Index of Species Information
SPECIES: Rubus idaeus
|
|
 |
| American red raspberry. Image by Mary Ellen (Mel) Harte, Bugwood.org. |
Introductory
SPECIES: Rubus idaeus
AUTHORSHIP AND CITATION:
Tirmenstein, D. 1990. Rubus idaeus. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station,
Fire Sciences Laboratory (Producer). Available:
https://www.fs.usda.gov/database/feis/plants/shrub/rubida/all.html [].
Revisions:
On 21 August 2018, the common name of this species was changed in FEIS
from: red raspberry
to: American red raspberry. Images were also added.
ABBREVIATION:
RUBIDA
SYNONYMS:
R. idaeus subsp. sachalinensis (Levl.) Focke = R. i. subsp. strigosus
NRCS PLANT CODE:
RUID
RUIDI
RUIDS2
COMMON NAMES:
American red raspberry
black-haired red raspberry
brilliant red raspberry
raspberry
red raspberry
smoothleaf red raspberry
wild raspberry
wild red raspberry
grayleaf raspberry
TAXONOMY:
The scientific name of American red raspberry is Rubus idaeus L. There
are two subspecies [53]:
Rubus idaeus subsp. idaeus, American red raspberry
Rubus idaeus subsp. strigosus (Michx.) Focke, grayleaf raspberry
Numerous American red raspberry hybrids have been reported, although many are
infertile [43,104]. This shrub hybridizes with many species in the
Rubus genus including R. arcticus, R. ursinus, R. occidentalis, R.
rubrisetus, and R. odoratus [49,55,68,104]. American red raspberry has
hybridized with thimbleberry (R. parviflorus) in the laboratory [49].
LIFE FORM:
Shrub
FEDERAL LEGAL STATUS:
No special status
OTHER STATUS:
NO-ENTRY
DISTRIBUTION AND OCCURRENCE
SPECIES: Rubus idaeus
GENERAL DISTRIBUTION:
American red raspberry occurs throughout most of the temperate regions of the
world [20]. In North America it grows from Alaska through Canada to
Newfoundland, southward to North Carolina and Tennessee in the East, and
to Arizona, California, and northern Mexico in the West [36,93,98]. The
native American red raspberry is Rubus idaeus subsp. strigosus [36].
R. i. subsp. idaeus grows across northern Europe to northwestern Asia
[36]. It is cultivated in Hawaii [109] and throughout much of North
America and has naturalized in many locations [36].
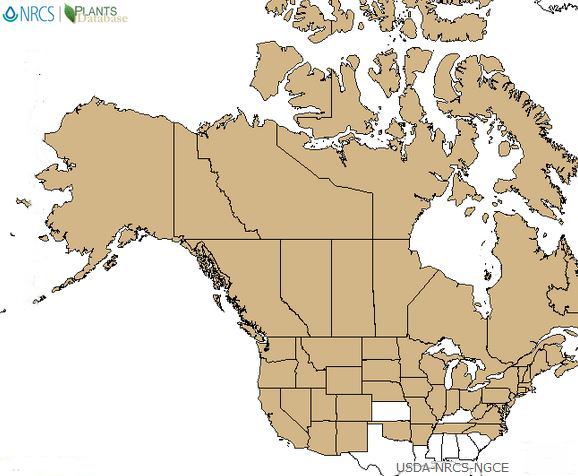 |
| Distribution of American red raspberry in North America. Map courtesy of USDA, NRCS. 2018. The PLANTS Database.
National Plant Data Team, Greensboro, NC [2018, August 21] [90]. |
ECOSYSTEMS:
FRES10 White - red - jack pine
FRES11 Spruce - fir
FRES15 Oak - hickory
FRES17 Elm - ash - cottonwood
FRES18 Maple - beech - birch
FRES19 Aspen - birch
FRES20 Douglas-fir
FRES21 Ponderosa pine
FRES22 Western white pine
FRES23 Fir - spruce
FRES24 Hemlock - Sitka spruce
FRES25 Larch
FRES26 Lodgepole pine
FRES28 Western hardwoods
FRES34 Chaparral - mountain shrub
FRES35 Pinyon - juniper
FRES37 Mountain meadows
FRES38 Plains grasslands
FRES39 Prairie
FRES44 Alpine
STATES:
AK AL AZ AR CA CO CT DE GA HI
ID IL IN IA KY ME MD MA MI MN
MO MT NV NH NJ NM NY NC ND OH
OR PA RI SC SD TN UT VA WA WV
WI WY AB BC MB NB NF ON PQ SK
MEXICO
BLM PHYSIOGRAPHIC REGIONS:
1 Northern Pacific Border
2 Cascade Mountains
3 Southern Pacific Border
5 Columbia Plateau
8 Northern Rocky Mountains
9 Middle Rocky Mountains
10 Wyoming Basin
11 Southern Rocky Mountains
12 Colorado Plateau
13 Rocky Mountain Piedmont
14 Great Plains
15 Black Hills Uplift
16 Upper Missouri Basin and Broken Lands
KUCHLER PLANT ASSOCIATIONS:
K001 Spruce - cedar - hemlock forest
K002 Cedar - hemlock - Douglas-fir forest
K004 Fir - hemlock forest
K011 Western ponderosa forest
K012 Douglas-fir forest
K014 Grand fir - Douglas-fir forest
K015 Western spruce - fir forest
K016 Eastern ponderosa forest
K017 Black Hills pine forest
K018 Pine - Douglas-fir forest
K020 Spruce - fir - Douglas-fir forest
K021 Southwestern spruce - fir forest
K023 Juniper - pinyon woodland
K025 Alder - ash forest
K037 Mountain mahogany - oak scrub
K052 Alpine meadows and barren
K064 Grama - needlegrass - wheatgrass
K067 Wheatgrass - bluestem - needlegrass
K074 Bluestem prairie
K081 Oak savanna
K093 Great Lakes spruce - fir forest
K095 Great Lakes pine forest
K096 Northeastern spruce - fir forest
K097 Southeastern spruce - fir forest
K098 Northern floodplain forest
K102 Beech - maple forest
K106 Northern hardwoods
K107 Northern hardwoods - fir forest
K108 Northern hardwoods - spruce forest
SAF COVER TYPES:
1 Jack pine
5 Balsam fir
12 Black spruce
13 Black spruce - tamarack
15 Red pine
16 Aspen
17 Pin cherry
18 Paper birch
21 Eastern white pine
22 White pine - hemlock
25 Sugar maple - beech - yellow birch
30 Red spruce - yellow birch
31 Red spruce - sugar maple - beech
32 Red spruce
33 Red spruce - balsam fir
35 Paper birch - red spruce - balsam fir
37 Northern white cedar
39 Black ash - American elm - red maple
42 Bur oak
60 Beech - sugar maple
107 White spruce
108 Red maple
109 Hawthorn
201 White spruce
202 White spruce - paper birch
204 Black spruce
210 Interior Douglas-fir
211 White fir
212 Western larch
213 Grand fir
215 Western white pine
217 Aspen
218 Lodgepole
222 Black cottonwood - willow
224 Western hemlock
226 Coastal true fir - hemlock
227 Western redcedar - western hemlock
228 Western redcedar
229 Pacific Douglas-fir
230 Douglas-fir - western hemlock
235 Cottonwood - willow
236 Bur oak
237 Interior ponderosa pine
239 Pinyon - juniper
252 Paper birch
253 Black spruce - white spruce
254 Black spruce - paper birch
HABITAT TYPES AND PLANT COMMUNITIES:
American red raspberry is well represented in many plant communities throughout
North America. It grows within the understory of many quaking aspen
(Populus tremuloides), mixed conifer, cottonwood (Populus spp.), cedar
(Thuja spp.)-hemlock (Tsuga spp.), ponderosa pine (Pinus ponderosa),
spruce (Picea spp.)-fir (Abies spp.), and Douglas-fir (Pseudotsuga
menziesii) forests of the West [6,23,37]. In the Lake States and
Northeast, American red raspberry frequently grows in old-field communities, or in
association with jack pine (Pinus banksiana), white spruce (Picea
glauca), black spruce (P. mariana), red spruce (P. rubens), Atlantic
white-cedar (Thuja occidentalis), balsam fir (Abies balsamea), aspen
(Populus spp.), beech (Fagus spp.), maple (Acer spp.), red pine (Pinus
resinosa), and eastern white pine (P. strobus) [1,3,29,30,34,41,40,77].
It is a common component of northern hardwood forests and often assumes
dominance on sites which have been subject to windthrow, fire, or timber
harvest [100]. American red raspberry is a prominent component of many taiga
communities in Alaska [25] and the Canadian North.
Associated species: American red raspberry grows with a wide variety of plants
across its extensive geographic range. The following species are
particularly common plant associates [1,40,12,13,95]: Canada beadruby
(Maianthemum canadense), thimbleberry, bunchberry (Cornus canadensis),
huckleberry (Vaccinium spp.), fireweed (Epilobium angustifolium),
bluejoint reedgrass (Calamagrostis canadensis), kinnikinnick
(Arctostaphylos uva-ursi), Virginia strawberry (Fragaria virginiana),
green alder (Alnus viridis subsp. crispa), twinflower (Linnaea borealis),
sedges (Carex spp.), prickly rose (Rosa acicularis), twinberry (Lonicera
spp.), lowbush blueberry (Vaccinium angustifolium), bog Labrador tea
(Ledum groenlandica), red currant (Ribes triste), highbush cranberry
(Viburnum edule), and red-osier dogwood (Cornus sericea).
American red raspberry occurs as a dominant in a number of plant communities. It
has been included as a codominant in rocky, high elevation alpine scree
communities with Colorado columbine (Aquilegia coerulea) and
littleflower alumroot (Heuchera parvifolia). American red raspberry has been
listed as an indicator or dominant member of a plant community in the
following publications:
Plant associations of Region Two: Potential plant communities of
Wyoming, South Dakota, Nebraska, Colorado, and Kansas [51]
Habitat types on selected parts of the Gunnison and Uncompahgre National
Forests [56]
MANAGEMENT CONSIDERATIONS
SPECIES: Rubus idaeus
IMPORTANCE TO LIVESTOCK AND WILDLIFE:
Raspberries provide food and cover for a wide range of wildlife species
[10,100]. Some herbivores browse raspberry, but in general, it offers
relatively poor forage. American red raspberry is browsed by moose in Alaska
but is not considered to be of primary importance [79]. In some
locations, deer, rabbits, mountain beaver, and elk eat the foliage of
raspberries [14,91]. Porcupine and beaver occasionally consume buds,
twigs, or cambium of species within the genus Rubus [91]. However,
thorns generally prevent excessive wildlife use of American red raspberry [95].
In general, raspberries have little forage value for domestic livestock
[91].
Fruits of many species within the genus Rubus are eaten by ruffed
grouse, blue grouse, sharp-tailed grouse, ring-necked pheasant, greater
prairie chicken, California quail, northern bobwhite, gray catbird,
northern cardinal, yellow-breasted chat, American robin, thrushes,
towhees, brown thrasher, orchard oriole, summer tanager, pine grosbeak,
gray (Hungarian) partridge, and band-tailed pigeon [14,91]. Mammals
such as the coyote, raccoon, black bear, common opossum, squirrels,
Townsend's chipmunk, skunks, red fox, and gray fox also seek out the
fruits of many raspberries [14,91]. The eastern chipmunk, western
chipmunk, deer mice, and grizzly bear consume American red raspberry fruit where
available [59,105]. Flowers of American red raspberry provide nutritious food
for bees [40].
PALATABILITY:
American red raspberry browse appears to be relatively unpalatable to most
ungulates. However, the fruits are highly palatable to many birds and
mammals. The degree of use shown by livestock and wildlife species for
American red raspberry is rated as follows [23]:
CO MT ND UT WY
Cattle poor poor poor fair poor
Sheep poor fair fair good fair
Horses poor poor poor poor poor
Pronghorn poor ---- ---- poor poor
Elk ---- poor ---- fair fair
Mule deer ---- fair ---- good fair
White-tailed deer fair ---- ---- ---- ----
Small mammals good ---- ---- good fair
Small nongame birds poor ---- ---- good fair
Upland game birds ---- ---- ---- good fair
Waterfowl ---- ---- ---- poor poor
NUTRITIONAL VALUE:
Browse: American red raspberry browse is rated as poor in both energy and
protein value [23]. Nitrogen, phosphorus, and potassium concentrations
are highest in young leaves but decrease as leaves mature [46].
Conversely, calcium and magnesium concentrations are generally highest
in mature leaves but lowest in young, developing leaves [46]. Zinc
typically increases through the growing season whereas manganese
decreases [46]. Levels of nitrogen, phosphorus, potassium, and calcium
generally decline as the growing season progresses but may increase in
the fall if additional rainfall allows plants to resume growth [46].
Fruit: Raspberry fruits are sweet and contain relatively high amounts
of both mono and disaccharides [88]. Relative glucose, starch, and
sugar content has been documented for a number of American red raspberry
cultivars [16].
COVER VALUE:
Dense American red raspberry thickets serve as favorable nesting habitat for many
small birds [14]. Small mammals such as rabbits and squirrels also find
shelter in raspberry thickets [91]. The degree to which American red raspberry
provides environmental protection during one or more seasons is rated as
follows [23]:
CO UT WY
Pronghorn ---- poor poor
Elk ---- poor poor
Mule deer ---- poor poor
White-tailed deer ---- ---- poor
Small mammals fair fair fair
Small nongame birds ---- fair fair
Upland game birds ---- good fair
Waterfowl ---- poor poor
VALUE FOR REHABILITATION OF DISTURBED SITES:
Some ecotypes of American red raspberry have value in reclamation [95]. Suitable
ecotypes are rated as having low to moderate value for short-term
revegetation, and at least moderate value for long-term revegetation
projects [23]. American red raspberry exhibits good potential for erosion
control on some sites [10,91,95]. It has been successfully used to
stabilize roadcuts and other disturbed sites in Utah and to revegetate
bare soils in subalpine zones of Colorado [95]. American red raspberry is
recommended for revegetation projects on well-drained sites in interior
Alaska where maximum spacing of 3.3 feet by 3.3 feet (1 meter x 1 meter)
is suggested [95]. Natural seedling establishment has been observed on
many types of harsh sites, such as on tailings and surface soil of oil
sand extraction plants in northern Alberta [95]. American red raspberry is
capable of establishing on acidic tailings which have been treated with
lime and on tar sands [95].
Propagation: American red raspberry can be propagated through leaf bud cuttings,
"rooted handles," stem cuttings, or root cuttings (suckers)
[24,67,89,95]. Success of establishment through root cuttings varies
according to the cultivar and planting date [89]. However, root cutting
success has ranged up to 60 percent in experimental tests [89]. Correct
choice of planting dates and techniques are important and significantly
influence subsequent growth and establishment [14,89]. In vitro
micropropagation techniques have also been developed for mass production
of American red raspberry [97].
American red raspberry seedlings may be transplanted, or seed may be sown
directly onto disturbed sites. Seed which has been scarified can be
successfully planted in the late summer or fall [10]. Cold treatment is
not required for fall seedings. Previously stratified and scarified
seed can be planted in the spring [10]. Good results have been obtained
after seeds were planted with a drill and covered with 1/8 to 3/16 inch
(0.3-0.5 cm) of soil. Cleaned seed averages approximately 328,000 per
pound (722,467/kg) [10]. Detailed information is available on
appropriate methods to obtain and plant American red raspberry seed [95].
OTHER USES AND VALUES:
The American red raspberry was traditionally an important food of many Native
American peoples. It was eaten fresh or preserved for winter use [66].
Approximately 0.27 quarts (250 ml) of wild American red raspberry fruit can be
hand-harvested within 30 minutes in good stands [66]. The fruit, bark
of roots, and stems of raspberries have been used to make various
medicinal preparations [10].
The unique edible fruit of the American red raspberry is delicious fresh or
preserved. Raspberries make excellent jams and jellies [93] and provide
flavorful additions to pies and other baked goods, candies, and dairy
products such as yogurt or ice cream. Raspberry tea is commercially
available and good although mild in flavor. The raspberry industry in
North America is a growing, multimillion dollar business [63]. Five
primary regions produce most of the raspberries grown commercially in
North America [63]:
1) Northeast-Atlantic Provinces: southern Quebec through
Pennsylvania
2) Central Atlantic Region: Maryland to South Carolina, eastern
Kentucky and Tennessee, northern Georgia and Alabama
3) Central Great Lakes Region: Michigan, southern Ontario, Indiana,
Illinois, Ohio to Iowa, Missouri, western Kentucky and
Tennessee
4) Prairie States Region: Minnesota, southern Manitoba, eastern North
and South Dakota, and Wisconsin
5) Pacific Northwest: southern British Columbia, western Washington
and Oregon
Cultivars: Many cultivars have been developed to meet the needs of
raspberry growers in a variety of climatic situations. Most are derived
from the European subspecies idaeus [98]. Desirable traits for red
raspberry cultivars include spinelessness, winter hardiness, high fruit
yields, resistance to disease, perennial stems, and primocane (or
autumn) fruiting [49]. Reviews of particular cultivars document the
extreme plasticity of this species and consider the suitability of each
to various geographic locations [20,20,22,69,83,67,63,97,16,48].
Cultivars exhibit great genetic variation in time of flowerbud
initiation, number of drupelets produced per fruit, time of fruit
ripening, amount and timing of root suckering, length of dormancy,
winter hardiness, fruit yield, and disease resistance
[19,22,48,67,69,71,83,92]. Consequently, care should be taken to select
cultivars with desirable traits which would enhance suitability for
growth in a specific location [20]. The commonly cultivated loganberry
may have been derived from a American red raspberry-trailing blackberry hybrid
[17].
Commercial cultivation: A wide array of studies detail commercial
propagation of the American red raspberry. Traditional techniques include hill
culture of canes (stems), removal of weeds, and elimination of intercane
suckers to increase fruit yield [64]. Older and weaker canes may be
mowed or otherwise pruned annually to improve yield, enhance access to
fruit, and to maintain the general health of the cane [67,102]. Trends
in American red raspberry propagation include increasing mechanization [63].
Various cultivation techniques have been shown to improve fruit yields
[16,65,72]. In some instances, application of nitrogen fertilizers can
increase both cane growth and the number of flowers produced per node
[67]. However, in other situations fertilizers appear to be of little
benefit [33]. Following the addition of nitrogen fertilizer, Lawson and
Waister [65] observed increased yields for two years, little effect
during the third year, and decreased yields during the next two years.
Similarly, irrigation appears to increase yields in some locations while
having little effect elsewhere [72].
OTHER MANAGEMENT CONSIDERATIONS:
Competition: American red raspberry typically increases dramatically after fire
or timber harvest [27,39]. In many areas this shrub can compete
vigorously with conifer seedlings for light, moisture, nutrients, and
space [30,34,62,74]. Dense thickets of American red raspberry reportedly
suppress the growth of balsam fir (Abies balsamea) and spruce (Picea
spp.) seedlings after spruce-fir forests of northern Maine are clearcut
[30] and after timber harvest in the boreal forests of Ontario [82].
Raspberries also compete effectively with jack pine (Pinus banksiana)
and red pine (P. resinosa) following timber harvest in northeastern
Minnesota and Manitoba [3,74]. Graber and Thompson [34] observed that
relatively few American red raspberry seeds are present within the soil of
northeastern hardwood forest harvested at 100-year intervals. However,
in forests harvested at more frequent intervals, large numbers of red
raspberry seed are present and massive simultaneous germination results
in intense competition with conifer seedlings [34].
Chemical control: American red raspberry is susceptible to a number of
herbicides [9]. Glyphosate is commonly used as a mid-to-late summer
foliar spray [82]. A number of herbicides have been suggested for use
in reducing weeds in cultivated American red raspberry patches [8].
BOTANICAL AND ECOLOGICAL CHARACTERISTICS
SPECIES: Rubus idaeus
GENERAL BOTANICAL CHARACTERISTICS:
American red raspberry is a deciduous, erect or arching, thicket-forming shrub
which grows from 1.6 to 9.8 feet (0.5-3 m) in height [36,80,86,93]. The
total height and extent of growth is largely attributable to climatic
factors [101]. Woody stems are bristly or prickly with shreddy,
exfoliating yellow-brown bark [36,93]. Leaves are alternate and
pinnately compound in leaflets of three to five [86,93]. Leaves are
green and glabrous to hairy above but white or gray, hairy to glabrate
and greenish beneath [98]. Small showy perfect white flowers are
borne in clusters of one to four in a compound cyme [36,55,80,93].
Fruit of the American red raspberry is made up of many to several, red or
pinkish-purple drupelets [80,98]. Aggregates of drupelets are commonly
referred to as a "berry."
American red raspberry is made up of mostly biennial canes (stems) on a
long-lived perennial rootstock [45,99]. This rootstock initially forms
from seedling establishment or the production of root suckers [99],
which gradually separate from parent plants as the connecting root
tissue dies [45]. Sterile first-year stems, or primocanes, develop from
buds at or below the ground surface and generally bear only leaves
[36,99]. During the second year, lateral branches, known as floricanes,
develop in the axils of the primocanes which produce both leaves and
fruit [36,100]. A "typical" raspberry rootstock system is made up of at
least one floricane and several primocanes [99]. It should be noted
that some commercially grown strains of American red raspberry are
primocane-fruiting; that is, they are capable of bearing fruit during
the fall of the first year of development [21,92]. Primocane-fruiting
appears to be absent entirely or represents an atypical situation in
native-growing populations of American red raspberry.
RAUNKIAER LIFE FORM:
Hemicryptophyte
REGENERATION PROCESSES:
American red raspberry reproduces through seed and also regenerates vegetatively.
It is capable of forming dense thickets through sprouting. Reproductive
versatility is well represented in the Rubus genus, with sexual
reproduction, parthenogenesis (development of the egg without
fertilization), pseudogamy (a form of apomixis in which pollination is
required), and parthenocarpy (production of fruit without fertilization)
occurring widely [17]. The following types of reproduction have been
documented within the genus: (1) sexual reproduction, (2) nonreduction
at meiosis on the female, male, or both sides, (3) apomixis with
segregation, (4) apomixis without segregation, and (5) haploid
parthenogenesis [17]. These modes of asexual reproduction are important
because they help contribute to the vigorous, aggressive spread of red
raspberry.
American red raspberry is capable of vigorous sprouting after disturbance [18]
but also expands in clonal area through vegetative regeneration
[95,100]. Natural vegetative regeneration occurs through root sprouts
or "suckers" [95,100,101], "stolons" [95], "rhizomes" [39,52], and basal
stem buds or root crowns [45,95,101]. The precise mode of vegetative
regeneration depends on the type and severity of disturbance. Dense
raspberry thickets form from the roots or stems of parent plants which
separate to form individual plants with the deterioration of connecting
tissue [45]. American red raspberry allocates most energy to vegetative
regeneration on recently disturbed sites with favorable growing
conditions [99]. With time, initially elevated nutrient levels decline,
and shading increases. As growing conditions deteriorate, American red raspberry
shifts its reproductive effort to the production of large numbers of
seed [39,100].
American red raspberry sprouts readily from portions of aboveground stems which
survive disturbance [52]. Many raspberry species are capable of rooting
from the stem nodes, and layering has been widely reported in the red
raspberry [95]. This shrub is also capable of sprouting from axillary
buds located "well above the ground level" [45]. Root crown or stembase
sprouting is an important regenerative mode, which in the raspberry
gives rise to biennial stems even in the absence of disturbance [36,45].
American red raspberry typically sprouts from the root crown if aerial foliage is
cut late in the growing season [95]. In related species such as
salmonberry (R. spectabilis), apical dominance exerted by extant
root crowns inhibits sprouting from belowground structures such as roots
or rhizomes [106].
Root "suckering" is a normal, on-going process in American red raspberry stands
[45,101]. However, particularly vigorous root suckering is often
observed after the aboveground vegetation is damaged or destroyed. This
shrub regenerates from buds located on the larger main roots as well as
those present on lateral roots which are often located fairly close to
the soil surface [52,101]. The mean depth of these underground
regenerative structures (root buds) was estimated at 2.4 inches (6 cm)
in a New Brunswick study [28]. During the first 2 to 3 years after
establishment, root suckers fill in spatial gaps in the clone [100].
Root sucker mortality is generally high during the third and fourth
years because of intense intraspecific competition for sunlight, space,
and nutrients which result in "self thinning" of stands [45,100].
Suckering ability declines with age, with production decreasing from an
average of 1.5 per square foot (16.0/sq m) in 3-year-old stands to 0.77
per square foot (8.25/sq m) in 4-year-old stands [100]. Although
relatively few root suckers actually reach the canopy, survival rates of
those that do is high [100]. Most root suckers live for only 1 or 2
months [100]. Several researchers report that American red raspberry is capable
of sprouting from rhizomes after fire or other disturbance [39,52].
However, others have observed that American red raspberry lacks rhizomes with any
regenerative capability [28]. The term "rhizome" may have been loosely
applied to rhizomelike roots which do possess the ability to sprout.
Geographic or genetic differences in American red raspberry morphology and
physiology are also possible.
Seed: Immature fruit, commonly referred to as "berries," are pink and
hard [10]. Ripe fruit is generally red, but less commonly white or
yellow [43]. Several to many small individual drupelets form an
aggregate fruit [10,98]. Fruit size appears to be related to soil
moisture [72], although significant genotypic variation has also been
noted in the size and number of fruits produced annually [22].
Decreased stored nutrient availability and water stress can influence
overall fruit production [16]. It is estimated that 70 to 90 percent of
American red raspberry flowers eventually mature into fruit which results in an
abundance of seed [99]. Whitney [100] observed that 77 percent of all
plants flowered, with 85 percent of those flowering producing seed.
Most species of raspberry produce good seed crops nearly every year
[10], but seed production does vary annually in the American red raspberry
according to climatic factors and the age of the cane. Whitney [100]
observed average seed production of 65 seeds per square foot (700 seeds/
sq m) in 2-year-old canes, with maximum production of 1,301 seeds per
square foot (14,000 seeds/sq m) in 4-year-old canes. Annual seed
production averaged 604 seeds per square foot (6,500 seeds/sq m) over a
4-year period [100].
Pollination: American red raspberry is primarily pollinated by bees, although
flies and beetles also pollinate some flowers [40]. Under natural
conditions, it is almost exclusively self-incompatible [55] which
contributes to morphological variability.
Germination: Seed of the American red raspberry is relatively large [31], with
viability averaging up to 92 to 99 percent in laboratory tests [95]. Red
raspberry seeds have a hard, thick, impermeable coat and dormant embryo
[10]. Seeds have the ability to become dormant a second time in
response to environmental factors [50]. Consequently, germination is
often slow. Most raspberry seeds require, as a minimum, warm
stratification at 68 to 86 degrees F (20 to 30 degrees C) for 90 days,
followed by cold stratification at 36 to 41 degrees F (2 to 5 degrees C)
for an additional 90 days [10]. Cold stratification alone is
insufficient to induce germination in American red raspberry [59]. Laboratory
tests indicate that exposure to sulfuric acid solutions or sodium
hyperchlorite prior to cold stratification can improve germination
[10,43,50,95]. Evidence suggests that the digestive enzymes of mammals
can also enhance germination, with seeds eaten by chipmunks and deer
mice exhibiting better germination than untreated seeds [59]. Sowing
seeds at greater depths with subsequent exposure to light can produce
better germination than shallow plantings, presumably because of greater
soil moisture [50]. Results of specific germination tests have been
documented in a number of studies [10,50,59].
Seed banking: American red raspberry amasses large numbers of seed which persist
in the soil until favorable germination conditions are encountered
[31,35,100]. Often, many seeds remain buried in the soil of stands
which lack any sign of the parent plants [31]. American red raspberry seed can
remain viable for 60 to 100 years or more [62,73,100]. Seeds are less
likely to germinate when fresh [50,62], and may reach maximum viability
at 50 to 100 years of age [34]. In a New Hampshire study, approximately
90 percent of Rubus (R. idaeus and R. alleghaniensis) seed germinated
during the first summer after disturbance in 38-, 95-, and >
200-year-old stands, whereas only 60 percent of those in 5-year-old
stands germinated [34]. More than 4,048,583 Rubus seeds per acre (10
million/ha) have been found in the soil of 5-year-old beech (Fagus
spp.)-birch (Betula spp.)-maple (Acer spp.) stands [34]. Numbers
declined to 48,588 per acre (120,000/ha) in 200-year-old stands [34].
Annual reductions in stored seed have been attributed to: (1)
degeneration resulting in death, (2) fungi or animal predation, and (3)
annual germination of some seeds. Fyles [31] reported 237 to 1,883
seeds per foot square (22-175/m sq) in organic soil and 0 to 2,582 per
foot square (0-240 m sq) in mineral soil of upland coniferous forests of
central Alberta. Distribution of germinating seeds by stand age in
beech-birch-maple forests of New Hampshire were as follows [34]:
stand age in years
5 38 95 200 +
#seeds/m sq. 1,016 286 68 12
Seed dispersal: American red raspberry seed is readily dispersed by birds and
mammals [87,100]. After they mature, the highly sought-after fruit
rarely remains on the plants for long [10]. Birds have been observed to
deposit 2,429 to 2,834 viable seeds per acre (6,000 to 7,000/ha)
annually in beech-birch-maple forests of New Hampshire [34]. Mammals
such as mice and chipmunks may be important dispersal agents in some
areas [59].
Seedling establishment: Most seedlings germinate during the first year
after disturbance [99,100] and produce stands which are primarily
even aged. In many instances, as much as 70 to 90 percent of all
individuals establish during the first year after disturbance [100].
Researchers have observed minimal recruitment in the second, third, and
fourth years after fire [100]. Little seedling establishment occurs
beneath the shade of a closed forest canopy [100].
SITE CHARACTERISTICS:
American red raspberry grows across a wide range of sites throughout most of the
world's temperate regions [20]. It commonly occurs in clearings or
borders in boreal forests, in ravines, on bluffs and streambanks of
prairie regions, and on talus or scree above timberline [39,86,93,95].
Soil: Raspberries are tolerant of a wide range of soil pH and texture
but do require adequate soil moisture [14]. American red raspberry grows on
imperfectly to well-drained sandy loam to silty clay loam, but best
growth occurs on moderately well-drained soils [95]. Although red
raspberry grows well on barren and infertile soils, it reportedly has a
relatively high demand for soil nutrients and is most abundant on
nutrient-rich soils [39]. This shrub is moderately tolerant of acidic
soils [95].
Elevation: Generalized elevational ranges for selected locations are as
follows [23,98]:
from 6,500 to 11,700 feet (1,981 to 3,569 m) in CO
2,400 to 7,000 feet (732 to 2,134 m) in MT
5,500 to 9,600 feet (1,676 to 3,420 m) in UT
6,500 to 11,000 feet (1,981 to 3,355 m) in WY
SUCCESSIONAL STATUS:
American red raspberry vigorously invades and colonizes many types of disturbed
sites [62,95,100]. It is generally considered a pioneer or early seral
species [35] which flourishes and completes its life cycle during the
first years after disturbance [100]. This shade-intolerant species
often dominates sites during early successional stages but decreases as
the canopy closes [62,100]. Although the plants themselves remain
prominent for only a relatively brief period, viable seeds can persist
for 60 years or more in the soil or duff [73]. Widespread germination
after disturbance frequently leads to the development of even-aged
stands [100]. In many areas, American red raspberry is absent beneath the canopy
of mature forests but persists in forest openings [39]. Whitney [99]
reports that few stands of American red raspberry persist for longer than 5 to 12
years.
American red raspberry invades black and white spruce stands in Alaska during the
first years after disturbance but declines as taller shrubs and trees
become established [29,39]. In many northern black spruce forests, red
raspberry is present only in early successional stages [29]. On mesic
and submesic sites in sub-boreal forests of British Columbia it
typically increases during the first 10 years after timber harvest or
fire but is virtually eliminated within 14 years because of rapid
increases in shade [39]. American red raspberry often dominates jack pine stands
of Minnesota within 5 years after disturbance [2,39] and subsequently
declines as the canopy develops. In parts of western Montana, red
raspberry initially grows rapidly but begins to decline within 3 to 4
years after disturbance as nutrient levels decrease [18]. American red raspberry
can persist for up to 4 or 5 years in northern hardwood forests as long
as stands remain relatively open [47,73]. In birch-maple forests of New
Hampshire, American red raspberry reaches peak abundance in the second through
fourth years after disturbance [100]. However, it rarely persists for
more than 10 years [100]. American red raspberry is subsequently replaced by
species such as aspen, chokecherry (Prunus spp.), and birch [100].
SEASONAL DEVELOPMENT:
American red raspberry is typically biennial, with each shoot passing through
well-defined phenological stages during its 2-year lifespan [45].
Vegetative shoots develop from the roots or stems of parent plants, or
as seedlings, during the first year [45,99,100,101]. Lateral flowering
stalks (floricanes) are produced during the second year [22,99,100].
Floricanes leaf out early and exhibit rapid growth [99]. After
producing fruit in late summer, the leaves of floricanes senesce and the
cane gradually dies [99]. Stages of the 2-year growth cycle of red
raspberry are detailed below [45]:
YEAR 1
Phases
phase 1: initiation of root buds
phase 2: subterranean suckering
phase 3: emergent suckers; elongation slows or stops
as sucker reaches surface; leaves form a rosette
at or above the soil surface.
phase 4: 1st winter dormancy-most leaves shed
YEAR 2
phase 5: elongating shoot; rapid elongation.
phase 6: initiation of flower buds; shoot stops
elongation at end of growing season.
secondary rosettes form; axillary meristems
initiate flower primordia; dormant fruit buds;
leaves become senescent and fall.
phase 7: breaking dormancy of flower buds; require
cold to break dormancy; buds grow in spring
(some cultivars produce fruit before dormancy).
phase 8: flowering and fruiting; basal buds elongate
into a vegetative replacement shoot which
repeats the biennial cycle.
phase 9: senescence and death. (after fruiting the
shoot dies back "to the position from which
a replacement shoot has grown").
Flowering: Flowerbud initiation is influenced by temperature, genetics
(cultivar), and geographic location [21,48,69,92]. Flowering is also
related to the age and vigor of the plant and the date at which
vegetative growth terminates [16,67]. Flowerbud initiation is triggered
by low temperatures and short days and generally begins in late summer
or autumn [16,21]. Flowerbud initiation can be induced by exposure to
temperatures of 55 degrees F (12.8 degrees C) at 9 hour days or 50
degrees F (10.0 degrees C) at 16 hour days [21]. Although flowerbud
initiation occurs over winter in most red raspberries, initiation in
primocane-fruiting cultivars begins in summer [92]. Bud break typically
occurs in early spring [16]. Evidence suggests that higher spring
temperatures may promote earlier and more rapid flowering [67].
Fruiting: Fruit maturation begins soon after flowering [88]. Timing of
flowerbud initiation largely determines fruiting season [21], although
fruiting dates also vary according to cultivar and geographic location
[21]. Annual variation in fruit ripening has also been reported [21].
Both flowering and fruiting proceeds from the top of the floricane
downward [22]. After maturation, fruit spoils rapidly [88].
Generalized fruiting and flowering dates by geographic location are as
follows [21,23,36,54,80,84,86,88,93]:
location flowering fruiting
AK June-July July-September
AZ June-July -----
BC ----- July-August
CO June-July -----
East ----- July-October
Great Plains May-July -----
MT June-August -----
NC, SC June-August July-August (or later)
ND June -----
New England ---- late June-August
nc Plains June-July July-August
UT May-July -----
WY June-August -----
FIRE ECOLOGY
SPECIES: Rubus idaeus
FIRE ECOLOGY OR ADAPTATIONS:
The life cycle of American red raspberry is integrally associated with
disturbances such as fire. In many areas of vigorous fire suppression,
both plant vigor and abundance have decreased [66]. American red raspberry
typically flourishes, completes its life cycle and declines within the
early years after disturbance [73]. As shade levels increase in the
postfire community and soil nitrate levels drop (generally during the
first 5 years after fire), American red raspberry shifts resource allocation from
vegetative growth to seed production [39,99].
Although the plants themselves soon senesce and die, viable seed
persists for decades [62,73], germinating in great numbers after the
next fire [100] creates favorable conditions for growth and
establishment. Seed is effectively scarified by heat [78,94], and
exposed mineral soil serves as a favorable substrate for early growth
and development [26]. Underground regenerative structures appear to be
well protected from the damaging effects of heat [28,52], and
reestablishment is typically rapid where plants were present in the
preburn community.
FIRE REGIMES:
Find fire regime information for the plant communities in which this
species may occur by entering the species name in the FEIS home page under
"Find Fire Regimes".
POSTFIRE REGENERATION STRATEGY:
Tall shrub, adventitious-bud root crown
Rhizomatous shrub, rhizome in soil
Geophyte, growing points deep in soil
Ground residual colonizer (on-site, initial community)
Initial-offsite colonizer (off-site, initial community)
FIRE EFFECTS
SPECIES: Rubus idaeus
IMMEDIATE FIRE EFFECT ON PLANT:
American red raspberry is described as "resistant" to fire [39,103]. However,
foliage is extremely susceptible to fire-induced mortality [52]. In an
Alberta study, all aboveground stems were completely killed wherever
supplemental fuels contributed to relatively intense fires [52]. Where
fuels were reduced and fires less intense, the stems of many plants were
only partially killed [52]. However, all aerial stems experienced at
least partial mortality, regardless of fire intensity.
Belowground regenerative structures appear to be relatively resistant to
fire [39]. Johnston and Woodard [52] observed belowground mortality
only on plots with high surface fuel loadings (3.94 or 9.65 kg/m sq).
Here, tissue mortality extended as far as 0.4 to 1.2 inches (1-3 cm)
below the duff surface. Raspberry is capable of sprouting from lateral
buds on relatively shallow roots. These roots, which are small and
succulent, are poorly protected by duff and can be damaged by fires of
high intensity and severity [52]. However, at least some regenerative
structures typically grow to 2 inches (5 cm) below the soil surface, and
many are apparently unharmed by fires of even high intensity and
severity [52]. In general, the effects of fire on American red raspberry are
much less pronounced wherever nutrients and water are abundant [39].
The long-lived seed of American red raspberry is generally unharmed by fire when
protected by overlying soil [39,78,94].
DISCUSSION AND QUALIFICATION OF FIRE EFFECT:
In an Alberta study, both dead and live woody stems remained where fuel
loadings of 0.00, 0.17, and 0.87 kg/m sq were recorded [52]. However,
all foliage was completely consumed on plots with fuel loadings of 3.94
and 9.65 kg/m sq [52].
PLANT RESPONSE TO FIRE:
American red raspberry is well adapted to reoccupy a site quickly after fire.
This common "fire follower" is favored by increased amounts of nitrates
present on burned sites and generally exhibits rapid and vigorous
postfire growth through sprouting and/or seedling establishment [4,95].
Vegetative response: American red raspberry typically sprouts readily after fire
wherever present in preburn communities [18,95]. Most belowground
regenerative structures appear to be well protected from the damaging
effects of heat [52]. Postfire sprouting of root buds is commonly
observed. Although more shallow root buds may be damaged or killed by
heat, root bud depths can range from 1.9 to 2.4 inches (5-6 cm) or
greater [28,52] and many escape serious damage. Postfire sprouting from
rhizomes may also occur [39], although a number of researchers have
reported no evidence of any rhizomes with regenerative capabilities in
the American red raspberry [28]. Where light fires damage but do not kill the
aboveground foliage, aerial stems generally sprout and quickly resume
growth [52]. Consequently, American red raspberry is reported to be
"rejuvenated" by fire [103].
Johnston and Woodard [52] reported that fire intensity and severity had
little effect on the sprouting ability of American red raspberry in aspen
communities of east-central Alberta. Both the number of sprouts
produced per plant and total biomass appeared unaffected by fire
intensity and severity. However, the height growth of individual
sprouts was greatest after fires of low severity. Thus, although high
severity fires reduced the rate of sprout growth, they did not affect
the number of sprouts produced by each plant [52].
Seedling establishment: Rapid postfire establishment through on-site
seed is common in the American red raspberry [95]. Long-lived seed, which is
produced in abundance, accumulates in seed banks in the soil or duff
[35,38]. Germination is enhanced by exposure to heat [78,94], and large
numbers of seed germinate soon after disturbance [100]. Mineral soil
creates a favorable seedbed [26] and elevated nitrate levels enhance
early seedling growth. Most germination occurs within the first year
after fire [38].
Limited evidence suggests that fires of high intensity and severity may
promote American red raspberry seedling establishment more than light fires.
Bock and Bock [6] observed vigorous seedling establishment after crown
fires in ponderosa pine forest of the southern Black Hills. However,
large increases in American red raspberry did not occur after lighter, cooler
ground fires in the same area [6,7]. Extremely light fires may provide
insufficient heat scarification and do little to prepare a seedbed.
Postfire recovery: Postfire recovery of American red raspberry is generally
rapid, with vigorous expansion in cover during early seral stages. This
shade-intolerant species [95,100] declines as tree cover increases [29].
In many communities, American red raspberry begins to decline within only 3 or 4
years after fire [18,103]. It is important to note that many variables
can significantly influence the speed of postfire recovery and
subsequent persistence within the community. Such variables may include
season of burn, fire intensity and severity, site characteristics,
genetic variation, and climatic factors. Specific postfire response of
American red raspberry by community is discussed in the 'Successional Status'
slot.
DISCUSSION AND QUALIFICATION OF PLANT RESPONSE:
In some areas, significant differences in postfire recovery of red
raspberry have been noted after fires of varying intensity and severity.
The following data document postfire recovery in ponderosa pine stands
of the southern Black Hills of South Dakota [7,6]:
------------------------------------------------------------------------
# of stems per subplot
fire type preburn 1st year 2nd year 5th year
light burn 0 0 0.07 ---
crown fire --- --- 0.01 116.8
# of individuals
fire type 1st year 2nd year 3rd year 5th year
cool ground fire 0 0 3 ---
hot crown fire --- --- --- 4,672
------------------------------------------------------------------------
For further information on American red raspberry response to fire, see
Fire Case Studies. Hamilton's Research Papers (Hamilton 2006a, Hamilton 2006b)
also provide information on prescribed fire and postfire response
of plant community species, including American red raspberry, that was not
available when this species review was originally written.
FIRE MANAGEMENT CONSIDERATIONS:
Fire generally benefits animals that consume the fruits of species
within the genus Rubus [58].
SPECIES: Rubus idaeus
FIRE CASE STUDY CITATION:
Tirmenstein, D., compiler. 1990. Effects of prescribed fire on American red raspberry
on Elk Island, Alberta. In: Rubus idaeus. In: Fire Effects Information
System, [Online]. U.S. Department of Agriculture, Forest Service,
Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available:
https://www.fs.usda.gov/database/feis/plants/shrub/rubida/all.html#FireCaseStudies [].
REFERENCE:
Johnston, Mark; Woodard, Paul. 1985. The effect of fire severity level
on postfire recovery of hazel and raspberry in east-central Alberta.
Canadian Journal of Botany. 63: 672-677. [52].
SEASON/SEVERITY CLASSIFICATION:
May 5, 1980/variable
STUDY LOCATION:
The study site is located in Elk Island National Park, approximately 23
miles (37 km) east of Edmonton, Alberta.
PREFIRE VEGETATIVE COMMUNITY:
Preburn overstory vegetation was made up of quaking aspen (Populus
tremuloides) and balsam poplar (P. balsamifera), which ranged from 50 to
70 years of age. Understory vegetation was dominated by American red raspberry
(Rubus idaeus), beaked hazelnut (Corylus cornuta), strawberry (Fragaria
spp.), fleabane (Erigeron spp.), and reedgrass (Calamagrostis spp.).
TARGET SPECIES PHENOLOGICAL STATE:
not reported.
SITE DESCRIPTION:
Soil - orthic gray luvisol
Elevation - not reported
Topography - not reported
Size of treated area - 9.9 acres (4 ha)
Weather conditions - dry
FIRE DESCRIPTION:
Seven artificial fuel beds (24 x 24 inches [60 x 60 cm]) were
constructed of varying amounts of excelsior, or excelsior mixed with
white spruce slats. The prescribed head fire did not spread to the beds
because of discontinuous fuels. The beds were ignited with matches
after the passage of the flame front. Specific weather and fire
behavior characteristics were as follows:
Fire weather:
Dry bulb temperature (C) - 14.5
Relative humidity (%) - 33
Wind speed at 10 m (kh/h) - 6
Fine fuel moisture code - 88
Duff moisture code - 57
Drought code - 106
Initial spread index - 4
Buildup index - 58
Fire weather index - 12
Estimated fire behavior characteristics recorded for the seven
fuel beds ignited with matches -
fuel loading flame length frontal fire residence time
(kg/m sq.) (m) intensity (minutes)
(kW/m)
0.17 0.5 57 1.5
0.87 1 258 2
0.87 1 258 2
3.94 1.5 622 4
3.94 1.5 622 4
9.65 1.5 1,162 10
9.65 2.5 1,905 10
FIRE EFFECTS ON TARGET SPECIES:
All aboveground American red raspberry stems were killed where fuels were added.
On fuel-free plots, only portions of the aboveground stems were killed.
Plants sprouted from underground regenerative structures where
aboveground mortality was complete, but from both aboveground and
belowground tissues where portions of the aerial stems were killed.
Depth of underground regenerative structures ranged from 0 to 2 inches
(0-5 cm), with mortality of tissues occasionally extending to 0.4 to 1.2
inches (1-3 cm) below the duff surface. Portions of most deeper
"rhizomes" apparently survived. American red raspberry also reproduces through
lateral buds located on small, shallow, succulent, poorly protected
roots which can be damaged by fires of high intensity.
The number of sprouts produced per plant did not vary according to burn
treatment. Height growth and the number of leaves did vary by
treatment, indicating that American red raspberry recovery may be affected by
fire intensity and severity. Specific recovery rate data for red
raspberry were as follows:
date response fuel loading (kg/m sq.)
parameter 0.00 0.17 0.87 3.94 9.65
July 4 #sprouts/plot 5 4 4 4 5
ht. growth/sprout(cm) 52.0 32.7 17.4 12.2 8.8
avg. # leaves/sprout -- 8 6 5 3
July 22 #sprouts/plot 6 4 5 4 7
ht. growth/sprout(cm) 58.7 36.5 19.5 16.1 12.8
avg. # leaves/sprout -- 8 6 5 5
Aug. 1 #sprouts/plot 3 4 4 4 7
ht. growth/sprout(cm) 58.4 36.8 19.1 16.2 13.5
avg. # leaves/sprout -- 8 6 5 5
Aug. 16 #sprouts/plot 3 4 5 4 7
ht. growth/sprout(cm) 60.0 37.2 21.0 16.7 13.5
avg. # leaves/sprout -- 7 7 6 6
Aug. 30 #sprouts/plot 3 2 4 3 7
ht. growth/sprout(cm) 60.2 37.0 20.8 16.8 13.8
avg. # leaves/sprout -- 7 6 5 6
Mean abovegrd. ovendry biomass
(grams/plot) 2.5 4.1 5.0 4.6 4.4
FIRE MANAGEMENT IMPLICATIONS:
American red raspberry may be somewhat susceptible to high intensity fires
because of shallow, fairly poorly protected root buds. This study
suggests that hot prescribed fires may be useful in reducing red
raspberry in some carefully selected instances.
REFERENCES
SPECIES: Rubus idaeus
REFERENCES:
1. Ahlgren, Clifford E. 1959. Some effects of fire on forest reproduction
in northeastern Minnesota. Journal of Forestry. 57: 194-200. [208]
2. Ahlgren, Clifford E. 1966. Small mammals and reforestation following
prescribed burning. Journal of Forestry. 64: 614-618. [206]
3. Ahlgren, Clifford E. 1976. Regeneration of red pine and white pine
following wildfire and logging in northeastern Minnesota. Journal of
Forestry. 74: 135-140. [7242]
4. Ahlgren, I. F.; Ahlgren, C. E. 1960. Ecological effects of forest fires.
Botanical Review. 26: 458-533. [205]
5. Bernard, Stephen R.; Brown, Kenneth F. 1977. Distribution of mammals,
reptiles, and amphibians by BLM physiographic regions and A.W. Kuchler's
associations for the eleven western states. Tech. Note 301. Denver, CO:
U.S. Department of the Interior, Bureau of Land Management. 169 p.
[434]
6. Bock, Jane H.; Bock, Carl E. 1984. Effects of fires on woody vegetation
in the pine-grassland ecotone of the southern Black Hills. American
Midland Naturalist. 112(1): 35-42. [477]
7. Bock, Jane H.; Bock, Carl E. [n.d.]. Some effects of fire on vegetation
and wildlife in ponderosa pine forests of the southern Black Hills.
Final Report. Contracts CX-1200-9-B034, CX-1200-0-B018, CX-1200-1-B022;
Grant No. RM-80-105 GR. Unpublished report on file with: U.S. Department
of Agriculture, Forest Service, Intermountain Research Station, Fire
Sciences Lab, Missoula, MT. 58 p. [479]
8. Bonanno, A. Richard .x.
1987. Raspberry and blackberry weed management in Michigan. In: 117th
annual report, Michigan State Horticulture Society: 177-182. [7059]
9. Bovey, Rodney W. 1977. Response of selected woody plants in the United
States to herbicides. Agric. Handb. 493. Washington, DC: U.S. Department
of Agriculture, Agricultural Research Service. 101 p. [8899]
10. Brinkman, Kenneth A. 1974. Rubus L. blackberry, raspberry. In:
Schopmeyer, C. S., ed. Seeds of woody plants in the United States.
Agriculture Handbook No. 450. Washington, DC: U.S. Department of
Agriculture, Forest Service: 738-743. [7743]
11. Chrosciewicz, Z. 1976. Burning for black spruce regeneration on a
lowland cutover site in southeastern Manitoba. Canadian Journal of
Forest Research. 6(2): 179-186. [7280]
12. Chrosciewicz, Z. 1978. Slash and duff reduction by burning on clear-cut
jack pine sites in central Saskatchewan. Information Report NOR-X-200.
Edmonton, AB: Forestry Service, Fisheries and Environment Canada,
Northern Forest Research Centre. 12 p. [7288]
13. Chrosciewicz, Z. 1978. Large-scale operational burns for slash disposal
and conifer reproduction in central Saskatchewan. Information Report
NOR-X-201. Edmonton, AB: Forestry Service, Fisheries and Environment
Canada, Northern Forest Research Centre. 11p. [7289]
14. Core, Earl L. 1974. Brambles. In: Gill, John D.; Healy, William M.,
compilers. Shrubs and vines for Northeastern wildlife. Gen. Tech. Rep.
NE-9. Broomall, PA: U.S. Department of Agriculture, Forest Service:
16-19. [8923]
15. Corns, Ian G. W. 1989. Ecosystems with potential for aspen management.
Managing for aspen--a shared responsibility: Proceedings of the Joint
TechnicalSession of the Forest Ecology, Silviculture and Tree
Improvement Forest Management, and Forest Economics and Policy Working
Groups; September 1988; Prince Albert, SK. In: The Forestry Chronicle.
February: 16-22. [6919]
16. Crandall, P. C.; Allmendinger, D. F.; Biderbost, K. A. 1974. Influence
of cane number and diameter, irrigation, and carbohydrate reserves on
the fruit number of red raspberries. Journal of the American
Horticultural Society. 99(6): 524-526. [6652]
17. Crane, M. B. 1940. Reproductive versatility in Rubus. I. Morphology and
inheritance. Journal of Genetics. 40: 109-118. [8443]
18. Crane, Marilyn F. 1982. Fire ecology of Rocky Mountain Region forest
habitat types. Final Report Contract No. 43-83X9-1-884. Missoula, MT:
U.S. Department of Agriculture, Forest Service, Region 1. 272 p. On file
with: U.S. Department of Agriculture, Forest Service, Intermountain
Research Station, Fire Sciences Laboratory, Missoula, MT. [5292]
19. Dale, A. 1979. Varietal differences in the relationships between some
characteristics of red raspberry fruiting laterals and their position on
the cane. Journal of the Horticultural Society. 54(4): 257-265. [6983]
20. Dale, Adam; Daubeny, Hugh A. 1985. Genotype-environmental interactions
involving British and Pacific Northwest red raspberry cultivars.
HortScience. 20(1): 68-69. [6834]
21. Dale, Adam; Daubeny, Hugh A. 1987. Flower-bud initiation in red
raspberry (Rubus idaeus L.) in two environments. Crop Research. 27:
61-66. [6980]
22. Dale, A.; Topham, Pauline B. 1980. Fruiting structure of the red
raspberry: multivariate analysis of lateral characteristics. Journal of
the Horticultural Society. 55(4): 397-408. [6982]
23. Dittberner, Phillip L.; Olson, Michael R. 1983. The plant information
network (PIN) data base: Colorado, Montana, North Dakota, Utah, and
Wyoming. FWS/OBS-83/86. Washington, DC: U.S. Department of the Interior,
Fish and Wildlife Service. 786 p. [806]
24. Doran, William L. 1957. Propagation of woody plants by cuttings.
Experiment Station Bul. No. 491. Amherst, MA: University of
Massachusetts, College of Agriculture. 99 p. [6399]
25. Dyrness, C. T.; Viereck, L. A.; Van Cleve, K. 1986. Fire in taiga
communities of interior Alaska. In: Forest ecosystems in the Alaskan
taiga. New York: Springer-Verlag: 74-86. [3881]
26. Dyrness, C. T.; Viereck, L. A.; Foote, M. J.; Zasada, J. C. 1988. The
effect on vegetation and soil temperature of logging flood-plain white
spruce. Res. Pap. PNW-RP-392. Portland, OR: U.S. Department of
Agriculture, Forest Service, Pacific Northwest Research Station. 45 p.
[7471]
27. Eyre, F. H., ed. 1980. Forest cover types of the United States and
Canada. Washington, DC: Society of American Foresters. 148 p. [905]
28. Flinn, Marguerite A.; Wein, Ross W. 1977. Depth of underground plant
organs and theoretical survival during fire. Canadian Journal of Botany.
55: 2550-2554. [6362]
29. Foote, M. Joan. 1983. Classification, description, and dynamics of plant
communities after fire in the taiga of interior Alaska. Res. Pap.
PNW-307. Portland, OR: U.S. Department of Agriculture, Forest Service,
Pacific Northwest Forest and Range Experiment Station. 108 p. [7080]
30. Fox, Thomas R. 1986. Raspberry (Rubus idaeus L.) competition effects on
balsam fir (Abies balsamea (L.) Mill.) seedlings in northern Maine. Tree
Planter's Notes. 37(2): 20-23. [6825]
31. Fyles, James W. 1989. Seed bank populations in upland coniferous forests
in central Alberta. Canadian Journal of Botany. 67: 274-278. [6388]
32. Garrison, George A.; Bjugstad, Ardell J.; Duncan, Don A.; [and others].
1977. Vegetation and environmental features of forest and range
ecosystems. Agric. Handb. 475. Washington, DC: U.S. Department of
Agriculture, Forest Service. 68 p. [998]
33. Goode, J. E. 1970. Nitrogen nutrition and susceptibility of Malling
Jewel raspberries to infection by spur blight (Didymella applanata).
Plant Pathology. 19: 108-110. [6591]
34. Graber, Raymond E.; Thompson, Donald F. 1978. Seeds in the organic
layers and soil of four beech-birch-maple stands. Res. Pap. NE-401.
Broomall, PA: U.S. Department of Agriculture, Forest
Service,Northeastern Forest Experiment Station. 8 p. [5983]
35. Granstrom, Anders. 1982. Seed banks in five boreal forest stands
originating between 1810 and 1963. Canadian Journal of Botany. 60:
1815-1821. [5940]
36. Great Plains Flora Association. 1986. Flora of the Great Plains.
Lawrence, KS: University Press of Kansas. 1392 p. [1603]
37. Haeussler, S.; Pojar, J.; Geisler, B. M.; [and others]. 1985. A guide to
the interior cedar-hemlock zone, northwestern transitional subzone
(ICHg), in the Prince Rupert Forest Region, British Columbia. Land
Management Report Number 26; ISSN 0702-9861. Victoria, BC: British
Columbia, Ministry of Forests. 263 p. [6930]
38. Hamilton, Evelyn H. 1988. Impacts of prescribed burning on
soil-vegetation relationships in the sub-boreal spruce zone. In: Feller,
M. C.; Thomson, S. M., eds. Wildlife and range prescribed burning
workshop proceedings; 1987 October 27-28; Richmond, BC. Vancouver, BC:
The University of British Columbia, Faculty of Forestry: 171-184.
[3110]
39. Bare, B. Bruce. 1982. The economics of true fir management. In: Oliver,
Chadwick Dearing; Kenady, Reid M., eds. Proceedings of the biology and
management of true fir in the Pacific Northwest symposium; 1981 February
24-26; Seattle-Tacoma. Contribution No. 45. Seattle, WA: University of
Washington, College of Forest Resources: 9-14. [6760]
40. Hansen, Richard W.; Osgood, Eben A. 1983. Insects visiting flowers of
wild red raspberry in spruce-fir forested areas of eastern Maine.
Entomological News. 94(4): 147-151. [7001]
41. Hansen, H. L.; Krefting, L. W.; Kurmis, V. 1973. The forest of Isle
Royale in relation to fire history and wildlife. Tech. Bull. 294;
Forestry Series 13. Minneapolis, MN: University of Minnesota,
Agricultural Experiment Station. 44 p. [8120]
42. Hansen, Paul L.; Hoffman, George R.; Steinauer, Gerry A. 1984. Upland
forest and woodland habitat types of the Missouri Plateau, Great Plains
Province. In: Noble, Daniel L.; Winokur, Robert P., eds. Wooded draws:
characteristics and values for the Northern Great Plains: Symposium
proceedings; 1984 June 12-13; Rapid City, SD. Great Plains Agricultural
Council Publ. No. 111. Rapid City, SD: South Dakota School of Mines and
Technology, Biology Department: 15-26. [1078]
43. Heit, C. E. 1967. Propagation from seed. Part 7: Germinating six
hardseeded groups. American Nurseryman. 125(12): 10-12; 37-41; 44-45.
[1120]
44. Hogdon, A. R.; Steele, Frederic. 1966. Rubus subgenus Eubatus in New
England: a conspectus. Rhodora. 68: 474-513. [6213]
45. Hudson, J. P. 1959. Effects of environment on Rubus idaeus L. I.
Morphology and development of the raspberry plant. Journal of
Horticultural Science. 34: 163-169. [6607]
46. Hughes, Megan; Chaplin, M. H.; Dixon, A. R. 1979. Elemental composition
of red raspberry leaves as a function of time of season and position on
cane. HortScience. 14(1): 46-47. [6833]
47. Hughes, Jeffrey W.; Fahey, Timothy J.; Bormann, F. Herbert. 1988.
Population persistence and reproductive ecology of a forest herb: Aster
acuminatus. American Journal of Botany. 75(7): 1057-1064. [8972]
48. Jennings, D. L.; Cormack, M. R. 1969. Factors affecting the water
content and dormancy of overwintering raspberry canes. Horticultural
Research. 9: 18-25. [6655]
49. Jennings, D. L.; Ingram, Ruth. 1983. Hybrids of Rubus parviflorus
(Nutt.) with raspberry and blackberry, and the inheritance of
spinelessness derived from this species. Crop Research. 23(2): 95-101.
[7029]
50. Jennings, D. L.; Tulloch, M. M. 1964. Studies on factors which promote
germination of raspberry seeds. Journal of Experimental Botany. 16(47):
329-340. [6535]
51. Johnston, Barry C. 1987. Plant associations of Region Two: Potential
plant communities of Wyoming, South Dakota, Nebraska, Colorado, and
Kansas. 4th ed. R2-ECOL-87-2. Lakewood, CO: U.S. Department of
Agriculture, Forest Service, Rocky Mountain Region. 429 p. [3519]
52. Johnston, Mark; Woodard, Paul. 1985. The effect of fire severity level
on postfire recovery of hazel and raspberry in east-central Alberta.
Canadian Journal of Botany. 63: 672-677. [6277]
53. Kartesz, John T. 1994. A synonymized checklist of the vascular flora of
the United States, Canada, and Greenland. Volume II--thesaurus. 2nd ed.
Portland, OR: Timber Press. 816 p. [23878]
54. Kearney, Thomas H.; Peebles, Robert H.; Howell, John Thomas; McClintock,
Elizabeth. 1960. Arizona flora. 2d ed. Berkeley, CA: University of
California Press. 1085 p. [6563]
55. Keep, Elizabeth. 1968. Incompatibility in Rubus with special reference
to R. idaeus L. Canadian Journal of Genetic Cytology. 10: 253-262.
[6654]
56. Komarkova, Vera. 1986. Habitat types on selected parts of the Gunnison
and Uncompahgre National Forests. Final Report Contract No. 28-K2-234.
Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky
Mountain Forest and Range Experiment Station. 270 p. [1369]
57. Kooiman, Marianne; Linhart, Yan B. 1986. Structure and change in
herbaceous communities of four ecosystems in the Front Range, Colorado,
U.S.A. Arctic and Alpine Research. 18(1): 97-110. [4076]
58. Kramp, Betty A.; Patton, David R.; Brady, Ward W. 1983. The effects of
fire on wildlife habitat and species. RUN WILD: Wildlife/ habitat
relationships. Albuerque, NM: U.S. Department of Agriculture, Forest
Service, Southwestern Region, Wildlife Unit Technical Report. 29 p.
[152]
59. Krefting, Laurits W.; Roe, Eugene I. 1949. The role of some birds and
mammals in seed germination. Ecological Monographs. 19(3): 269-286.
[8847]
60. Kuchler, A. W. 1964. Manual to accompany the map of potential vegetation
of the conterminous United States. Special Publication No. 36. New York:
American Geographical Society. 77 p. [1384]
61. Larsen, J. A. 1929. Fires and forest succession in the Bitterroot
Mountains of northern Idaho. Ecology. 10: 67-76. [6990]
62. Lautenschlager, R. A. 1987. Interactions among red rapsberries and
northern forest trees. Proceedings of the Annual Meetings Northeastern
Weed Science Soc. 41: 186-187. [7007]
63. Lawrence, F. J. 1980. The current status of red raspberry cultivars in
the United States and Canada. Fruit Varieties Journal. 34(4): 84-89.
[6984]
64. Lawson, H. M.; Waister, P. D. 1972. The effects of soil cultivation
techniques on the growth and yield of the raspberry crop. Weed Research.
12: 96-106. [6604]
65. Lawson, H. M.; Waister, P. D. 1972. The response to nitrogen of a
raspberry plantation under contrasting systems of management for weed
and sucker control. Horticultural Research. 12: 43-55. [6653]
66. Lepofsky, Dana; Turner, Nancy J.; Kuhnlein, Harriet V. 1985. Determining
the availability of traditional wild plant foods: an example of Nuxalk
foods, Bella Coola, British Columbia. Ecology of Food and Nutrition. 16:
223-241. [7002]
67. Lockshin, L. S.; Elfving, D. C. 1981. Flowering response of 'Heritage'
red raspberry to temperature and nitrogen. HortScience. 16(4): 527-528.
[6824]
68. Longley, Albert E. 1924. Cytological studies in the genus Rubus.
American Journal of Botany. 11: 249-282. [6249]
69. Luby, J. J.; Hoover, E. E.; Bedford, D. S.; Munson, S. T.; Gray, W. H.;
Wildung, D. K.; Stushnoff, C. 1987. 'Redwing' raspberry. HortScience.
22(4): 681-682. [2957]
70. Lyon, L. Jack; Stickney, Peter F. 1976. Early vegetal succession
following large northern Rocky Mountain wildfires. In: Proceedings, Tall
Timbers fire ecology conference and Intermountain Fire Research Council
fire and land management symposium; 1974 October 8-10; Missoula, MT. No.
14. Tallahassee, FL: Tall Timbers Research Station: 355-373. [1496]
71. Mackenzie, K. A. D. 1979. The structure of the fruit of the red
raspberry (Rubus idaeus L.) in relation to abscission. Annals of Botany.
43(3): 355-362. [6822]
72. MacKerron, D. K. L. 1982. Growth and water use in the red raspberry
(Rubus idaeus L.) I. Growth and yield under differenct levels of soil
moisture stress. Journal of Horticultural Science. 57(3): 295-306.
[6979]
73. Marks, P. L. 1974. The role of pin cherry (Prunus pensylvanica L.) in
the maintenance of stability in northern hardwood ecosystems. Ecological
Monographs. 44: 73-88. [4144]
74. McRae, D. J. 1979. Prescribed burning in jack pine logging slash: a
review. Report 0-X-289. Sault Ste. Marie, ON: Canadian Forestry Service,
Great Lakes Forest Research Centre. 57 p. [7290]
75. Munz, Philip A. 1973. A California flora and supplement. Berkeley, CA:
University of California Press. 1905 p. [6155]
76. Ohmann, Lewis F.; Ream, Robert R. 1971. Wilderness ecology: virgin plant
communities of the Boundary Waters Canoe Area. Res. Pap. NC-63. St.
Paul, MN: U.S. Department of Agriculture, Forest Service, North Central
Forest Experiment Station. 55 p. [9271]
77. Ohmann, Lewis F.; Cushwa, Charles T.; Lake, Roger E.; [and others].
1973. Wilderness ecology: the upland plant communities, woody browse
production, and small mammals of two adj. 33-year-old wildfire areas in
northeastern Minnesota. Gen. Tech. Rep. NC-7. St. Paul, MN: U.S.
Department of Agriculture, Forest Service, North Central Forest
Experiment Station. 30 p. [6862]
78. Parminter, John. 1983. Fire history and fire ecology in the Prince
Rupert Forest region. In: Trowbridge, R. L.; Macadam, A., eds.
Prescribed fire--forest soils: Symposium proceedings; 1982 March 2-3;
Smithers, BC. Land Management Report Number 16. Victoria, BC: Province
of British Columbia, Ministry of Forests: 1-35. [8849]
79. Peek, J. M. 1974. A review of moose food habits studies in North
America. Le Naturaliste Canadien. 101: 195-215. [7420]
bsp; the vascular flora of the Carolinas. Chapel Hill, NC: The University of
North Carolina Press. 1183 p. [7606]
81. Raunkiaer, C. 1934. The life forms of plants and statistical plant
geography. Oxford: Clarendon Press. 632 p. [2843]
82. Roy, D. N.; Konar, S. K.; Banerjee, S.; [and others]. 1989. Uptake and
persistence of the herbicide glyphosate (Vison) in fruit of wild
blueberry and red raspberry. Canadian Journal of Forestry. 19: 842-847.
[8907]
83. Sanford, J. C.; Ourecky, D. K.; Reich, J. E. 1985. 'Titan' red
raspberry. HortScience. 20(6): 1133-1134. [6832]
84. Seymour, Frank Conkling. 1982. The flora of New England. 2d ed.
Phytologia Memoirs 5. Plainfield, NJ: Harold N. Moldenke and Alma L.
Moldenke. 611 p. [7604]
85. Siccama, T. G. 1974. Vegetation, soil, and climate on the Green
Mountains of Vermont. Ecological Monographs. 44: 325-249. [6859]
86. Stephens, H. A. 1973. Woody plants of the North Central Plains.
Lawrence, KS: The University Press of Kansas. 530 p. [3804]
87. Sundance Forest Fire, northern Idaho. Gen. Tech. Rep. INT-197. Ogden,
UT: U.S. Department of Agriculture, Forest Service, Intermountain
Research Station. 26 p. [2255]
88. Stiles, Edmund W. 1980. Patterns of fruit presentation and seed
dispersal in bird-disseminated woody plants in the Eastern deciduous
forest. American Naturalist. 116(5): 670-688. [6508]
89. Torre, Louis C.; Barritt, Bruce H. 1979. Red raspberry establishment
from root cuttings. Journal of the American Society for Horticultural
Science. 104(1): 28-31. [7028]
90. U.S. Department of Agriculture, Natural Resources Conservation Service. 2018.
PLANTS Database, [Online]. U.S. Department of Agriculture, Natural Resources
Conservation Service (Producer). Available: https://plants.usda.gov/.
[34262]
91. Van Dersal, William R. 1938. Native woody plants of the United States,
their erosion-control and wildlife values. Washington, DC: U.S.
Department of Agriculture. 362 p. [4240]
92. Temperature and development of red raspberry flower buds. Journal of the
American Horticultural Society. 104(1): 61-62. [6976]
93. Viereck, Leslie A.; Little, Elbert L., Jr. 1972. Alaska trees and
shrubs. Agric. Handb. 410. Washington, DC: U.S. Department of
Agriculture, Forest Service. 265 p. [6884]
94. Viereck, Leslie A.; Schandelmeier, Linda A. 1980. Effects of fire in
Alaska and adjacent Canada--a literature review. BLM-Alaska Tech. Rep.
6. Anchorage, AK: U.S. Department of the Interior, Bureau of Land
Mangement, Alaska State Office. 124 p. [7075]
95. Watson, L. E.; Parker, R. W.; Polster, D. F. 1980. Manual of plant
species suitablity for reclamation in Alberta. Vol. 2. Forbs, shrubs and
trees. Edmonton, AB: Land Conservation and Reclamation Council. 537 p.
[8855]
96. Weber, M. G. 1987. Decomposition, litter fall, and forest floor nutrient
dynamics in relation to fire in eastern Ontario jack pine ecosystems.
Canadian Journal of Forest Research. 17: 1496-1506. [7240]
97. Welander, Margareta. 1985. In vitro culture of raspberry (Rubus idaeus)
for mass propagation. Journal of Horticultural Science. 60(4): 493-499.
[6981]
98. Welsh, Stanley L.; Atwood, N. Duane; Goodrich, Sherel; Higgins, Larry
C., eds. 1987. A Utah flora. Great Basin Naturalist Memoir No. 9. Provo,
UT: Brigham Young University. 894 p. [2944]
99. Whitney, Gordon G. 1982. The productivity and carbohydrate economy of a
developing stand of Rubus idaeus. Canadian Journal of Botany. 60:
2697-2703. [6534]
100. Whitney, Gordon G. 1986. A demographic analysis of Rubus idaeus and
Rubus pubescens. Canadian Journal of Botany. 64: 2916-2921. [120]
101. Williams, I. H. 1959. Effects of environment on Rubus idaeus L. II.
Field observations on the variety Malling Promise. Journal of
Horticultural Science. 34: 170-175. [6606]
102. Williamson, B.; Lawson, H. M.; Woodford, J. A. T.; Hargreaves, A. J. ;
[and others].. 1979. Vigour control, an integrated approach to cane,
pest and disease management in red raspberry (Rubus idaeus). Annal of
Applied Biology. 92: 359-368. [6536]
103. Wright, Henry A. 1973. Fire as a tool to manage tobosa grasslands. In:
Proceedings--annual tall timbers fire ecology conference; 1972 June 8-9;
Lubbock, TX. Number 12. Tallahassee, FL: Tall Timbers Research Station:
153-167. [2612]
104. Yarnell, S. H. 1936. Chromosome behavior in blackberry-raspberry
hybrids. Journal of Agricultural Research. 52(5): 385-396. [6201]
105. Zager, Peter Edward. 1980. The influence of logging and wildfire on
grizzly bear habitat in northwestern Montana. Missoula, MT: University
of Montana. 131 p. Dissertation. [5032]
106. Zasada, John; Tappeiner, John; Maxwell, Bruce. 1989. Manual treatment of
Salmonberry or which bud's for you?. Cope Report, Coastal Oregon
Productivity Enhancement Program. 2(2): 7-9. [7060]
107. Stickney, Peter F. 1989. Seral origin of species originating in northern
Rocky Mountain forests. Unpublished draft on file at: U.S. Department of
Agriculture, Forest Service, Intermountain Research Station, Fire
Sciences Laboratory, Missoula, MT; RWU 4403 files. 7 p. [20090]
108. U.S. Department of the Interior, National Biological Survey. [n.d.]. NP
Flora [Data base]. Davis, CA: U.S. Department of the Interior, National
Biological Survey. [23119]
109. St. John, Harold. 1973. List and summary of the flowering plants in the
Hawaiian islands. Hong Kong: Cathay Press Limited. 519 p. [25354]
FEIS Home Page