and Game. 156 p. [12756]
104. Horton, Jerome S. 1960. Vegetation types of the San Bernardino Mountains. Tech. Pap. No. 44. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 29 p. [10687]
105. Hosie, R. C. 1969. Native trees of Canada. 7th ed. Ottawa, ON: Canadian Forestry Service, Department of Fisheries and Forestry. 380 p. [3375]
106. Huff, Mark H.; Agee, James K. 1983. Fire effects on flora, fuels, and fauna in the western hemlock - Douglas-fir forest type. Technical Completion Report 3/NPS Contract CX-9000-9-E079: Ecological effects of the Hoh Fire. Seattle, WA: University of Washington, College of Forest Resources, NPS Cooperative Park Studies Unit. 5 p. [+ appendices]. [18555]
107. Huff, Mark H.; Raley, Cathrine M. 1991. Regional patterns of diurnal breeding bird communities in Oregon and Washington. In: Ruggiero, Leonard F.; Aubry, Keith B.; Carey, Andrew B.; Huff, Mark H., technical coordinators. Wildlife and vegetation of unmanaged Douglas-fir forests. Gen. Tech. Rep. PNW-GTR-285. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 177-205. [17313]
108. Hyatt, Timothy L.; Naiman, Robert J. 2001. The residence time of large woody debris in the Queets River, Washington, USA. Ecological Applications. 11(1): 191-202. [83178]
109. Iddrisu, Mohammed N.; Ritland, Kermit. 2004. Genetic variation, population structure, and mating system in bigleaf maple (Acer macrophyllum Pursh). Canadian Journal of botany. 82(12): 1817-1825. [41794]
110. Ince, Peter J. 1979. How to estimate recoverable heat energy in wood or bark fuels. Gen. Tech. Rep. FPL 29. Madison, WI: U.S. Department of Agriculture, Forest Service, Forest Products Laboratory. 7 p. [13241]
111. Jameson, E. W., Jr. 1952. Food of deer mice, Peromyscus maniculatus and P. boylei, in the northern Sierra Nevada, California. Journal of Mammalogy. 33(1): 50-60. [21605]
112. Johnson, David H.; O'Neil, Thomas A., eds. 2001. Wildlife-habitat relationships in Oregon and Washington. Corvallis, OR: Oregon State University. 736 p. [65053]
113. Julian-Gordon, A. Lynn. 1993. The sex life of the big-leaf maple: Acer macrophyllum Pursh. Corvallis, OR: University of Oregon. 257 p. Thesis. [83182]
114. Junak, Steve; Knapp, Denise A.; Haller, J. Robert; Philbrick, Ralph; Schoenherr, Allan; Keeler-Wolf, Todd. 2007. The California Channel Islands. In: Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. Terrestrial vegetation of California. Berkeley, CA: University of California Press: 229-252. [82702]
115. Kartesz, John T. 1999. A synonymized checklist and atlas with biological attributes for the vascular flora of the United States, Canada, and Greenland. 1st ed. In: Kartesz, John T.; Meacham, Christopher A. Synthesis of the North American flora (Windows Version 1.0), [CD-ROM]. Chapel Hill, NC: North Carolina Botanical Garden (Producer). In cooperation with: The Nature Conservancy; U.S. Department of Agriculture, Natural Resources Conservation Service; U.S. Department of the Interior, Fish and Wildlife Service. [36715]
116. Kellman, M. C. 1969. Plant species interrelationships in a secondary succession in coastal British Columbia. Syesis. 2: 201-212. [6589]
117. Kennedy, Rebecca S. H.; Spies, Thomas A. 2005. Dynamics of hardwood patches in a conifer matrix: 54 years of change in a forested landscape in coastal Oregon, USA. Biological Conservation. 122(3): 363-374. [50866]
118. King, David A. 1991. Tree allometry, leaf size and adult tree size in old-growth forests of western Oregon. Tree Physiology. 9(3): 369-381. [48473]
119. Klinka, K.; Krajina, V. J.; Ceska, A.; Scagel, A. M. 1989. Indicator plants of coastal British Columbia. Vancouver, BC: University of British Columbia Press. 288 p. [10703]
120. Knowe, Steven A.; Carrier, Byron D.; Dobkowski, Alex. 1995. Effects of bigleaf maple sprout clumps on diameter and height growth of Douglas-fir. Western Journal of Applied Forestry. 10(1): 5-11. [25493]
121. Krajina, V. J.; Klinka, K.; Worrall, J. 1982. Distribution and ecological characteristics of trees and shrubs of British Columbia. Vancouver, BC: University of British Columbia. 131 p. [6728]
122. Lamberti, Gary A.; Gregory, Stan V.; Ashkenas, Linda R.; Wildman, Randall C.; Steinman, Alan D. 1989. Influence of channel geomorphology on retention of dissolved and particulate matter in a Cascade Mountain stream. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 33-39. [13510]
123. LANDFIRE Rapid Assessment. 2005. Potential Natural Vegetation Group (PNVG) R#SSHE--Sitka Spruce - Hemlock, [Online]. In: Rapid assessment reference condition models. In: LANDFIRE. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory; U.S. Geological Survey; The Nature Conservancy (Producers). Available: http://www.landfire.gov/zip/PNW/RSSHE_Aug08.pdf [2011, October 27]. [83815]
124. LANDFIRE Rapid Assessment. 2005. Potential Natural Vegetation Group (PNVG) R1CHAP--Chaparral, [Online]. In: Rapid assessment reference condition models. In: LANDFIRE. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory; U.S. Geological Survey; The Nature Conservancy (Producers). Available: http://www.landfire.gov/zip/CA/R1CHAP_Aug08.pdf [2011, October 27]. [83823]
125. LANDFIRE Rapid Assessment. 2005. Potential Natural Vegetation Group (PNVG) R1CHAPm, montane chaparral [Online]. In: Rapid assessment reference condition models. In: LANDFIRE. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory; U.S. Geological Survey; The Nature Conservancy (Producers). Available: http://www.landfire.gov/zip/CA/R1CHAPmn_Aug08.pdf [2011, October 31]. [83867]
126. LANDFIRE Rapid Assessment. 2005. Potential Natural Vegetation Group (PNVG) R1SAGEco--Coastal sage scrub, [Online]. In: Rapid assessment reference condition models. In: LANDFIRE. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory; U.S. Geological Survey; The Nature Conservancy (Producers). Available: http://www.landfire.gov/zip/CA/R1SAGEco_Aug08.pdf [2011, October 27]. [83824]
127. LANDFIRE Rapid Assessment. 2005. Reference condition modeling manual (Version 2.1), [Online]. In: LANDFIRE. Cooperative Agreement 04-CA-11132543-189. Boulder, CO: The Nature Conservancy; U.S. Department of Agriculture, Forest Service; U.S. Department of the Interior (Producers). 72 p. Available: http://www.landfire.gov/downloadfile.php?file=RA_Modeling_Manual_v2_1.pdf [2007, May 24]. [66741]
128. LANDFIRE Rapid Assessment. 2007. Rapid assessment reference condition models, [Online]. In: LANDFIRE. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Lab; U.S. Geological Survey; The Nature Conservancy (Producers). Available: http://www.landfire.gov/models_EW.php [2008, April 18] [66533]
129. Larsen, David R.; Hann, David W. 1987. Height-diameter equations for seventeen tree species in southwest Oregon. Research Paper 49. Corvallis, OR: Oregon State University, College of Forestry, Forest Research Lab. 16 p. [49458]
130. Larson, Bruce C.; Oliver, Chadwick Dearing. 1982. Forest dynamics and fuelwood supply of the Stehekin Valley, Washington. In: Means, Joseph E., ed. Forest succession and stand development research in the Pacific Northwest: Proceedings of the symposium; 1981 March 26; Corvallis, OR. Corvallis, OR: Oregon State University, Forest Research Laboratory: 127-134. [78734]
131. Lauterbach, Paul; Warren, L. E. 1982. Control of resprouting hardwood clumps with applications of triclopyr ester by hovering helicopter. In: Proceedings, Western Society of Weed Science; 1982 March 9-11; Denver, CO. In: Proceedings, Western Weed Science Society. 35: 36-38. [67927]
132. Lei, T. T.; Lechowicz, M. J. 1997. The photosynthetic response of eight species of Acer to simulated light regimes from the centre and edges of gaps. Functional Ecology. 11(1): 16-23. [83795]
133. Lillybridge, Terry R.; Kovalchik, Bernard L.; Williams, Clinton K.; Smith, Bradley G. 1995. Field guide for forested plant associations of the Wenatchee National Forest. Gen. Tech. Rep. PNW-GTR-359. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 335 p. In cooperation with: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region, Wenatchee National Forest. [29851]
134. Lininger, Jay C. 2004. Fire history and need for fuel management in mixed Douglas-fir forests of the Klamath-Siskiyou region, northwest California and southwest Oregon, USA. In: 2nd international wildland fire ecology and fire management congress; 5th symposium on fire and forest meteorology: Proceedings; 2003 November 16-20; Orlando, FL. Boston, MA: American Meteorological Society: 1-18. [64193]
135. Lutz, J. A.; van Wagtendonk, J. W.; Franklin, J. F. 2009. Twentieth-century decline of large-diameter trees in Yosemite National Park, California, USA. Forest Ecology and Management. 257(11): 2296-2307. [74287]
136. Lutz, James A.; Halpern, Charles B. 2006. Tree mortality during early forest development: a long-term study of rates, causes, and consequences. Ecological Monographs. 76(2): 257-275. [83796]
137. Mallory, James I. 1980. Canyon live oak. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 125-126. [7608]
138. Mason, Herbert L. 1957. A flora of the marshes of California. Berkeley, CA: University of California Press. 878 p. [16905]
139. McCain, Cindy; Christy, John A. 2005. Field guide to riparian plant communities in northwestern Oregon. Tech. Pap. R6-NR-ECOL-TP-01-05. [Portland, OR]: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 357 p. [63114]
140. McComb, William C. 2001. Management of within-stand forest habitat features. In: Johnson, David H.; O'Neil, Thomas A., managing directors. Wildlife-habitat relationships in Oregon and Washington. Corvallis, OR: Oregon State University Press: 140-153. [82880]
141. McComb, William C.; McGarigal, Kevin; Anthony, Robert G. 1993. Small mammal and amphibian abundance in streamside and upslope habitats of mature Douglas-fir stands, western Oregon. Northwest Science. 67(1): 7-15. [20564]
142. McDadi, Omar; Hebda, Richard J. 2008. Change in historic fire disturbance in a Garry oak (Quercus garryana) meadow and Douglas-fir (Pseudotsuga menziesii) mosaic, University of Victoria, British Columbia, Canada: a possible link with First Nations and Europeans. Forest Ecology and Management. 256(10): 1704-1710. [72957]
143. McDonald, Philip M. 1980. Pacific ponderosa pine--Douglas-fir. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 120. [50055]
144. McGee, Ann; Feller, M. C. 1993. Seed banks of forested and disturbed soils in southwestern British Columbia. Canadian Journal of Botany. 71: 1574-1583. [25756]
145. Miller, Frank L. 1968. Observed use of forage and plant communities by black-tailed deer. The Journal of Wildlife Management. 32(1): 142-148. [83797]
146. Miller, Melanie. 2000. Fire autecology. In: Brown, James K.; Smith, Jane Kapler, eds. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42-vol. 2. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 9-34. [36981]
147. Miller, Richard E. 1980. Red alder. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 100. [50035]
148. Minnich, Richard A. 1976. Vegetation of the San Bernardino Mountains. In: Latting, June, ed. Symposium proceedings: Plant communities of southern California; 1974 May 4; Fullerton, CA. Special Publication No. 2. Berkeley, CA: California Native Plant Society: 99-124. [4232]
149. Minore, Don. 1978. The Dead Indian Plateau: a historical summary of forestry observations and research in a severe southwestern Oregon environment. PNW-72. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 23 p. [8423]
150. Minore, Don. 1980. Western hemlock-Sitka spruce. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 103. [50040]
151. Minore, Don; Zasada, John C. 1990. Acer macrophyllum Pursh bigleaf maple. In: Burns, Russell M.; Honkala, Barbara H., technical coordinators. Silvics of North America. Volume 2. Hardwoods. Agric. Handb. 654. Washington, DC: U.S. Department of Agriculture, Forest Service: 33-40. [83260]
152. Mirov, N. T.; Kraebel, C. J. 1937. Collecting and propagating the seeds of California wild plants. Res. Note No. 18. Berkeley, CA: U.S. Department of Agriculture, Forest Service, California Forest and Range Experiment Station. 27 p. [9787]
153. Morris, William G. 1958. Influence of slash burning on regeneration, other plant cover, and fire hazard in the Douglas-fir region: A progress report. Res. Pap. PNW-29. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 49 p. [4803]
154. Norris, L. A.; Lorz, H. W.; Gregory, S. V. 1983. Forest chemicals. Gen. Tech. Rep. PNW-149. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 95 p. (Meehan, William R., tech. ed.; Influence of forest and rangeland management on anadromous fish habitat in western North America; pt. 9). [13658]
155. Norris, L. A.; Montgomery, M. L.; Warren, L. E.; Mosher, W. D. 1982. Brush control with herbicides on hill pasture sites in southern Oregon. Journal of Range Management. 35(1): 75-80. [7872]
156. O'Geen, Anthony T.; Dahlgren, Randy A.; Sanchez-Mata, Daniel. 2007. California soils and examples of ultramafic vegetation. In: Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. Terrestrial vegetation of California. 3rd ed. Berkeley, CA: University of California Press: 71-106. [82692]
157. O'Neil, Thomas A.; Johnson, David H. 2001. Oregon and Washington wildlife species and their habitats. In: Johnson, David H.; O'Neil, Thomas A., managing directors. Wildlife-habitat relationships in Oregon and Washington. Corvallis, OR: Oregon State University Press: 1-21. [82878]
158. Pabst, Robert J.; Spies, Thomas A. 1999. Structure and composition of unmanaged riparian forests in the coastal mountains of Oregon, U.S.A. Canadian Journal of Forest Research. 29: 1557-1573. [36366]
159. Parminter, John. 1991. Fire history and effects on vegetation in three biogeoclimatic zones of British Columbia. In: Nodvin, Stephen C.; Waldrop, Thomas A., eds. Fire and the environment: ecological and cultural perspectives: Proceedings of an international symposium; 1990 March 20-24; Knoxville, TN. Gen. Tech. Rep. SE-69. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: 263-272. [16651]
160. Paysen, Timothy E.; Derby, Jeanine A.; Black, Hugh, Jr.; Bleich, Vernon C.; Mincks, John W. 1980. A vegetation classification system applied to southern California. Gen. Tech. Rep. PSW-45. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 33 p. [1849]
161. Peterson, David L.; Hammer, R. David. 2001. From open to closed canopy: a century of change in a Douglas-fir forest, Orcas Island, Washington. Northwest Science. 75(3): 262-269. [40208]
162. Peterson, E. B.; Peterson, N. M.; Comeau, P. G.; Thomas, K. D. 1999. Bigleaf maple manager's handbook for British Columbia. Hardwood and Vegetation Management Technical Advisory Committee Series. Victoria, BC: Ministry of Forests, Forestry Division Services Branch. 105 p. [83798]
163. Pojar, J.; Meidinger, D. 1991. British Columbia: The environmental setting. In: Meidinger, Del; Pojar, Jim, eds. Ecosystems of British Columbia. Special Report Series 6. Victoria, BC: British Columbia Ministry of Forests: 39-67. [83866]
164. Pojar, Jim; MacKinnon, Andy, eds. 1994. Plants of the Pacific Northwest coast: Washington, Oregon, British Columbia and Alaska. Redmond, WA: Lone Pine Publishing. 526 p. [25159]
165. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press. 632 p. [2843]
166. Reukema, Donald L. 1980. Douglas-fir--western hemlock. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 107-108. [50045]
167. Rickard, W. H. 1975. Litterfall in a Douglas-fir forest near the Trojan Nuclear Power Station, Oregon. Northwest Science. 49(4): 183-189. [8178]
168. Rizzo, David M.; Garbelotto, Matteo; Davidson, Jennifer M.; Slaughter, Garey W.; Koike, Steven T. 2002. Phytophthora ramorum and sudden oak death in California: I. Host relationships. In: Standiford, Richard B.; McCreary, Douglas; Purcell, Kathryn L., tech. coords. Proceedings of the 5th symposium on oak woodlands: oaks in California's changing landscape; 2001 October 22-25; San Diego, CA. Gen. Tech. Rep. PSW-GTR-184. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 733-740. [42368]
169. Roberts, Catherine Anne. 1975. Initial plant succession after brown and burn site preparation on an alder-dominated brushfield in the Oregon Coast Range. Corvallis, OR: Oregon State University. 90 p. Thesis. [9786]
170. Robinson, Cyril S. 1937. Plants eaten by California mule deer on the Los Padres National Forest. Journal of Forestry. 35(3): 285-292. [51853]
171. Roy, D. F. 1955. Hardwood sprout measurements in northwestern California. Forest Research Notes No. 95. Berkeley, CA: U.S. Department of Agriculture, Forest Service, California Forest and Range Experiment Station. 6 p. [8999]
172. Roy, Douglas F. 1980. Redwood. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 109-110. [50047]
173. Rundel, Philip W. 1987. Origins and adaptations of California hardwoods. In: Plumb, Timothy R.; Pillsbury, Norman H., technical coordinators. Proceedings of the symposium on multiple-use management of California's hardwood resources; 1986 November 12-14; San Luis Obispo, CA. Gen. Tech. Rep. PSW-100. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 11-17. [5355]
174. Ruth, Robert H.; Underwood, J. Clyde; Smith, Clark E.; Yang, Hoya Y. 1972. Maple sirup production from bigleaf maple. PNW-181. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 12 p. [8592]
175. Sabhasri, Sanga; Ferrell, William K. 1960. Invasion of brush species into small stand openings in the Douglas-fir forests of the Willamette foothills. Northwest Science. 34(3): 77-89. [8652]
176. Safford, Hugh Deforest. 1995. Woody vegetation and succession in the Garin Woods, Hayward Hills, Alameda County, California. Madrono. 42(4): 470-489. [40868]
177. Sampson, Arthur W.; Jespersen, Beryl S. 1963. California range brushlands and browse plants. Berkeley, CA: University of California, Division of Agricultural Sciences; California Agricultural Experiment Station, Extension Service. 162 p. [3240]
178. Sauer, Jonathan D. 1977. Fire history, environmental patterns, and species patterns in Santa Monica mountain chaparral. In: Mooney, Harold A.; Conrad, C. Eugene, technical coordinators. Proceedings of the symposium of the environmental consequences of fire and fuel management in Mediterranean ecosystems; 1977 August 1-5; Palo Alto, CA. Gen. Tech. Rep. WO-3. Washington, DC: U.S. Department of Agriculture, Forest Service: 383-386. [4866]
179. Sawyer, John O. 2007. Forests of northwestern California. In: Barbour, Michael G.; Keeler-Wolf, Todd; Schoenherr, Allan A., eds. Terrestrial vegetation of California. Berkeley, CA: University of California Press: 253-295. [82703]
180. Schroer, Greg L.; Jenkins, Kurt J.; Moorhead, Bruce B. 1993. Roosevelt elk selection of temperate rain forest seral stages in western Washington. Northwest Science. 67(1): 23-29. [20563]
181. Sedell, James R.; Everest, Fred H.; Gibbons, David R. 1989. Streamside vegetation management for aquatic habitat. In: Proceedings of the national silviculture workshop; 1987 May 11-14; Sacramento, CA. Washington, DC: U.S. Department of Agriculture, Forest Service: 115-125. [6403]
182. Shaffer, Kevin E.; Laudenslayer, William F., Jr. 2006. Fire and animal interactions. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 118-144. [65536]
183. Sheridan, Chris D.; Spies, Thomas A. 2005. Vegetation-environment relationships in zero-order basins in coastal Oregon. Canadian Journal of Forest Research. 35: 340-355. [60361]
184. Skinner, Carl N.; Taylor, Alan H. 2006. Southern Cascades bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 195-224. [65540]
185. Skinner, Carl N.; Taylor, Alan H.; Agee, James K. 2006. Klamath Mountains bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 170-194. [65539]
186. Smith, J. Harry G. 1974. Dynamics of stand development as related to wildlife. In: Black, Hugh C., ed. Wildlife and forest management in the Pacific Northwest: Proceedings of a symposium; 1973 September 11-12; Corvallis, OR. Corvallis, OR: Oregon State University, School of Forestry, Forest Research Laboratory: 1-14. [7973]
187. Spenger, Connie. 1985. The northernmost Tecate cypress. Fremontia. 13(3): 8-10. [22532]
188. Spies, Thomas A. 1991. Plant species diversity and occurrence in young, mature, and old-growth Douglas-fir stands in western Oregon and Washington. In: Ruggiero, Leonard F.; Aubry, Keith B.; Carey, Andrew B.; Huff, Mark H., technical coordinators. Wildlife and vegetation of unmanaged Douglas-fir forests. Gen. Tech. Rep. PNW-GTR-285. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 111-121. [17309]
189. Spies, Thomas A.; Franklin, Jerry F. 1991. The structure of natural young, mature, and old-growth Douglas-fir forests in Oregon and Washington. In: Ruggiero, Leonard F.; Aubry, Keith B.; Carey, Andrew B.; Huff, Mark H., technical coordinators. Wildlife and vegetation of unmanaged Douglas-fir forests. Gen. Tech. Rep. PNW-GTR-285. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 91-109. [17308]
190. Stalmaster, Mark V.; Knight, Richard L.; Holder, Barbara L.; Anderson, Robert J. 1985. Bald eagles. In: Brown, E. Reade, tech. ed. Management of wildlife and fish habitats in forests of western Oregon and Washington: Part 1--Chapter narratives. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region: 269-290. In cooperation with: U.S. Department of the Interior, Bureau of Land Management. [74833]
191. Stein, William A. 1980. Port Orford-cedar. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 108-109. [50046]
192. Stein, William I. 1980. Oregon white oak. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 110-111. [9857]
193. Stein, William I. 1981. Regeneration outlook on BLM lands in the southern Oregon Cascades. Res. Pap. PNW-284. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 68 p. [5031]
194. Stickney, Peter F. 1989. Seral origin of species comprising secondary plant succession in Northern Rocky Mountain forests. FEIS workshop: Postfire regeneration. Unpublished draft on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 10 p. [20090]
195. Stuart, John D.; Stephens, Scott L. 2006. North Coast bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 147-169. [65538]
196. Stubblefield, Cynthia H. 1993. Food habits of black bear in the San Gabriel Mountains of southern California. The Southwestern Naturalist. 38(3): 290-293. [22146]
197. Sugihara, Neil G.; van Wagendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E. 2006. Fire in California's ecosystems. Berkeley, CA: University of California Press. 596 p. [65256]
198. Tappeiner, John C., II; Zasada, John; Huffman, David; Maxwell, Bruce D. 1996. Effects of cutting time, stump height, parent tree characteristics, and harvest variables on development of bigleaf maple sprout clumps. Western Journal of Applied Forestry. 11(4): 120-124. [83175]
199. Tappeiner, John C.; Zasada, John C. 1993. Establishment of salmonberry, salal, vine maple, and bigleaf maple seedlings in the coastal forests of Oregon. Canadian Journal of Forest Research. 23(9): 1775-1780. [83176]
200. Tarrant, Robert F.; Isaac, Leo A.; Chandler, Robert F., Jr. 1951. Observations on litter fall and foliage nutrient content of some Pacific northwest tree species. Journal of Forestry. 49: 914-915. [8179]
201. Taylor, Alan H.; Skinner, Carl N. 1998. Fire history and landscape dynamics in a late-successional reserve, Klamath Mountains, California, USA. Forest Ecology and Management. 111(2-3): 285-301. [30321]
202. Taylor, Alan H.; Skinner, Carl N. 2003. Spatial patterns and controls on historical fire regimes and forest structure in the Klamath Mountains. Ecological Applications. 13(3): 704-719. [52969]
203. Ter-Mikaelian, Michael T.; Korzukhin, Michael D. 1997. Biomass equations for sixty-five North American tree species. Forest Ecology and Management. 97: 1-24. [28034]
204. Thompson, Lisa C.; Voss, Jenna L.; Larsen, Royce, E.; Tietje, William D.; Cooper, Ryan A.; Moyle, Peter B. 2008. Role of hardwood in forming habitat for southern California steelhead. In: Merenlender, Adina; McCreary, Douglas; Purcell, Kathryn L., tech. eds. Proceedings of the 6th symposium on oak woodlands: today's challenges, tomorrow's opportunities; 2006 October 9-12; Rohnert Park, CA. Gen. Tech. Rep. PSW-GTR-217. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: 307-319. [80928]
205. Thompson, Robert S.; Anderson, Katherine H.; Bartlein, Patrick J. 1999. Digital representations of tree species range maps from "Atlas of United States trees" by Elbert L. Little, Jr. (and other publications), [Online]. In: Atlas of relations between climatic parameters and distributions of important trees and shrubs in North America--GIS files of tree species range maps. U.S. Geological Survey Professional Paper 1650 A&B. Reston, VA: U.S. Geological Survey, Geology and Environmental Change Science Center, Earth Surface Processes (Producer). Available: http://esp.cr.usgs.gov/data/atlas/little/ [2011, June 8]. [82831]
206. Thorne, Robert F. 1976. The vascular plant communities of California. In: Latting, June, ed. Symposium proceedings: plant communities of southern California; 1974 May 4; Fullerton, CA. Special Publication No. 2. Berkeley, CA: California Native Plant Society: 1-31. [3289]
207. Topik, Christopher. 1989. Plant association and management guide for the grand fir zone, Gifford Pinchot National Forest. R6-Ecol-TP-006-88. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Region. 110 p. [11361]
208. Trush, William J.; Connor, Edward C.; Knight, Allen W. 1989. Alder establishment and channel dynamics in a tributary of the South Fork Eel River, Mendocino County, California. In: Abell, Dana L., technical coordinator. Proceedings of the California riparian systems conference: Protection, management, and restoration for the 1990's; 1988 September 22-24; Davis, CA. Gen. Tech. Rep. PSW-110. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 14-21. [13509]
209. Turk, Tanya D.; Schmidt, Margaret G.; Roberts, Nicholas J. 2008. The influence of bigleaf maple on forest floor and mineral soil properties in a coniferous forest in coastal British Columbia. Forest Ecology and Management. 255(5-6): 1874-1882. [83793]
210. U.S. Department of Agriculture, Forest Service, Forest Products Laboratory. 1974. Wood handbook: wood as an engineering material. Agric. Handb. No. 72. Washington, DC. 415 p. [16826]
211. U.S. Department of Agriculture, Natural Resources Conservation Service. 2011. PLANTS Database, [Online]. Available: https://plants.usda.gov /. [34262]
212. Uzoh, Fabian C. C.; Richie, Martin W. 1996. Crown area equations for 13 species of trees and shrubs in northern California and southwestern Oregon. Res. Paper PSW-RP-227. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station. 13 p. [38453]
213. Valachovic, Y. S.; Caldwell, B. A.; Cromack, K., Jr.; Griffiths, R. P. 2004. Leaf litter chemistry controls on decomposition of Pacific Northwest trees and woody shrubs. Canadian Journal of Forest Research. 34: 2131-2147. [62192]
214. van Wagtendonk, Jan W.; Fites-Kaufman, Joann. 2006. Sierra Nevada bioregion. In: Sugihara, Neil G.; van Wagtendonk, Jan W.; Shaffer, Kevin E.; Fites-Kaufman, Joann; Thode, Andrea E., eds. Fire in California's ecosystems. Berkeley, CA: University of California Press: 264-294. [65544]
215. Wagner, Robert G.; Rogozynski, Michael W. 1994. Controlling sprout clumps of bigleaf maple with herbicides and manual cutting. Western Journal of Applied Forestry. 9(4): 118-124. [23936]
216. Wang, Chao-Huan; Hann, David W. 1988. Height-diameter equations for sixteen tree species in the central western Willamette Valley of Oregon. Research Paper 51. Corvallis, OR: Oregon State University, College of Forestry, Forest Research Lab. 7 p. [48627]
217. Waring, R. H.; Major, J. 1964. Some vegetation of the California coastal redwood region in relation to gradients of moisture, nutrients, light, and temperature. Ecological Monographs. 34: 167-215. [8924]
218. Weisberg, Peter J. 2009. Historical fire frequency on contrasting slope facets along the McKenzie River, western Oregon Cascades. Western North American Naturalist. 69(2): 206-217. [81463]
219. Whittaker, R. H. 1960. Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monographs. 30(3): 279-338. [6836]
220. Williamson, Richard L. 1980. Pacific Douglas-fir. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 106-107. [50044]
221. Wilson, James L.; Ayres, Debra R.; Steinmaus, Scott; Baad, Michael. 2009. Vegetation and flora of a biodiversity hotspot: Pine Hill, El Dorado County, California, USA. Madrono. 56(4): 246-278. [80220]
222. Wilson, Pamela L.; Funck, James W.; Avery, Robert B. 2010. Fuelwood characteristics of northwestern conifers and hardwoods. PNW-GTR-810 [Updated]. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 50 p. In cooperation with: Oregon State University. [Updated by Parrent, Daniel J.; Funck, James W.; Reeb, James; Brackley, Allen M]. [81905]
223. Wolter, B. H. K.; Fonda, R. W. 2002. Gradient analysis of vegetation on the north wall of the Columbia River Gorge, Washington. Northwest Science. 76(1): 61-76. [65993]
224. Yerkes, Vern P. 1960. Occurrence of shrubs and herbaceous vegetation after clear cutting old-growth Douglas-fir. Res. Pap. PNW-34. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 12 p. [8937]
225. Zasada, John C.; Strong, Terry F. 2008. Acer L.: maple. In: Bonner, Franklin T.; Karrfalt, Robert P., eds. Woody plant seed manual. Agric. Handbook No. 727. Washington, DC: U.S. Department of Agriculture, Forest Service: 204-216. [79013]
226. Zasada, John C.; Tappeiner, John C., II; Max, Timothy A. 1990. Viability of bigleaf maple seeds after storage. Western Journal of Applied Forestry. 5(2): 52-55. [10940]