Index of Species Information
SPECIES: Vaccinium vitis-idaea
|
|
 |
| Lingonberry. Image by Mary Ellen (Mel) Harte, Bugwood.org. |
Introductory
SPECIES: Vaccinium vitis-idaea
AUTHORSHIP AND CITATION:
Tirmenstein, D. 1991. Vaccinium vitis-idaea. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station,
Fire Sciences Laboratory (Producer). Available:
https://www.fs.usda.gov/database/feis/plants/shrub/vacvit/all.html [].
Revisions:
17 September 2013: The Fire Case Study of Zasada and others' [123,124,125]
study was converted to a Research Project Summary. Images were added
on 30 August 2018.
ABBREVIATION:
VACVIT
SYNONYMS:
NO-ENTRY
NRCS PLANT CODE:
VAVI
VAVIM
COMMON NAMES:
lingonberry
lowbush cranberry
northern mountain cranberry
mountain cranberry
TAXONOMY:
The scientific name of lingonberry is Vaccinium vitis-idaea Linnaeus
(Ericaceae) [6,106,107]. Northern mountain cranberry (Vaccinium
vitis-idaea subsp. minus (Lodd.) Hulten) is the only recognized subspecies
occurring in North America [54].
In some areas, lingonberry hybridizes with dwarf bilberry (V.
myrtillus) [1]. A naturally occurring hybrid (V. x intermedium Ruthe.)
has been identified [87].
LIFE FORM:
Shrub
FEDERAL LEGAL STATUS:
No special status
OTHER STATUS:
NO-ENTRY
DISTRIBUTION AND OCCURRENCE
SPECIES: Vaccinium vitis-idaea
GENERAL DISTRIBUTION:
Lingonberry is a circumpolar, circumboreal species that occurs
throughout parts of North America, Eurasia, and Japan [101,106]. The
New World subspecies (ssp. minus) extends from northwestern Greenland
across the Canadian Arctic southward to New England [114]. It grows
westward to the Great Lakes and British Columbia and reaches islands in
the Bering Sea [42,114]. In North America, lingonberry is
restricted to areas north of the glacial boundary [106]. The subspecies
vitis-idaea occurs throughout northern Europe from Scandinavia to
northern Italy and the Caucasus, across northern Siberia and Japan
southward into northern China and Korea [42].
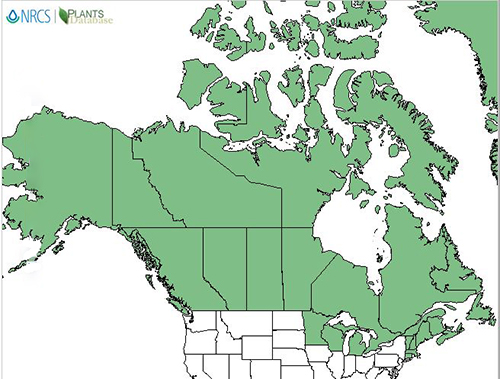 |
| Distribution of lingonberry in North America. Map courtesy of USDA, NRCS. 2018. The PLANTS Database.
National Plant Data Team, Greensboro, NC. [2018, August 30] [103]. |
ECOSYSTEMS:
FRES10 White - red - jack pine
FRES11 Spruce - fir
FRES19 Aspen - birch
FRES23 Fir - spruce
FRES26 Lodgepole pine
FRES44 Alpine
STATES:
AK CT ME MA MN NH VT WI AB BC
LB MB NB NF NT NS ON PE PQ SK
YT
BLM PHYSIOGRAPHIC REGIONS:
None
KUCHLER PLANT ASSOCIATIONS:
K015 Western spruce - fir forest
K093 Great Lakes spruce - fir forest
K094 Conifer bog
K095 Great Lakes pine forest
K096 Northeastern spruce - fir forest
K106 Northern hardwoods
K107 Northern hardwoods - fir forest
K108 Northern hardwoods - spruce forest
SAF COVER TYPES:
1 Jack pine
5 Balsam fir
12 Black spruce
13 Black spruce - tamarack
16 Aspen
17 Pin cherry
18 Paper birch
38 Tamarack
107 White spruce
201 White spruce
202 White spruce - paper birch
204 Black spruce
218 Lodgepole pine
251 White spruce - aspen
253 Black spruce - white spruce
254 Black spruce - paper birch
SRM (RANGELAND) COVER TYPES:
None
HABITAT TYPES AND PLANT COMMUNITIES:
Lingonberry grows as an understory dominant or codominant in a
variety of forest communities including many dominated by jack pine
(Pinus banksiana) and lodgepole pine (P. contorta). It also occurs as a
dominant or indicator in dwarf shrub and shrub tundra communities.
Common codominants include dwarf birch (Betula nana), alpine bearberry
(Arctostaphylos alpina), Labrador tea (Ledum spp.), feather moss
(Pleurozium spp.), willow (Salix spp.), sedges (Carex spp.), lichen
(Cladina spp.), and crowberry (Empetrum nigrum).
Lingonberry is listed as a dominant or indicator in the following
plant association, ecosystem association, habitat type, and community
type classifications:
Forest community types of west-central Alberta in relation to selected
environmental factors [17]
Field guide to forest ecosystems of west-central Alberta [18]
Vegetation types in northwestern Alaska and comparisons with
communities in other arctic regions [39]
Plant associates: Cloudberry (Rubus chamaemorus), Canada beadruby
(Maianthemum canadense), prickly rose (Rosa acicularis), paper birch (B.
papyrifera), sedge, mountain-laurel (Kalmia angustifolia), bearberry
(Arctostaphylos uva-ursi), crowberry, twinflower (Linnaea borealis),
willow, bog blueberry (Vaccinium uliginosum), fireweed (Epilobium
angustifolium), bluejoint reedgrass (Calamagrostis canadensis), bog
Labrador tea, and feather moss commonly occur with lingonberry in
white and black spruce and jack pine communities [7,22,26,38,48,120].
Willows, bog Labrador tea, prickly rose, crowberry, bog blueberry,
sedges, cottongrass (Eriophorum vaginatum), and cloudberry are common
associates in treeless sphagnum bogs, cottongrass muskeg, and dwarf
shrub marsh communities [84,111,114,120].
MANAGEMENT CONSIDERATIONS
SPECIES: Vaccinium vitis-idaea
IMPORTANCE TO LIVESTOCK AND WILDLIFE:
Browse: Lingonberry browse is readily eaten by barren-ground
caribou, black bear, moose, arctic hare, and snowshoe hare [38,42]. In
parts of Alaska, it is an important if not key moose browse [3].
Utilization by moose is typically heaviest when available browse is
limited and when light snow accumulations allow the animals to reach the
plants easily [93]. On the Kenai Peninsula, it may comprise up to 25
percent of winter moose diets. Moose may dig through 20 inches (50 cm)
of snow to feed on the foliage, but if winter snow depths are excessive,
the animals rarely expend the energy necessary to reach the plants [77].
Generally, moose eat only trace amounts of lingonberry during the
summer [10,77].
In some parts of Canada, lingonberry browse is a primary food of
barren-ground caribou [73]. The evergreen leaves are an important item
in the winter diet [38]. In the Mackenzie District of northwestern
Saskatchewan, leaves of lingonberry and bog blueberry (V.
uliginosum) accounted for 21.5 percent of the barren-ground caribou
winter diet but only 3.8 percent of the summer diet [38]. However, in
some areas, caribou continue to feed heavily on lingonberry
browse throughout the summer [111].
In Newfoundland, snowshoe hares often consume large amounts of shoots
during the winter [38]. Where snow depths prohibit winter use, hares
may feed on leaves made available by melting snows. Seasonal percent
composition of leaves of lingonberry and bog blueberry in the
diet of snowshoe hares in Newfoundland was as follows [118]:
winter April May summer fall
0.3 17.4 9.3 6.6 10.9
Lingonberry browse is of little value to domestic livestock but
provides some winter browse for reindeer [23,73]. It is not eaten by
domestic sheep if more preferred forage is available [88].
Fruit: Berries of lingonberry are an important food source for
many species of birds and mammals. Many wildlife species feed on fruit
left on the ground from the previous year [38,55]. Berries are an
important spruce grouse food during spring, summer, and fall. Berries
persisting from the previous year are eaten from late spring through
early August. In interior Alaska, percent volume use of mountain
cranberry by spruce grouse was 37.6 in July and August, 40.1 in
September, and 17.3 in September [29].
In many areas, berries are an essential food source for birds migrating
northward in the spring [38,55]. The common raven, ring-necked
pheasant, rock ptarmigan, sea gulls, geese, grouse, partridges, and many
species of songbirds, such as the scarlet tanager, eastern bluebird, and
thrushes, readily consume lingonberry fruit [38,42,88]. Fruit of
Vacciniums are readily eaten by the northern mockingbird, rufous-sided
towhee, gray catbird, American robin, brown thrasher, ruffed grouse,
spruce grouse, whimbrel, herring gull, and Canada goose [72,105,106].
The red-backed vole eats large quantities of lingonberry fruit
during the fall. Berries are a primary winter food source as well; the
rodents burrow under snow to reach the persistent fruit [117]. The red
fox also consumes large amounts of fruit during late fall [38].
Lingonberry fruit is an important black bear food in many areas
but is of particular importance in Alaska [40]. Berries remain on the
plant over winter, and black bears begin feeding on berries during the
early spring as soon as the snow has melted [38,40]. Fruit again
assumes importance in black bear diets during the fall [40]. Many other
mammals, including the polar bear, eastern chipmunk, and white-footed
mouse, also feed on the fruit of lingonberry [38,55]. Fruits of
many Vacciniums are readily eaten by species such as the red squirrel,
gray fox, skunks, and chipmunks [72,106].
PALATABILITY:
Lingonberry browse is at least seasonally palatable to many
species of mammals including the barren-ground caribou, snowshoe and
arctic hares, and moose. Berries are readily eaten by a variety of
birds and mammals. Palatability of the fruit increases over winter
[99].
NUTRITIONAL VALUE:
Browse: Nutrient content of browse varies according to factors such as
soils, phenological development, and proximity to smelters [42,45,95].
Calcium, manganese, aluminum, silver, lead, and boron tend to accumulate
in plant tissue even at low soil levels [42]. Food value peaks in
summer [38]. In winter, acid-detergent, fiber, and lignin content
increase but levels of magnesium, zinc, manganese, calcium, potassium,
sodium, copper, and iron decline. Protein content remains relatively
constant throughout the year at 5 to 6 percent [77]. Energy content has
been estimated at 509 kcal/100 g [73]. Nutritional value of browse from
the Kenai Peninsula of Alaska was documented as follows [77]:
August February
Protein (%) 5.7 5.4
Ca (ppm) 4920.0 26.7
Mg (ppm) 1328.0 4.6
K (ppm) 438.3 29.8
Na (ppm) 55.0 22.8
Cu (ppm) 5.8 0.2
Fe (ppm) 51.3 3.2
Mg (ppm) 17.6 1.9
Zn (ppm) 8.3 0.3
Fruit: Berries are high in tannins and anthocyanins. The caloric
content is moderate [38].
COVER VALUE:
NO-ENTRY
VALUE FOR REHABILITATION OF DISTURBED SITES:
Potential rehabilitation value of lingonberry has not been well
documented. Plants are able to survive on extremely harsh sites, and
some rehabilitation potential is possible. On the Arctic Coastal Plain,
sprouts have been observed on and under debris left from oil exploration
activities [28].
Lingonberry can be readily propagated from seed and stem or
rhizome cuttings [32,42]. Meristem propagation techniques have also
been described [101]. Stem cuttings root easily if planted in the
spring or early fall but exhibit slow rhizome development and poor
subsequent vegetative spread [32]. Clumps of wild lingonberry
can be divided and transplanted onto disturbed sites [42]. Survival of
these transplants is variable, ranging from 30 to 90 percent [32].
Propagation techniques have been examined in detail [25,32,42,61,63].
OTHER USES AND VALUES:
Lingonberry fruit can be eaten raw or cooked to make a tart sauce
[6,99]. Berries are used to make preserves, jam, jelly, candy, syrup,
pickles, juice beverages, and wine [42,47]. Fruit can be added to rose
hips to make a tasty jelly [38], or added to various ice cream products
[42]. In some areas, berry-picking is an important recreational
activity [59]. Fruit is widely processed and marketed in Japan and
Europe [42] and is harvested commercially in parts of Alaska,
Scandinavia, Russia, and Canada [42,43]. Considerable amounts of
fruit are imported into the United States annually [11]. Much of this
imported fruit is consumed by peoples of Scandinavian descent who use
the so-called "Swedish lingenberry" in traditional dishes [6]. Mountain
cranberry has the potential for more extensive commercial development
[15,37,74]. Some native stands could be managed with a minimum of
cultivation, as are those of low sweet blueberry [74]. The
feasibility of expanded commercial operations is currently being tested
in parts of North America [42].
Many Native Americans and indigenous peoples of Eurasia used the leaves
and fruit of lingonberry as food or medicine [57,106].
Preparations made from the leaves were used to treat bladder problems,
gout, and rheumatism [90]. Medicinal fruit jellies were used to treat
sore throats and colds [106]. The Slave, Athabaska, Cree, and Inuit
people ate the fruit fresh and preserved them for winter use [38,106].
Berries were often boiled and mixed with oil to facilitate storage for
long periods [106].
Arbutin, which is obtained from the leaves and stems, is used by the
pharmaceutical industry in preparations used to treat intestinal
disorders. Lingonberry forms a dense, attractive mat and has
been planted as an ornamental ground cover [24]. It was first
cultivated in 1789 [42]. Lingonberry has shown promise for use
in developing hardy fruit-producing cultivars [64].
OTHER MANAGEMENT CONSIDERATIONS:
Fruit production: Fruit production in lingonberry varies widely
according to geographic location, site factors such as shade and soil,
annual weather conditions, and the genetic make-up of the individual
clone [62,63,71,78,81]. Poor fruit production may be due to a lack of
pollinators, cold damp weather during flowering, late spring frosts, or
hail [42,43,57]. Plants growing in the shade rarely produce fruit or
flowers, but plants growing in full sun commonly bear an abundance of
fruit [62]. Some geographic variation in this pattern has been noted.
On dry sunny sites in Alberta, flower bud production may be greatest in
partial shade of aspen (Populus tremuloides) [38]. In the cool, rainy
climates of the Maritime Provinces, flower bud production is typically
best on exposed sites [38]. Kuchko [57] reported poor yields beneath
forest canopy, although yields were often good in adjacent gaps created
by timber harvest.
Fruit yields are generally greater on peat than on mineral soil [63].
Under experimental conditions, plants produced 82 kg/100 m sq on peat
but produced only 14 kg/100 m sq on mineral soil [63]. Temperatures of
30 degrees F (-1.5 degrees C) can kill 50 percent of all flowers, and
exposure to 26 degrees F (-3.5 degrees C) can destroy 50 percent of the
buds and unripe fruit [62]. In harsh arctic environments, only plants
in protected areas, such as on south-facing rock crevices, flower [42].
Maximum yields in cultivated stands may reach 9,140 pounds per acre
(8,150 kg/ha) [38]. Elsewhere, yields may range from 19.5 pounds per
acre (17.4 kg/ha) [51] in Swedish peatlands to 560 pounds per acre (500
kg/ha) in some Finnish forests [38]. Yields are generally highest where
lingonberry cover is greatest and competitors are few [71].
Details on fruit yields are available [32,57,81].
Cultivation: Lingonberry generally responds more favorably to
fertilizer and irrigation than do other members of the genus [56].
However, the application of fertilizer does not always increase fruit
yields. Comparatively little fertilizer is required for good growth and
development [42]; if too much is added, vegetative growth may be
promoted at the expense of fruit production [101]. Where weeds are a
problem, fertilizer may increase competitors at the expense of mountain
cranberry [62]. Mulches such as milled peat can increase fruit
production in some instances [42]. The effects of mulch, fertilizers,
and irrigation have been examined in detail [32,42,46,53,62,63].
Fruit yields may be increased by various means. Herbicides have been
used to reduce weeds in commercially managed fields of mountain
cranberry [37,63]. Honeybees can be used to supplement native bee
populations when pollinator availability is low [74]. Fruit is
generally harvested by hand [42]. Small comb-sieves or rakes are
commonly used [38,101].
Chemical response: Lingonberry is susceptible to herbicides such
as 2,4-D and 2,4,5-T [38]. These herbicides cause browning of stems and
leaves and at high concentrations can kill the plants [38]. The effect
of herbicides has been documented [8,38].
Damage/disease: Plants can be killed by exposure to cold temperatures
in the absence of a protective snow cover [83]. Unacclimated plants can
be killed by exposure to temperatures of 28 degrees F (-2.5 degrees C)
or below;, acclimated plants can survive exposure to temperatures as low
as 8 degrees F (-22 degrees C) [42]. Lingonberry is susceptible
to several diseases and insect infestations [38,42].
Environmental considerations: Lingonberry growing near smelters
can accumulate high concentrations of heavy metals [95]. Plants growing
near a zinc smelter in Poland exhibited reduced leaf size and other
types of damage [20]. Lingonberry can also accumulate a wide
range of radionuclides such as radium-226, lead-210, and uranium [97].
Tests indicate that summer oil spills are more damaging to mountain
cranberry than those that occur in February [38]. Predisturbance cover
of 48 percent was reduced to 0 by a summer crude oil spill. A
low-intensity winter spill reduced cover to 12 percent while a
high-intensity oil winter spill reduced cover to 6 percent [38].
Recovery of lingonberry can occur 10 to 15 years after an oil
spill [38].
Timber harvest: After some types of logging treatments in a mature
white spruce (Picea glauca) forest in Alaska, cover and frequency of
lingonberry increased fairly rapidly [26].
Biomass: Lingonberry biomass is strongly correlated with canopy
cover [44,77]. Maximum dry matter accumulation occurs in full sunlight
[43]. Holloway [42] observed that 80 percent of the total biomass of
mature plants was underground. Biomass has been examined in detail
[38,42,94].
BOTANICAL AND ECOLOGICAL CHARACTERISTICS
SPECIES: Vaccinium vitis-idaea
GENERAL BOTANICAL CHARACTERISTICS:
Lingonberry is a low, creeping, evergreen subshrub that commonly
reaches 2 to 6 inches (5-15 cm) in height [4,90,114]. It typically
grows in dense rhizomatous colonies and frequently forms mats [106].
Stems are slender and trailing [38,114]; stem
morphology has been
examined in detail [76]. The root system is variable [6]. Plants have
a network of fine, shallow, fibrous roots, and may possess a taproot
[32,38]. The dichotomously branched rhizomes possess numerous hairlike
roots [98]. Maximum rooting depths of 2 to 11 inches (5-28 cm) have
been reported [42,100].
The thick, simple, leaves are obovate, oblong, or elliptic [38,90]. The
green leaves turn purplish in fall [38].
Flowers occur on terminal racemes singly or in groups of up to 15 [90].
Floral morphology has been examined in detail [79]. Fruit is a bright
to dark red, globular berry approximately 0.2 to 0.4 inch (6-10 mm) in
diameter [4,55,106,114]. The four-celled berries are acidic to sour or
bitter [6,90,106]. Yellow, short-beaked seeds average 0.04 inch (1 mm)
in length [42,106].
RAUNKIAER LIFE FORM:
Chamaephyte
Geophyte
REGENERATION PROCESSES:
Lingonberry reproduces through seed and by vegetative means [38].
Seed: In many areas, seedlings first bear fruit at 3 or 4 years of age
[32,63]. However, British studies suggest that few flowers are produced
until plants reach 5 to 10 years of age [88]. Flowers are pollinated by
bumblebees and bee flies (syrphid flies) [38,42]. Plants may be self-
or cross-pollinated, but fruit set is much greater after
cross-pollination [6]. Berries are often produced in abundance. In
parts of North America, berries average 3 to 15 seeds per berry [43].
Seeds are dispersed by birds and mammals [38].
Germination: Seed can germinate on bare ground, but only if conditions
are favorable [38]. Fresh seed generally exhibits best germination
[37,38]. Germination declined from an average of 76.5 percent for seed
extracted from fresh fruit and then planted immediately to less than 10
percent for seed stored 12 to 16 months before planting [38]. In
laboratory tests, good germination was reported after stratification at
32 to 41 degrees F (0-5 degrees C) for up to 5 months [42,61]. Seeds
typically germinate within 3 weeks after exposure to temperatures of -7
to -4 degrees F (20-25 degrees C) in light or dark [32,42]. Germination
characteristics of lingonberry have been examined in detail
[19,62,63].
Seedling establishment: Seedlings are rarely observed in the field
[42,62,75,102]. In Estonia, seedlings are generally observed only in
protected areas such as near tree stumps, fallen logs, or stones [71].
Some seedlings do develop in favorable years in parts of Nova Scotia and
Newfoundland [38].
Seed banking: Seed of lingonberry has been detected in soil
samples [75,106], but seed banking potentials for this species are
unknown. In black spruce (Picea mariana)-jack pine forests of the
Northwest Territories, seeds of lingonberry and bog blueberry
were found in 71 percent of the soil samples tested [50]. Studies near
the Great Slave Lake revealed lingonberry and bog blueberry seed
in 65 percent of the cores sampled. Seed densities averaged 3.3 per
1,000 cc, but only 21 percent were viable. A second study revealed 4.8
seeds per 1,000 cc, but none of the seeds were viable [38].
Vegetative regeneration: Vegetative regeneration is of primary
importance in the lingonberry [88]. Plants commonly expand
through horizontal rhizomes [88]. Rhizomes may sprout singly or in
groups of 1 or 2 per square meter [42]. Large, older clones may be
separated into numerous daughter clones by disturbances such as frost,
fire, or burrowing mammals [38,88]. Rhizome length, depth, and the
location of shoots on the rhizomes are greatly influenced by soil and
other site characteristics [98]. Rhizome depth is inversely related to
the thickness of soil organic layers [98]. Rhizomes grow well in peat
but can also penetrate to mineral soil. In Britain, rhizomes are
generally confined to the humus layer [38] and are estimated to average
4 to 8 inches (10-20 cm) deep [88]. Smith [98] reported that rhizome
depth in Alberta varied from 8 to 11 inches (19-28 cm). He found that
22.1 percent of the shoots were located terminally and 77.9 percent
arose at midrhizome locations. Rhizome characteristics as related to
various site characteristics have been examined in detail.
The trailing or creeping stems of lingonberry also root at the
nodes [38,114]. This mode of regeneration may be important on some
harsh, subarctic sites [38].
SITE CHARACTERISTICS:
Lingonberry is widely distributed in northern temperate forests
and in many arctic and alpine communities [38,60,114]. It commonly
grows on exposed sites, such as windswept crags, bare headlands, rocky
ledges, scree, sea cliffs, hilly rocky barrens, and mountain summits
[21,38,81,88,91]. At the southern edge of its range, lingonberry
occurs primarily in bogs, but in the north it grows on both wet and dry
sites [38]. Lingonberry occurs on high moors, heath barrens,
sand dunes, and in peatlands, forest swamps, and bogs [38,41,114]. In
mature forests, plants often grow on top of decaying tree stumps [42].
Climate: Lingonberry grows under a variety of climatic regimes.
In much of Canada, it occurs in areas characterized by short cool
summers and long cold winters [38]. In black spruce-white spruce-jack
pine forests of northern Canada, its distribution may be correlated with
arctic air masses. However, in harsh rockfield and tussock communities
of the far North, it may be related to the influence of moist Pacific
air masses [38,60]. In taiga communities of Alaska, winters are long
and cold, but summers are short and hot [110]. Mean annual
precipitation is 8 inches (21 cm), and average annual temperature is 20
degrees F (-6.7 degrees C) [84]. In parts of the Northwest Territories,
annual precipitation averages 12 inches (30.4 cm) [68].
Soils: Lingonberry grows on shallow, poorly developed mineral
soil as well as on drained peat [51,88]. Soils are often of low
fertility and have little calcium but may be high in decaying organics
[42,101]. Lingonberry commonly grows on acidic sandy loams or
loamy clays [42,57]. Holloway and others [45] reported poorest
vegetative growth on sandy soils. Soil pH ranges from 2.7 to 8.2, but
best growth has been reported at 4.0 to 4.9 [38,42,49]. Soils are often
characterized by low base saturation and low lime content [45]. Soils
may be derived from a variety of parent materials, including sandstone,
gneiss, granite, and glacial outwash sands and gravel [38].
Elevation: In New England, lingonberry is generally restricted
to higher mountains [55]. In the Northwest Territories, plants often
occur at lower elevations (to 4,950 feet [1,500 m]) [38]. Generalized
elevational ranges by geographic location are as follows:
Location Elevation Authority
Adirondacks up to 5,300 ft (1,615 m) Keeler 1969
e Canada sea level to 4,250 ft (0-1,290 m) Hall and Shay 1981
AB to 7,400 ft (2,250 m) Hall and Shay 1981
Yukon 6,900 to 7,900 ft (2,100-2,400 m) Hall and Shay 1981
SUCCESSIONAL STATUS:
Lingonberry is noted for its wide ecological amplitude [69]. It
is not generally considered a pioneer species but does occur in early
seral stages in some communities [38,65]. Lingonberry persists
indefinitely, unless shaded out by conifers, and assumes a climax role
in various rockfield communities of the far North [38].
Lingonberry commonly invades tundra bog communities dominated by
species such as alpine sweetgrass (Hierochloe alpina), lichens
(Alectoria ochroleuca, A. nitidula), and woodrush (Luzula confusa) from
adjacent summit rockfields. Lingonberry also invades senescent
cottongrass tussock communities and areas of frost activity after the
establishment of initial pioneers. However, on some sites, seral mat
communities made up of lingonberry, crowberry, and lichens
eventually give rise to white spruce stands. In barrens of
Newfoundland, lingonberry grows as a seral species which is
displaced by black spruce and balsam fir [38]. It also occurs in some
early seral communities dominated by paper birch [65].
Black spruce: Lingonberry is important in stable climax black
spruce communities but also dominates many seral stages [34,65,80].
Stands are initially colonized by bryophytes and herbaceous species such
as fireweed and willow [7,27]. Lingonberry generally reaches
stable levels within 25 years after fire or other disturbances [38].
However, maximum cover and frequency were attained at 144 years in
certain black spruce/lingonberry communities [38]. In black
spruce stands in interior Alaska, lingonberry is present within 5
to 30 years after disturbance and persists for many years. It is common
in stands 200 years old or older and represents the most abundant low
shrub in tree-dominated stages [34]. In black spruce stands of the
Northwest Territories, lingonberry remains abundant in 200- to
300-year-old stands despite the decline of most vascular plants [7].
Chapin and others [14] reported that lingonberry becomes more
prominent as succession progresses from immature black spruce to muskeg.
White spruce: Lingonberry occurs in many climax white spruce
forests on uplands of interior Alaska [65]. It is present during the
moss-herb stage which occurs 1 to 5 years after fire [27,34]. Mountain
cranberry peaks and declines after the dense tree stage, which occurs
from 15 to 40-46 years or longer after fire, but remains present in
later stages [34]. Dyrness and others [27] reported that in interior
Alaska, lingonberry was common in 150-year-old white spruce
stands.
Jack pine: In jack pine-lichen woodlands of the northern Canada,
lingonberry is an early colonizer on recently burned sites [13].
It persists after "the cessation of major successional changes" at 25 to
45 years [13] and remains common in stands up to 280 years of age [38].
Tundra communities: In sedge-tussock tundra and shrub tundra
communities of Alaska's Seward Peninsula, bryophytes initially
reestablished burned sites. Bryophytes often reach maximum cover within
2 to 4 years after fire, but the recovery of shrubs such as mountain
cranberry is often much slower. In shrub-tundra communities, mountain
cranberry may not recover to prefire levels even by 5 to 6 years after
fire or other disturbance [84,85].
SEASONAL DEVELOPMENT:
In Alaska, vegetative buds began growth during the first week of June
and underwent rapid elongation throughout June. The growth rate of
terminal vegetative buds decreased by July 1. Leaf expansion began
during the last week of May and the first week of June; all leaves had
expanded within 1 month [42]. Karlsson [52] observed that old leaves
became photosynthetically active approximately 2 weeks after bud break.
Near Mt. Washington, New Hampshire, and in parts of Nova Scotia,
vegetative growth began in late June [38,42]. In Britain and perhaps
elsewhere, leaf expansion can begin as early as March, although it
usually occurs from mid-May to mid-June. Shoot growth generally ends in
mid-July. Leaves may persist for up to 3 years. However, some old
leaves may be shed by August of the second year [38]. Plants become
dormant by fall [42].
Flowers develop from buds initiated the previous year [101]. In
interior Alaska, reproductive bud growth begins in mid-May [42]. In
parts of Britain, two periods of flowering (spring and summer) have been
observed at certain low-elevation sites [88]. Flowering may last 9 to
18 days [57] or as long as 19 to 27 days. Fruit ripens approximately 78
to 84 days after full bloom [42].
Phenological development may be related to the timing of snowmelt [38].
In interior Alaska, plants were in maximum full bloom approximately 6
weeks after snowmelt and exhibited first visible signs of growth 2 weeks
after snowmelt. Unusually cool temperatures can delay phenological
development. Roots and rhizomes undergo two periods of active growth
annually in early spring and fall [42].
FIRE ECOLOGY
SPECIES: Vaccinium vitis-idaea
FIRE ECOLOGY OR ADAPTATIONS:
Lingonberry occurs in a variety of communities across a wide
climatic range. It persists under a regime of relatively frequent fires
but also grows in areas that rarely burn. Black spruce communities are
dependent on frequent fires, and most associated species, including
lingonberry, are well adapted to fire [110]. Fires in black
spruce communities of Alaska and northern Canada are commonly lightning
caused and tend to be large [68,110]. Fire frequencies average 80 to
200 years [96,110]. In moister black spruce/lingonberry
communities in eastern Canada, fires may occur at 500-year intervals
[35]. Lingonberry remains important in jack pine stands that
burn at 20- to 40-year intervals and in Swedish pine forests that burn
every 40 years [13,30].
Fire may be an important factor in treeline communities of the North.
In Siberia, past extensive fires may have destroyed forest communities.
Trees may have been unable to reestablish on these harsh sites under the
current climatic regimes. Low-shrub-dominated tundra communities
composed of species such as lingonberry may have eventually
replaced these forest stands [108]. Fire intervals in shrub subzones of
forest-tundra communities have been estimated at 1,460 years [96].
Lingonberry continues to be abundant on these infrequently burned
sites.
In many forest communities, lingonberry requires fire for its
maintenance [30]. Increases in cover and vigor after fire are commonly
observed [9]. Lingonberry generally reestablishes a site through
sprouting from rhizomes and aerial stems. Very limited reestablishment
may occur on exceptional sites in good years by seed transported from
off-site.
FIRE REGIMES:
Find fire regime information for the plant communities in which this
species may occur by entering the species name in the FEIS home page under
"Find Fire Regimes".
POSTFIRE REGENERATION STRATEGY:
Small shrub, adventitious-bud root crown
Rhizomatous shrub, rhizome in soil
Initial-offsite colonizer (off-site, initial community)
FIRE EFFECTS
SPECIES: Vaccinium vitis-idaea
IMMEDIATE FIRE EFFECT ON PLANT:
Underground regenerative structures of lingonberry generally survive
light fires [102,115]. Plants often survive even when aerial portions are
consumed by fire [92]. However, plants may be killed by moderate to heavy,
duff-consuming fires [115]. Survival is related to many factors including
soil moisture levels, season of burn, fire severity and intensity, and rhizome
depth [38].
Rhizomes can sometimes survive soil surface temperatures of 820 degrees
F (438 degrees C) [102]. In arctic tussock communities, plants often
survive severe fires which remove all aboveground material [116]. The
heat-sensitive seeds of lingonberry are usually destroyed by fire
[115].
DISCUSSION AND QUALIFICATION OF FIRE EFFECT:
NO-ENTRY
PLANT RESPONSE TO FIRE:
Lingonberry commonly sprouts from rhizomes or buds located on
surviving portions of aerial stems after fire damages or consumes
aboveground material [115,116]. Sprouting from stumps, or "rootstocks"
has also been reported [13,65]. Reestablishment through seed is
extremely rare [102]. Surviving portions of the aerial stems sprout
within a short time, but rhizome sprouting may be delayed until the
following year [115].
The speed of reestablishment varies according to the season of burn,
site characteristics, and fire intensity and severity. Reestablishment
is generally rapid after light fires [27]; plants are often common on
lightly burned sites [102]. Regeneration may be slow after hot fires
that damage or destroy underground regenerative structures [27,102,111].
In northern spruce communities, intense, stand-destroying, late summer
fires which consume the organic layer [110] can be particularly damaging
to lingonberry [65].
On some sites, plants may sprout within months after a light burn and
regain prefire cover within a few years [102]. Lingonberry
generally appears within the first 6 years after fire in black
spruce-lichen, jack pine-lichen, and white spruce-birch communities
[38]. Residual survivors were observed at the end of the fifth growing
season in black spruce communities of southeastern Manitoba [16]. In a
severely burned black spruce community of interior Alaska, it became
abundant within 5 years after fire but set little fruit [117]. Viereck
[109] observed slow recovery after fire in a black spruce/feather moss-
lichen community of interior Alaska. Recovery was as follows:
percent cover (1971 fire)
unburned 1972 1975 1980
18 1 1 7
Lingonberry was present within 1 to 5 years after fire in white
spruce communities of Alaska [34]. Lingonberry is a common early
colonizer in jack pine communities, although reestablishment generally
takes at least several years [13]. In North America, postfire recovery
may be more rapid in moister, eastern boreal forests [68].
Reestablishment of lingonberry is often slow in tundra
communities [85]. Plants attained prefire coverage by the end of two
full growing seasons in arctic tussock communities [116]. In
northwestern Alaska, production was still significantly lower on sites
burned 13 years earlier than on unburned sites [33].
DISCUSSION AND QUALIFICATION OF PLANT RESPONSE:
Fire severity: Recovery is typically much more rapid after light fires.
Postfire recovery in Alaska has been documented as follows [111]:
percent cover
1971 1972 1973 1974 control
heavy burn .05 .50 .30 .90 ----
light burn 3.45 3.55 1.65 6.10 20.35
heavy burn 0 .15 .20 .35 6.90
In sedge-shrub tundra on the Seward Peninsula of Alaska, little or no
sprouting was observed within 2 years on severely burned sites where
lingonberry was a prefire dominant. After light to moderate
severity burns in sedge-tussock communities, lingonberry
sprouted and recovered relatively quickly. Shoot densities increased
significantly on two of the four sites, from 43 shoots per meter square
to 126 per meter square and from 25 per meter square to 43 per meter
square. However, sprouts were generally located on the surface or sides
of tussocks, suggesting that they escaped burning within the tussock
mass [84].
Recovery of lingonberry may be relatively slow in many types of
tundra shrub communities. Recovery of lingonberry by tundra
community was as follows [84]:
sedge tussock-shrub tundra (burned 1977)
Frequency (no. of plots)* Cover (%)
1973 1978 1979 1973 1978 1979
site 2 10 10 10 6.9 0.8 1.3
site 3 -- 10 10 --- 0.5 0.5
site 4 -- 10 10 --- 0.5 0.5
site 5 -- 9 9 --- 0.5 0.5
*Number of 1-m2 plots in which species occurs (ten plots sampled)
birch and ericaceous shrub tundra (burned 1977)
1973 1978 1979
Cover Cover Cover
_ _ _
Freq.* (x %) Freq.* (x %) Freq.* (x %)
nonfrost boils 10 7 0 0 0 0
frost boils 10 4 0 0 0 0
*Number of 1-m2 quadrats in which species occurs/no. of quadrats on that
site X 10
sedge-shrub tundra (burned 1977)
Prefire (1973) 1 yr after (1978) 2 yrs after (1979)
Freq.* Cover** Freq.* Cover** Dens.*** Freq.* Cover** Dens.***
site 8 10 2.8 3 0.2 2 4 0.2 3
site 9 10 15.5 6 0.3 2 7 0.3 5
* Number of 1-m2 plots in which the species occurs (ten plots sampled)
** Mean percent cover averaged over 10 plots
*** shoots/m2
Postfire frequencies of lingonberry 1 year after a summer fire in
sedge tussock-shrub communities of the Seward Peninsula of Alaska were
greatly reduced [119]:
sampling date late May 1978 mid-June 1978
freq. % freq. %
burned 0.23 0.05
unburned 1.00 1.00
See the Research Project Summary of Wright's [119] study for further information.
For information on prescribed fire and postfire responses of many plant
species, including lingonberry, see these Research Project Summaries:
FIRE MANAGEMENT CONSIDERATIONS:
Postfire biomass: Postfire reduction in lingonberry production
was as follows after a fire in an arctic tussock community [116]:
mean annual production (g/m sq)
burned unburned
site 1 0.1 5.8
site 2 0.6 7.5
site 3 0.6 1.8
site 4 4.0 9.5
Biomass: Biomass following a late June wildfire in interior Alaska was
measured at 0.04 grams per square meter during postfire year 1, 0.08
grams per square meter during postfire year 2, and 1.4 grams per square
meter during postfire year, compared to a control measurement of 5.1
grams per square meter [108].
Fuels and flammability: Engelmark [30] reported that Vacciniums are
highly flammable due to specific chemical properties. In northern
Sweden, species such as lingonberry can serve as ignition points
and as a continuous fuel mat for surface fires. In many black spruce
stands of Alaska and northern Canada, an open, highly flammable,
ericaceous shrub layer can carry a fire [110]. However, Quintilio and
others [82] observed that an extensive mat of lingonberry and
alpine bearberry served as an effective fire barrier in a jack pine
stand near Darwin Lake, Alberta. Fire seldom penetrated more than a few
centimeters into the vegetative mat. The extensive ground mat
noticeably reduced the fire spread rate and coverage [82].
REFERENCES
SPECIES: Vaccinium vitis-idaea
REFERENCES:
1. Ahokas, Hannu. 1971. Notes on polyploidy and hybridity in Vaccinium
species. Annales Botanici Fennici. 8: 254-256. [9699]
2. Ahokas, Hannu. 1979. Artificial, reciprocal hybrids between Vaccinium
microcarpum and V. vitis-idaea. Annales Botanici Fennici. 16: 3-6.
[9695]
3. Allen, Arthur W.; Jordan, Peter A.; Terrell, James W. 1987. Habitat
suitability index models: moose, Lake Superior region. Biol. Rep. 82
(10.155). Washington, DC: U.S. Department of the Interior, Fish and
Wildlife Service. 47 p. [11710]
4. Anderson, J. P. 1959. Flora of Alaska and adjacent parts of Canada.
Ames, IA: Iowa State University Press. 543 p. [9928]
5. Bernard, Stephen R.; Brown, Kenneth F. 1977. Distribution of mammals,
reptiles, and amphibians by BLM physiographic regions and A.W. Kuchler's
associations for the eleven western states. Tech. Note 301. Denver, CO:
U.S. Department of the Interior, Bureau of Land Management. 169 p.
[434]
6. Biermann, John E. 1975. A description of Vaccinium vitis-idaea. Fruit
Varieties Journal. 29(1): 5-7. [1901]
7. Black, R. A.; Bliss, L. C. 1978. Recovery sequence of Picea mariana -
Vaccinium uliginosum forests after burning near Inuvik, Northwest
Territories, Canada. Canadian Journal of Botany. 56: 2020-2030. [7448]
8. Bovey, Rodney W. 1977. Response of selected woody plants in the United
States to herbicides. Agric. Handb. 493. Washington, DC: U.S. Department
of Agriculture, Agricultural Research Service. 101 p. [8899]
9. Bradshaw, Richard H. W.: Zackrisson, Olle. 1990. A two thousand year
history of a northern Swedish boreal forest stand. Journal of Vegetation
Science. 1(4): 519-528. [12762]
10. Butler, C. E. 1986. Summer food utilization and observations of a tame
moose Alces alces. Canadian Field-Naturalist. 100: 85-88. [8871]
11. Camp, W. H. 1942. On the structure of populations in the genus
Vaccinium. Brittonia. 4(2): 189-204. [9512]
12. Camp, W. H. 1945. The North American blueberries with notes on other
groups of Vacciniaceae. Brittonia. 5(3): 203-275. [9515]
13. Carroll, S. B.; Bliss, L. C. 1982. Jack pine - lichen woodland on sandy
soils in northern Saskatchewan and northeastern Alberta. Canadian
Journal of Botany. 60: 2270-2282. [7283]
14. Chapin, F. Stuart, III; Van Cleve, Keith. 1981. Plant nutrient
absorption and retention under differing fire regimes. In: Mooney, H.
A.; Bonnicksen, T. M.; Christensen, N. L.; [and others], technical
coordinators. Fire regimes and ecosystem properties: Proceedings of the
conference; 1978 December 11-15; Honolulu, HI. Gen. Tech. Rep. WO-26.
Washington, DC: U.S. Department of Agriculture, Forest Service: 301-321.
[4397]
15. Christ, E. 1977. Crossbreedings between cranberries (Vaccinium
macrocarpon Ait.) and cowberries (Vaccinium vitis idaea L.). Acta
Horticulturae. 61: 285-294. [9506]
16. Chrosciewicz, Z. 1976. Burning for black spruce regeneration on a
lowland cutover site in southeastern Manitoba. Canadian Journal of
Forest Research. 6(2): 179-186. [7280]
17. Corns, I. G. W. 1983. Forest community types of west-central Alberta in
relation to selected environmental factors. Canadian Journal of Forest
Research. 13: 995-1010. [691]
18. Corns, I. G. W.; Annas, R. M. 1986. Field guide to forest ecosystems of
west-central Alberta. Edmonton, AB: Canadian Forestry Service, Northern
Forestry Centre. 251 p. [8998]
19. Crossley, John A. 1974. Vaccinium L. Blueberry. In: Schopmeyer, C. S.,
ed. Seeds of woody plants in the United States. Agric. Handb. 450.
Washington, DC: U.S. Department of Agriculture, Forest Service: 840-843.
[7774]
20. Czuchajowska, Zuzanna. 1987. Influence of zinc smelter emissions on
leaves of Pinus sylvestris and Vaccinium spp. as revealed by some
morphological & ecophys. indices. Environmental and Experimental
Biology. 27(1): 67-83. [9255]
21. Dansereau, Pierre. 1959. The principal plant associations of the Saint
Lawrence Valley. No. 75. Montreal, Canada: Contrib. Inst. Bot. Univ.
Montreal. 147 p. [8925]
22. Daubenmire, Rexford. 1953. Notes on the vegetation of forested regions
of the far northern Rockies and Alaska. Northwest Science. 27: 125-138.
[10816]
23. Dayton, William A. 1931. Important western browse plants. Misc. Publ.
101. Washington, DC: U.S. Department of Agriculture. 214 p. [768]
24. De Vault, Dorothea. 1977. Four uncommon groundcovers. American Rock
Garden Society Bulletin. 35(1): 36-40. [9508]
25. Doran, William L. 1957. Propagation of woody plants by cuttings.
Experiment Station Bul. No. 491. Amherst, MA: University of
Massachusetts, College of Agriculture. 99 p. [6399]
26. Dyrness, C. T.; Viereck, L. A.; Foote, M. J.; Zasada, J. C. 1988. The
effect on vegetation and soil temperature of logging flood-plain white
spruce. Res. Pap. PNW-RP-392. Portland, OR: U.S. Department of
Agriculture, Forest Service, Pacific Northwest Research Station. 45 p.
[7471]
27. Dyrness, C. T.; Viereck, L. A.; Van Cleve, K. 1986. Fire in taiga
communities of interior Alaska. In: Forest ecosystems in the Alaskan
taiga. New York: Springer-Verlag: 74-86. [3881]
28. Ebersole, James J. 1987. Short-term vegetation recovery at an Alaskan
arctic coastal plain site. Arctic and Alpine Research. 19(4): 442-450.
[9476]
29. Ellison, Laurence. 1966. Seasonal foods and chemical analysis of winter
diet of Alaskan spruce grouse. Journal of Wildlife Management. 30(4):
729-735. [9735]
30. Engelmark, Ola. 1987. Fire history correlations to forest type and
topography in northern Sweden. Annales Botanici Fennici. 24(4): 317-324.
[6688]
31. Eyre, F. H., ed. 1980. Forest cover types of the United States and
Canada. Washington, DC: Society of American Foresters. 148 p. [905]
32. Fernqvist, I. 1977. Results of experiments with cowberries and
blueberries in Sweden. Acta Horticulturae. 61: 295-300. [9609]
33. Fetcher, Ned; Beatty, Thomas F.; Mullinax, Ben; Winkler, Daniel S. 1984.
Changes in arctic tussock tundra thirteen years after fire. Ecology.
65(4): 1332-1333. [7234]
34. Foote, M. Joan. 1983. Classification, description, and dynamics of plant
communities after fire in the taiga of interior Alaska. Res. Pap.
PNW-307. Portland, OR: U.S. Department of Agriculture, Forest Service,
Pacific Northwest Forest and Range Experiment Station. 108 p. [7080]
35. Foster, David R. 1985. Vegetation development following fire in Picea
mariana (black spruce) - Pleurozium forests of south-eastern Labrador,
Canada. Journal of Ecology. 73: 517-534. [7222]
36. Garrison, George A.; Bjugstad, Ardell J.; Duncan, Don A.; [and others].
1977. Vegetation and environmental features of forest and range
ecosystems. Agric. Handb. 475. Washington, DC: U.S. Department of
Agriculture, Forest Service. 68 p. [998]
37. Hall, Ivan V.; Beil, Charles E. 1970. Seed germination, pollination, and
growth of Vaccinium vitis-idaea var. minus Lodd. Canadian Journal of
Plant Science. 50(6): 731-732. [9174]
38. Hall, Ivan V.; Shay, Jennifer, M. 1981. The biological flora of Canada.
3. Vaccinium vitis-idaea L. var. minus Lodd. Supplementary Account.
Canadian Field-Naturalist. 95(4): 434-464. [9125]
39. Hanson, Herbert C. 1953. Vegetation types in northwestern Alaska and
comparisons with communities in other arctic regions. Ecology. 34(1):
111-140. [9781]
40. Hatler, David F. 1972. Food habits of black bears in interior Alaska.
Canadian Field-Naturalist. 86(1): 17-31. [10389]
41. Heinselman, M. L. 1970. Landscape evolution, peatland types and the
environment in the Lake Agassiz Peatlands Natural Area, Minnesota.
Ecological Monographs. 40(2): 235-261. [8378]
42. Holloway, Patricia Sue. 1981. Studies on vegetative and reproductive
growth of lingonberry, Vaccinium vitis-idaea L. Saint Paul, MN:
University of Minnesota. 148 p. Thesis. [9610]
43. Holloway, Patricia S.; Stushnoff, Cecil; Wildung, David K. 1982.
Gibberellic acid-induced fruiting of lingonberries, Vaccinium
vitis-idaea L. ssp. minus (Lodd.) Hult. HortScience. 17(6): 953-954.
[9192]
44. Holloway, Patricia S.; Van Veldhuizen, Robert M.; Stushnoff, Cecil;
Wildung, David K. 1982. Effects of light intensity on vegetative growth
of lingonberries. Canadian Journal of Plant Science. 62(4): 965-968.
[9164]
45. Holloway, Patricia S.; Van Veldhuizen, Robert M.; Stushnoff, Cecil;
Wildung, David K. 1982. Vegetative growth and nutrient levels of
lingonberries grown in four Alaskan substrates. Canadian Journal of
Plant Science. 62(4): 969-977. [9163]
46. Ingestad, Torsten. 1973. Mineral nutrient requirements of Vaccinium
vitis idaea and V. myrtillus. Physiological Plant. 29(2): 239-246.
[9116]
47. Iwagaki, H.; Ishikawa, S.; Tamada, T.; Koike, H. 1977. The present
status of blueberry work and wild Vaccinium species in Japan. Acta
Horticulturae. 61: 331-334. [9701]
48. Jameson, J. S. 1961. Observations on factors influencing jack pine
reproduction in Saskatchewan. Technical Note No. 97. Forest Research
Division, Department of Forestry, Canada. 24 p. [7284]
49. Jeglum, John K. 1971. Plant indicators of pH and water level in
peatlands at Candle Lake, Saskatchewan. Canadian Journal of Botany. 49:
1661-1676. [7450]
50. Johnson, E. A. 1975. Buried seed populations in the subarctic forest
east of Great Slave Lake, Northwest Territories. Canadian Journal of
Botany. 53: 2933-2941. [6466]
51. Kardell, Lars. 1986. Occurrence and berry production of Rubus
chamaemorus L., Vaccinium oxycoccus L. & Vaccinium microcarpum Turcz.
and Vaccinium vitis-idaea on Swedish peatlands. Scandinavian Journal
of Forest Research. 1(1): 125-140. [3711]
52. Karlsson, Staffan P. 1985. Photosynthetic characteristics and leaf
carbon economy of a deciduous and evergreen dwarf shrub: Vaccinium
uliginosum and V. vitis-idaea L. Holarctic Ecology. 8: 9-17. [9158]
53. Karlsson, P. Staffan. 1985. Effects of water and mineral nutrient supply
on a deciduous and an evergreen dwarf shrub: Vaccinium uliginosum L. and
V. vitis-idaea L. Holarctic Ecology. 8: 1-8. [9157]
54. Kartesz, John T. 1994. A synonymized checklist of the vascular flora of
the United States, Canada, and Greenland. Volume II--thesaurus. 2nd ed.
Portland, OR: Timber Press. 816 p. [23878]
55. Keeler, Harriet L. 1969. Vacciniaceae--huckleberry family. In: Our
northern shrubs and how to identify them. New York: Dover Publications,
Inc.: 315-342. [9272]
56. Korcak, Ronald F. 1988. Nutrition of blueberry and other calcifuges.
Horticultural Reviews. 10: 183-227. [9612]
57. Kuchko, A.A. 1988. Bilberry and cowberry yields and the factors
controlling them in the forests of Karelia, U.S.S.R. Acta Bot. Fennica.
136: 23-25. [8903]
58. Kuchler, A. W. 1964. Manual to accompany the map of potential vegetation
of the conterminous United States. Special Publication No. 36. New York:
American Geographical Society. 77 p. [1384]
59. Kuusipalo, Jussi; Niemensivu, Helena; Berg, Mari-Anna; Mikkola, Marja.
1989. A cross-sectional population survey on the consumption pattern of
berries and berry products in Finland. Silva Fennica. 23(1): 59-69.
[8529]
60. Larsen, James A. 1971. Vegetational relationships with air mass
frequencies: boreal forest and tundra. Arctic. 24: 177-194. [8258]
61. Lehmushovi, Aaro. 1975. Methods of propagating the cowberry. Annales
Agriculturae Fenniae. 14(4): 325-333. [9776]
62. Lehmushovi, A. 1977. Trials with the cowberry in Finland. Acta
Horticulturae. 61: 301-308. [9680]
63. Lehmushovi, Aaro; Sako, Jaakko. 1975. Domestication of the cowberry
(Vaccinium vitis-idaea L.) in Finland. Annales Agriculturae Fenniae. 14:
227-230. [9520]
64. Liebster, G. 1977. Experimental and research work on fruit species of
the genus Vaccinium in Germany. Acta Horticulturae. 61: 19-24. [9693]
65. Lutz, H. J. 1953. The effects of forest fires on the vegetation of
interior Alaska. Juneau, AK: U.S. Department of Agriculture, Forest
Service, Pacific Northwest Forest and Range Experiment Station. 36 p.
[7076]
66. Lutz, H. J. 1956. Ecological effects of forest fires in the interior of
Alaska. Tech. Bull. No. 1133. Washington, DC: U.S. Department of
Agriculture, Forest Service. 121 p. [7653]
67. Lyon, L. Jack; Stickney, Peter F. 1976. Early vegetal succession
following large northern Rocky Mountain wildfires. In: Proceedings, Tall
Timbers fire ecology conference and Intermountain Fire Research Council
fire and land management symposium; 1974 October 8-10; Missoula, MT. No.
14. Tallahassee, FL: Tall Timbers Research Station: 355-373. [1496]
68. Maikawa, E.; Kershaw, K. A. 1976. Studies on lichen-dominated systems.
XIX. The postfire recovery sequence of black spruce-lichen woodland in
the Abitau Lake region, N.W.T. Canadian Journal of Botany. 54:
2679-2687. [7225]
69. Maini, J. S. 1966. Pytoecological study of sylvotundra at Small Tree
Lake, N.W.T. Arctic. 19: 220-243. [8259]
70. Mallett, K. I.; Volney, W. J. A. 1990. Relationships among jack pine
budworm damage, selected tree characteristics, and Armillaria root rot
in jack pine. Canadian Journal of Forest Research. 20: 1791-1795.
[12760]
71. Manni, R. 1988. Biology and berry production of the cowberry in Estonian
SSR. Acta Bot. Fennica. 136: 33-36. [8904]
72. Martin, Alexander C.; Zim, Herbert S.; Nelson, Arnold L. 1951. American
wildlife and plants. New York: McGraw-Hill Book Company, Inc. 500 p.
[4021]
73. Miller, Donald R. 1976. Taiga winter range relationships and diet.
Canadian Wildlife Service Rep. Series No. 36. Ottawa, ON: Environment
Canada, Wildlife Service. 42 p. (Biology of the Kaminuriak population of
barren-ground caribou; pt 3). [13007]
74. Mohr, H. A.; Kevan, P. G. 1987. Pollinators and pollination requirements
of lowbush blueberry (Vaccinium angustifolium Ait. and V. myrtilloides
Michx.) and cranberry .... Proceedings of the Entomological Society of
Ontario. 118(0): 149-154. [10806]
75. Morin, Hubert; Payette, Serge. 1988. Buried seed populations in the
montane, subalpine, and alpine belts of Mont Jacques-Cartier, Quebec.
Canadian Journal of Botany. 66: 101-107. [6376]
76. Odell, A. E.; Vander Kloet, S. P.; Newell, R. E. 1989. Stem anatomy of
Vaccinium section Cyanococcus and related taxa. Canadian Journal of
Botany. 67(8): 2328-2334. [8944]
77. Oldemeyer, John L.; Seemel, Robert K. 1976. Occurrence and nutritive
quality of lowbush cranberry on the Kenai Peninsula, Alaska. Canadian
Journal of Botany. 54: 966-970. [9641]
78. Paal, Taimi. 1988. The structure of South Karelian (U.S.S.R.) cowberry
coenopopulations. Acta Botanica Fennica. 136: 27-31. [9606]
79. Palser, Barbara F. 1961. Studies of floral morphology in the Ericales.
V. Organography and vascular anatomy in several United States species of
the Vacciniaceae. Botanical Gazette. 123(2): 79-111. [9032]
80. Parminter, John. 1983. Fire-ecological relationships for the
biogeoclimatic zones and subzones of the Fort Nelson Timber Supply Area:
summary report. In: Northern Fire Ecology Project: Fort Nelson Timber
Supply Area. Victoria, BC: Province of British Columbia, Ministry of
Forests. 53 p. [9203]
81. Penney, B. G.; McRae, K. B.; Hall, I. V.; Morris, R. F.; Hendrickson,
P. A. 1985. Effect of harvest date and location on the yield of
Vaccinium vitis-idaea L. var. minus Lodd in eastern Newfoundland. Crop
Research. 25(1): 21-26. [9603]
82. Quintilio, D.; Fahnestock, G. R.; Dube, D. E. 1977. Fire behavior in
upland jack pine: the Darwin Lake project. Information Report NOR-x-174.
Edmonton, AB: Forestry Service, Fisheries and Environment Canada,
Northern Forest Research Centre. 49 p. [7244]
83. Raatikainen, Mikko; Vanninen, Irene. 1988. The effects of the 1984-1985
cold winter on the bilberry and ligonberry yield in Finland. Acta Bot.
Fennica. 136: 43-47. [8902]
84. Racine, Charles H. 1981. Tundra fire effects on soils and three plant
communities along a hill-slope gradient in the Seward Peninsula, Alaska.
Arctic. 34(1): 71-84. [7233]
85. Racine, Charles H.; Johnson, Lawrence A.; Viereck, Leslie A. 1987.
Patterns of vegetation recovery after tundra fires in northwestern
Alaska, U.S.A. Arctic and Alpine Research. 19(4): 461-469. [6114]
86. Raunkiaer, C. 1934. The life forms of plants and statistical plant
geography. Oxford: Clarendon Press. 632 p. [2843]
87. Ritchie, J. C. 1955. A natural hybrid in Vaccinium I. The structure,
performance, and chorology of the cross Vaccinium intermedium Ruthe. New
Phytology. 54: 49-67. [9014]
88. Ritchie, J. C. 1955. Biological flora of the British Isles: Vaccinium
vitus-idaea L. Journal of Ecology. 43: 701-708. [9025]
89. Ritchie, J. C. 1957. The vegetation of northern Manitoba. II. A prisere
on the Hudson Bay lowlands. Ecology. 38(3): 429-435. [10552]
90. Robuck, O. Wayne. 1985. The common plants of the muskegs of southeast
Alaska. Miscellaneous Publication/July 1985. Portland, OR: U.S.
Department of Agriculture, Forest Service, Pacific Northwest Forest and
Range Experiment Station. 131 p. [11556]
91. Roland, A. E.; Smith, E. C. 1969. The flora of Nova Scotia. Halifax, NS:
Nova Scotia Museum. 746 p. [13158]
92. Rowe, J. S.; Scotter, G. W. 1973. Fire in the boreal forest. Quaternary
Research. 3: 444-464. [72]
93. Schwartz, Charles C.; Regelin, Wayne L.; Franzmann, Albert W. 1988.
Estimates of digestibility of birch, willow, and aspen mixtures in
moose. Journal of Wildlife Management. 52(1): 33-37. [4535]
94. Shaver, Gaius R. 1986. Woody stem production in Alaskan tundra shrubs.
Ecology. 67(3): 660-669. [4928]
95. Shaw, George. 1981. Concentrations of twenty-eight elements in fruiting
shrubs downwind of the smelter at Flin Flon, Manitoba. Environmental
Pollution (Series A). 25(3): 197-209. [10794]
96. Sirois, Luc; Payette, Serge. 1989. Postfire black spruce establishment
in subarctic and boreal Quebec. Canadian Journal of Forestry Research.
19: 1571-1580. [10110]
97. Sheard, J. W. 1986. Distribution of uranium series radionuclides in
upland vegetation of northern Saskatchewan. I. Plant and soil
concentrations. Canadian Journal of Botany. 64(11): 2446-2452. [10680]
98. Smith, D. W. 1962. Ecological studies of Vaccinium species in Alberta.
Canadian Journal of Plant Science. 42: 82-90. [7004]
99. Soper, James H.; Heimburger, Margaret L. 1982. Shrubs of Ontario. Life
Sciences Misc. Publ. Toronto, ON: Royal Ontario Museum. 495 p. [12907]
100. Strong, W. L.; LaRoi, G. H. 1986. A strategy for concurrently monitoring
the plant water potentials of spatially separate forest ecosystems.
Canadian Journal of Forest Research. 16(2): 346-351. [10805]
101. Trajkovski, Viktor. 1987. Facts about lingonberries (cowberries,
partridgeberries). Fruit Varieties Journal. 41(1): 39. [9601]
102. Uggla, Evald. 1959. Ecological effects of fire on north Swedish forests.
Stockholm, Sweden: Almqvist and Wiksells. 18 p. [9911]
103. U.S. Department of Agriculture, Natural Resources Conservation Service.
2018. PLANTS Database, [Online]. U.S. Department of Agriculture, Natural
Resources Conservation Service (Producer). Available: https://plants.usda.gov/.
[34262]
104. Van Cleve, Keith; Dyrness, C. T. 1983. Introduction and overview of a
multidisciplinary research project: the structure and function of a
black spruce(Picea mariana) forest in relation to other fire-affected
taiga ecosystems Canadian Journal of Forest Research. 13: 695-702. [7883]
105. Van Dersal, William R. 1938. Native woody plants of the United States,
their erosion-control and wildlife values. Washington, DC: U.S.
Department of Agriculture. 362 p. [4240]
106. Vander Kloet, S. P. 1988. The genus Vaccinium in North America.
Publication 1828. Ottawa: Research Branch, Agriculture Canada. 201 p.
[11436]
107. Vander Kloet, S. P. 1989. Typification of some North American Vaccinium
species names. Taxon. 38: 129-134. [8918]
108. Viereck, Leslie A. 1979. Characteristics of treeline plant communities
in Alaska. Holarctic Ecology. 2: 228-238. [8251]
109. Viereck, Leslie A. 1982. Effects of fire and firelines on active layer
thickness and soil temperatures in interior Alaska. In: Proceedings, 4th
Canadian permafrost conference; 1981 March 2-6; Calgary, AB. The Roger
J.E. Brown Memorial Volume. Ottawa, ON: National Research Council of
Canada: 123-135. [7303]
110. Viereck, L. A. 1983. The effects of fire in black spruce ecosystems of
Alaska and northern Canada. In: Wein, Ross W.; MacLean, David A., eds.
The role of fire in northern circumpolar ecosystems. New York: John
Wiley and Sons Ltd.: 201-220. [7078]
111. Viereck, L. A.; Dyrness, C. T. 1979. Ecological effects of the
Wickersham Dome Fire near Fairbanks, Alaska. Gen. Tech. Rep. PNW-90.
Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific
Northwest Forest and Range Experiment Station. 71 p. [6392]
112. Viereck, L. A.; Dyrness, C. T.; Batten, A. R.; Wenzlick, K. J. 1992. The
Alaska vegetation classification. Gen. Tech. Rep. PNW-GTR-286. Portland,
OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest
Research Station. 278 p. [2431]
113. Viereck, L. A.; Foote, Joan; Dyrness, C. T.; [and others]. 1979.
Preliminary results of experimental fires in the black spruce type of
interior Alaska. Res. Note PNW-332. Portland, OR: U.S. Department of
Agriculture, Forest Service, Pacific Northwest Forest and Range
Experiment Station. 27 p. [7077]
114. Viereck, Leslie A.; Little, Elbert L., Jr. 1972. Alaska trees and
shrubs. Agric. Handb. 410. Washington, DC: U.S. Department of
Agriculture, Forest Service. 265 p. [6884]
115. Viereck, Leslie A.; Schandelmeier, Linda A. 1980. Effects of fire in
Alaska and adjacent Canada--a literature review. BLM-Alaska Tech. Rep.
6. Anchorage, AK: U.S. Department of the Interior, Bureau of Land
Mangement, Alaska State Office. 124 p. [7075]
116. Wein, Ross W.; Bliss, L. C. 1973. Changes in Arctic Eriophorum tussock
communities following fire. Ecology. 54(4): 845-852. [9827]
117. West, Stephen D. 1982. Dynamics of colonization and abundance in central
Alaskan populations of the northern red-backed vole, Clethrionomys
rutilus. Journal of Mammalogy. 63(1): 128-143. [7300]
118. Wolff, Jerry O. 1978. Food habits of snowshoe hare in interior Alaska.
Journal of Wildlife Management. 42(1): 148-153. [7443]
119. Wright, John M. 1981. Response of nesting lapland longspurs (Calcarius
lapponicus) to burned tundra on the Seward Peninsula. Arctic. 34(4):
366-369. [7885]
120. Zoltai, S. C.; Tarnocai, C. 1971. Properties of a wooded palsa in
northern Manitoba. Arctic and Alpine Research. 3(2): 115-129. [9778]
121. Stickney, Peter F. 1989. Seral origin of species originating in northern
Rocky Mountain forests. Unpublished draft on file at: U.S. Department of
Agriculture, Forest Service, Intermountain Research Station, Fire
Sciences Laboratory, Missoula, MT; RWU 4403 files. 7 p. [20090]
122. U.S. Department of the Interior, National Biological Survey. [n.d.]. NP
Flora [Data base]. Davis, CA: U.S. Department of the Interior, National
Biological Survey. [23119]
123. Dyrness, C. T.; Norum, Rodney A. 1983. The effects of experimental fires
on black spruce forest floors in interior Alaska. Canadian Journal of
Forest Research. 13: 879-893. [7299]
124. Zasada, John C.; Norum, Rodney A.; Teutsch, Christian E.; Densmore, Roseann.
1987. Survival and growth of planted black spruce, alder, aspen and willow
after fire on black spruce/feather moss sites in interior Alaska. The
Forestry Chronicle. 63(2): 84-88. [85354]
125. Zasada, John C.; Norum, Rodney A.; Van Veldhuizen, Robert M.; Teutsch, Christian
E. 1983. Artificial regeneration of trees and tall shrubs in experimentally
burned upland black spruce/feather moss stands in Alaska. Canadian Journal of
Forest Research. 13: 903-913. [6991]
FEIS Home Page