Index of Species Information
SPECIES: Kalmia angustifolia
|
|
 |
| Sheep laurel in bloom. Wikimedia Commons image by Jomegat. |
Introductory
SPECIES: Kalmia angustifolia
AUTHORSHIP AND CITATION:
Van Deelen, Timothy R. 1991. Kalmia angustifolia. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station,
Fire Sciences Laboratory (Producer). Available:
https://www.fs.usda.gov/database/feis/plants/shrub/kalang/all.html [].
Revisions:
Images were added on 17 July 2018.
ABBREVIATION:
KALANG
SYNONYMS:
Kalmia angustifolia forma candida Fernald [43]
NRCS PLANT CODE:
KAAN
COMMON NAMES:
sheep laurel
lambkill
sheepkill
calfkill
dwarf-laurel
wicky
TAXONOMY:
The scientific name of sheep laurel is Kalmia angustifolia L. (Ericaceae).
Sheep laurel does not hybridize with other North American Kalmia species [43].
LIFE FORM:
Shrub
FEDERAL LEGAL STATUS:
No special status
OTHER STATUS:
NO-ENTRY
DISTRIBUTION AND OCCURRENCE
SPECIES: Kalmia angustifolia
GENERAL DISTRIBUTION:
Sheep laurel is found in northeastern North America from Newfoundland
and Labrador west through Ontario, south through Michigan, and
occasionally as far south as Georgia. It is most common in the eastern
Great Lakes region, the St. Lawrence River region, northern New England,
and the Maritime Provinces. Sheep laurel is occasional in the
Appalachian Mountains, on the Piedmont Plateau, and on the United
States' upper Atlantic Coastal Plain [36,41].
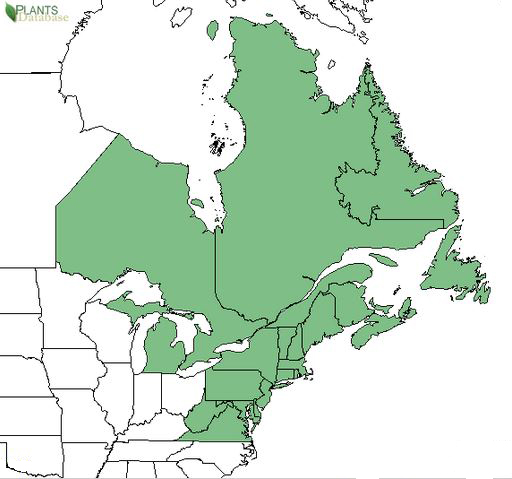 |
| Distribution of sheep laurel. Map courtesy of USDA, NRCS. 2018. The PLANTS Database.
National Plant Data Team, Greensboro, NC [2018, July 17] [40]. |
ECOSYSTEMS:
FRES10 White - red - jack pine
FRES11 Spruce - fir
FRES13 Loblolly - shortleaf pine
FRES19 Aspen - birch
STATES:
CT GA ME MD MA MI MN NH NJ NY
PA RI VT WI LB NB NF NS ON PE
PQ
BLM PHYSIOGRAPHIC REGIONS:
NO-ENTRY
KUCHLER PLANT ASSOCIATIONS:
K093 Great Lakes spruce - fir forest
K094 Conifer bog
K095 Great Lakes pine forest
K096 Northeastern spruce - fir forest
K106 Northern hardwoods
K107 Northern hardwoods - fir forest
K108 Northern hardwoods - spruce forest
K110 Northeastern oak - pine forest (Quercus-Pinus)
SAF COVER TYPES:
1 Jack pine
5 Balsam fir
12 Black spruce
13 Black spruce - tamarack
18 Paper birch
21 Eastern white pine
32 Red spruce
33 Red spruce - balsam fir
35 Paper birch - red spruce - balsam fir
37 Northern white cedar
45 Pitch pine
107 White spruce
SRM (RANGELAND) COVER TYPES:
NO-ENTRY
HABITAT TYPES AND PLANT COMMUNITIES:
Sheep laurel is a common understory shrub in eastern lowland forests.
It is characteristically found in coniferous, mixed, and hardwood stands
in eastern Canada and the northeastern United States [28,38]. On
peatlands, it often occurs in extensive, nearly pure stands known as
"heaths" [6]. Sheep laurel is a common dominant of bog communities in
the lower St. Lawrence lowlands [10] and grows in the New Jersey Pine
Barrens [11].
Common overstory associates include red spruce (Picea rubens) [2], black
spruce (Picea mariana) [8,42], jack pine (Pinus banksiana) [28], quaking
aspen (Populus tremuloides), and paper birch (Betula papyrifera) [37].
Understory associates include bog Labrador tea (Ledum groenlandicum),
low sweet blueberry (Vaccinium angustifolium), wintergreen (Gaultheria
procumbens), sweet fern (Comptonia peregrina) [43], Sphagnum spp., and
Cladonia spp. [8,37]
Published classification schemes listing sheep laurel as a dominant or
codominant member of a plant associations or community types include:
Geographical changes in the vegetation of raised bogs in the bay of
Fundy region of Maine and New Brunswick [7]
The principal plant associations of the St. Lawrence Valley [9].
MANAGEMENT CONSIDERATIONS
SPECIES: Kalmia angustifolia
IMPORTANCE TO LIVESTOCK AND WILDLIFE:
Sheep laurel is poisonous to livestock; hence the common names
sheepkill, lambkill, and calfkill [24,36,41]. Toxicity levels, defined
as the percent body weight of foliage needed to induce toxic symptoms,
are 0.15 percent for sheep, 0.20 percent for cattle, and 0.25 percent
for goats [24]. Sheep laurel also poisons horses. Poisoning
typically occurs during the winter when persistent sheep laurel leaves
are the only available vegetation above light snow cover. Poisoning
symptoms include salivation, watery eyes, running nose, vomiting with
convulsions, and paralysis [43].
Sheep laurel is not eaten by moose in Newfoundland [39] although it is
eaten by grouse [41], and is important winter food for caribou in
Ontario [1].
PALATABILITY:
Sheep laurel leaves are tough. Animals that eat them do so only when
other forage is unavailable [43].
NUTRITIONAL VALUE:
NO-ENTRY
COVER VALUE:
Sheep laurel provides nesting sites for willow ptarmigans [43].
VALUE FOR REHABILITATION OF DISTURBED SITES:
Although used to reclaim mined peatlands [13], sheep laurel is little
used for reclamation of other sites, so its value on other sites remains
unknown. It responds to transplanting by sprouting [17]. It returns
very little leaf litter to the soil. Sheep laurel builds up the raw
humus layer through root die-off which may include 20 percent of the
root mass greater than 0.33 inch (1 cm) in diameter and account for 9.2
tons of additional humus per acre (907 kg/ha) annually. It returns 19
pounds of nitrogen per acre (28 kg/ha) yearly [6].
Sheep laurel has a class IV gypsy-moth susceptibility: unfavored or
rarely fed upon [20].
OTHER USES AND VALUES:
NO-ENTRY
OTHER MANAGEMENT CONSIDERATIONS:
Sheep laurel is a serious pest in blueberry fields. Control by fire is
usually ineffective [34]. Sheep laurel has stronger rhizomes and sprout
growth than blueberry and requires repeated control [21]. Ten percent
of all the low sweet blueberry acreage in the Maritime Provinces
requires sheep laurel herbicide control treatment each year [43].
In addition to outcompeting conifer seedlings for nutrients, light, and
space [29], sheep laurel has an allelopathic effect on conifer seedlings
[5,29,39]. The establishment of sheep laurel-dominated heaths after
disturbance may produce soil conditions that prevent conifer seedling
establishment [5,6].
Several herbicide treatments have been tested for control of
sheep laurel. Most were difficult to use and ineffective [43].
BOTANICAL AND ECOLOGICAL CHARACTERISTICS
SPECIES: Kalmia angustifolia
GENERAL BOTANICAL CHARACTERISTICS:
Sheep laurel is a small, branchy, evergreen shrub which reaches 1 to 3
feet (30-90 cm) in height. It has pink, showy, five-part flowers on
stalked clusters. The fruit is a small capsule containing many seeds.
Capsules may persist on the branches for several years [4,36].
Sheep laurel forms a well-developed and closely interlacing network of
rhizomes. The root system consists of a taproot which may extend to a
depth of 3 feet (1 m) [43]. It also has fine roots nearer to the soil
surface [6]. Fine root depths vary from 3.5 inches (9 cm) in mineral
soil to 12 inches (31 cm) in bogs [14,39].
RAUNKIAER LIFE FORM:
Phanerophyte
REGENERATION PROCESSES:
Reproduction is primarily vegetative. Sheep laurel plants are clonal
and expand laterally. Sprouts grow from dormant buds on rhizomes. Very
little is known about sexual reproduction and seedling establishment.
Individual stems produce an estimated 7,100 seeds per year. Clones are
self-compatible, but seedlings from selfing are less vigorous than
seedlings from crossing [43].
SITE CHARACTERISTICS:
Sheep laurel grows on a variety of sites ranging from wet sphagnum bogs
to dry jack pine forests. It is frequently found on sites that are very
dry during the summer but saturated or flooded during the spring [34].
On peatland, it grows under both oligotrophic and ombrotrophic
conditions [3,8]. In the Northeast, sheep laurel grows in moist conifer
woods, pastures, barrens, roadsides, and open thickets [34].
Sheep laurel typically grows on podzols which have developed under a
cold to temperate climate on acidic parent material [43]. On peatland
it grows on well-drained peat and peaty podzols. On forested sites
sheep laurel grows on dry, acid, run-out soils; rocky, gravelly soils;
sandy loams; and iron-rich, lithosolic, or Ortstein podzols [7,34,36].
Its western range may be limited by high soil pH and lack of moisture
[43].
On peatlands in Newfoundland, sheep laurel grows independent of the
mineral soil, taking all of its nutrients from the organic layer. On
forested sites it roots in mineral soil [6].
Sheep laurel grows under a climate regime that is cold and wet for much
of the year. Minimum temperatures at the northern limits of its range
are -40 degrees Fahrenheit (-40 deg C) [43].
SUCCESSIONAL STATUS:
Sheep laurel is somewhat shade intolerant. Under low light conditions
it persists but does not grow appreciably. Overstory removal and
increased light trigger release, causing sheep laurel cover to quickly
increase [3]. It is considered early successional in the bog formation
sequence, although it may become dominant and arrest succession
following bog disturbance by fire or drainage [6]. On frequently
disturbed forest sites, most logging practices favor sheep laurel
establishment. Clones may expand and persist for several decades,
reducing tree establishment and creating heaths or "Kalmia barrens"
[5,6,32]. On undisturbed sites sheep laurel is replaced by trees such
as balsam fir and black spruce [5].
SEASONAL DEVELOPMENT:
Sheep laurel flowers during June and early July. Its fruit ripens
between late July and mid-September [23,36,43]. The seeds disperse in
early October. New shoot growth begins during late May and early June
[43].
FIRE ECOLOGY
SPECIES: Kalmia angustifolia
FIRE ECOLOGY OR ADAPTATIONS:
Sheep laurel sprouts soon after fire [35]. Persistent rhizomes in the
soil are protected from all but the most severe fires and allow it to
regenerate quickly [15,35].
FIRE REGIMES:
Find fire regime information for the plant communities in which this
species may occur by entering the species name in the FEIS home page under
"Find Fire Regimes".
POSTFIRE REGENERATION STRATEGY:
Small shrub, adventitious-bud root crown
FIRE EFFECTS
SPECIES: Kalmia angustifolia
IMMEDIATE FIRE EFFECT ON PLANT:
Fire kills aerial portions of sheep laurel stems. Light fires that do
not harm the buried rhizomes do not kill the plant. Severe fires
that consume the organic layer or sufficiently heat the soil surrounding
the rhizomes do kill sheep laurel [30].
DISCUSSION AND QUALIFICATION OF FIRE EFFECT:
Sheep laurel's ability to survive a fire depends on the survival of its
rhizomes. Sheep laurel rhizomes growing in forest mineral soils are
relatively shallowly buried and depend on the insulating value of the
soil and soil moisture for protection against lethal soil temperatures
[16]. On bogs or peatlands, sheep laurel rhizomes are generally deeper.
When wet, these soils offer both insulation and protection from fire.
When dry, these organic soils themselves may be consumed, killing the
sheep laurel rhizomes [15]. On peatlands, sheep laurel survives only
where the humus layer is not destroyed [30].
PLANT RESPONSE TO FIRE:
The immediate postfire response of sheep laurel is a decrease in
frequency and abundance [18]. However, it quickly responds to fire (and
cutting) with vigorous sprouting [29]. Fire stimulates the growth of
adventitious roots from the burned root stubs and rhizomes near the soil
surface [29,35]. Sprouts can be seen soon afterward. In an Ontario
study, new shoots were 2 inches (5 cm) tall 2 weeks after a fire, and 6
inches tall (15 cm) 6 weeks after the fire [35].
DISCUSSION AND QUALIFICATION OF PLANT RESPONSE:
Although visible on a site within 2 weeks of a fire, sheep laurel shows
the greatest increase in frequency between postfire year 1 and 2. Its
frequency may increase 500 percent and then remain relatively constant
for the next 40 years [31].
Regrowth after summer fires is slower than regrowth after spring or fall
fires, which shows a strong increase in sprout density. Summer fires
coincide with a period of low photosynthate reserves that follows the
spring growth spurt [14,17]. Light spring or fall fires encourage
prolific growth [31].
In a greenhouse experiment, Mallik [29] compared sheep laurel shrubs
that had been cut or cut and burned with control plants. There was
no significant difference between the number and density of new sprouts
for the treatments and for the control. Treatment sprouts were more
robust, but treatment rhizomes were smaller. Sheep laurel directs
photosynthates to aboveground growth at the expense of belowground
growth following disturbance.
FIRE MANAGEMENT CONSIDERATIONS:
All but the most severe fires enhance the growth of sheep laurel stands.
Permanent control of sheep laurel require fires severe enough to kill
the rhizomes; such fires often consume the organic layer of the soil
[31]. Managers should note that most sheep laurel-dominated communities
in Nova Scotia are associated with frequent fires. Logging and fire
promote heath formation [5]. The high stem density in heaths causes
severe fires when they eventually burn. Frequent fires reduce fuel
accumulation and, consequently, are less severe [26].
REFERENCES
SPECIES: Kalmia angustifolia
REFERENCES:
1. Ahti, T.; Hepburn, T. L. 1967. Preliminary studies on woodland caribou
range, especially on lichen stands, in Ontario. Res. Rep. (Wildlife) No.
74. Toronto, ON: Ontario Department of Lands and Forests, Research
Branch. 134 p. [13294]
2. Blum, Barton M. 1990. Picea rubens Sarg. red spruce. In: Burns, Russell
M.; Honkala, Barbara H., technical coordinators. Silvics of North
America. Volume 1. Conifers. Agric. Handb. 654. Washington, DC: U.S.
Department of Agriculture, Forest Service: 250-259. [13388]
3. Brumelis, G.; Carleton, T. J. 1989. The vegetation of post-logged black
spruce lowlands in central Canada. II. Understory vegetation. Journal of
Applied Ecology. 26: 321-339. [7864]
4. Chapman, William K.; Bessette, Alan E. 1990. Trees and shrubs of the
Adirondacks. Utica, NY: North Country Books, Inc. 131 p. [12766]
5. Damman, A. W. H. 1964. Some forest types of central Newfoundland and
their relation to environmental factors. Forest Science Monograph 8.
Washington, DC: Society of American Foresters. 62 p. [14281]
6. Damman, A. W. H. 1971. Effect of vegetation changes on the fertility of
a Newfoundland forest site. Ecological Monographs. 41(3): 253-270.
[9751]
7. Damman, A. W. H. 1977. Geographical changes in the vegetation pattern of
raised bogs in the Bay of Fundy region of Maine and New Brunswick.
Vegetatio. 35(3): 137-151. [10158]
8. Damman, Antoni W. H.; French, Thomas W. 1987. The ecology of peat bogs
of the glaciated northeastern United States: a community profile.
Biological Report 85(7.16). Washington, DC: U.S. Department of the
Interior, Fish and Wildlife Service, Research and Development, National
Wetlands Research Center. 100 p. [9238]
9. Dansereau, Pierre. 1959. The principal plant associations of the Saint
Lawrence Valley. No. 75. Montreal, Canada: Contrib. Inst. Bot. Univ.
Montreal. 147 p. [8925]
10. Dansereau, Pierre; Segadas-Vianna, Fernando. 1952. Ecological study of
the peat bogs of eastern North America. Canadian Journal of Botany.
30(5): 490-520. [8869]
11. Ehrenfeld, Joan G. 1986. Wetlands of the New Jersey Pine Barrens: the
role of species composition in community function. American Midland
Naturalist. 115(2): 301-313. [8650]
12. Eyre, F. H., ed. 1980. Forest cover types of the United States and
Canada. Washington, DC: Society of American Foresters. 148 p. [905]
13. Famous, Norman C.; Spencer, M. 1989. Revegetation patterns in mined
peatlands in central and eastern North America studied. Restoration and
Management Notes. 7(2): 95-96. [10171]
14. Flinn, Marguerite Adele. 1980. Heat penetration and early postfire
regeneration of some understory species in the Acadian forest. Halifax,
NB: University of New Brunswick. 87 p. Thesis. [9876]
15. Flinn, Marguerite A.; Pringle, Joan K. 1983. Heat tolerance of rhizomes
of several understory species. Canadian Journal of Botany. 61: 452-457.
[8444]
16. Flinn, Marguerite A.; Wein, Ross W. 1977. Depth of underground plant
organs and theoretical survival during fire. Canadian Journal of Botany.
55: 2550-2554. [6362]
17. Flinn, Marguerite A.; Wein, Ross W. 1988. Regrowth of forest understory
species following seasonal burning. Canadian Journal of Botany. 66:
150-155. [3014]
18. Foster, David R. 1985. Vegetation development following fire in Picea
mariana (black spruce) - Pleurozium forests of south-eastern Labrador,
Canada. Journal of Ecology. 73: 517-534. [7222]
19. Garrison, George A.; Bjugstad, Ardell J.; Duncan, Don A.; [and others].
1977. Vegetation and environmental features of forest and range
ecosystems. Agric. Handb. 475. Washington, DC: U.S. Department of
Agriculture, Forest Service. 68 p. [998]
20. Gottschalk, Kurt W. 1988. Gypsy moth and regenerating Appalachian
hardwood stands. In: Smith, H. Clay; Perkey, Arlyn W.; Kidd, William E.,
Jr., eds. Guidelines for regenerating Appalachian hardwood stands:
Workshop proceedings; 1988 May 24-26; Morgantown, WV. SAF Publ. 88-03.
Morgantown, WV: West Virginia University Books: 241-254. [13950]
21. Hall, I. V. 1959. Plant populations in blueberry stands developed from
abandoned hayfields and woodlots. Ecology. 40(4): 742-743. [9108]
22. Kartesz, John T.; Kartesz, Rosemarie. 1980. A synonymized checklist of
the vascular flora of the United States, Canada, and Greenland. Volume
II: The biota of North America. Chapel Hill, NC: The University of North
Carolina Press; in confederation with Anne H. Lindsey and C. Richie
Bell, North Carolina Botanical Garden. 500 p. [6954]
23. Keppie, Daniel M.; Towers, Julie. 1990. Using phenology to predict
commencement of nesting of female spruce grouse (Dendragapus
canadensis). American Midland Naturalist. 124(1): 164-170. [12590]
24. Kingsbury, John M. 1964. Poisonous plants of the United States and
Canada. Englewood Cliffs, NJ: Prentice-Hall, Inc. 626 p. [122]
25. Kuchler, A. W. 1964. Manual to accompany the map of potential vegetation
of the conterminous United States. Special Publication No. 36. New York:
American Geographical Society. 77 p. [1384]
26. Little, S. 1964. Fire ecology and forest management in the New Jersey
pine region. In: Proceedings, 3rd annual Tall Timbers fire ecology
conference; 1964 April 9-10; Tallahassee, FL. No. 3. Tallahassee, FL:
Tall Timbers Research Station: 35-59. [5893]
27. Lyon, L. Jack; Stickney, Peter F. 1976. Early vegetal succession
following large northern Rocky Mountain wildfires. In: Proceedings, Tall
Timbers fire ecology conference and Intermountain Fire Research Council
fire and land management symposium; 1974 October 8-10; Missoula, MT. No.
14. Tallahassee, FL: Tall Timbers Research Station: 355-373. [1496]
28. MacLean, David A.; Wein, Ross W. 1977. Changes in understory vegetation
with increasing stand age in New Brunswick forests: species composition,
cover, biomass, and nutrients. Canadian Journal of Botany. 55:
2818-2831. [10106]
29. Mallik, A. U. 1991. Cutting, burning, and mulching to control Kalmia:
results of a greenhouse experiment. Canadian Journal of Forest Research.
21: 417-420. [14426]
30. Martin, J. Lynton. 1955. Observations on the origin and early
development of a plant community following a forest fire. Forestry
Chronicle. 31: 154-161. [11363]
31. Martin, J. Lynton. 1956. An ecological survey of burned-over forest land
in southwestern Nova Scotia. Forestry Chronicle. 32: 313-336. [8932]
32. Niering, William A.; Goodwin, Richard H. 1974. Creation of relatively
stable shrublands with herbicides: arresting "succession" on
rights-of-way and pastureland. Ecology. 55: 784-795. [8744]
33. Raunkiaer, C. 1934. The life forms of plants and statistical plant
geography. Oxford: Clarendon Press. 632 p. [2843]
34. Roland, A. E.; Smith, E. C. 1969. The flora of Nova Scotia. Halifax, NS:
Nova Scotia Museum. 746 p. [13158]
35. Smith, David William. 1966. Studies in the taxonomy and ecology of
blueberries (Vaccinium, subgenus Cyanococcus) in Ontario. Toronto, ON:
University of Toronto. 276 p. Dissertation. [10872]
36. Soper, James H.; Heimburger, Margaret L. 1982. Shrubs of Ontario. Life
Sciences Misc. Publ. Toronto, ON: Royal Ontario Museum. 495 p. [12907]
37. Strang, R. M. 1971. The ecology of the rocky heathlands of western Nova
Scotia. In: Proceedings, annual Tall Timbers fire ecology conference;
1970 August 20-21; Fredericton, NB. No. 10. Tallahassee, FL: Tall
Timbers Research Station: 287-292. [5466]
38. Telfer, E. S. 1972. Understory biomass in five forest types in
southwestern Nova Scotia. Canadian Journal of Botany. 50: 1263-1267.
[13933]
39. Thompson, I. D.; Mallik, A. U. 1989. Moose browsing and allelopathic
effects of Kalmia angustifolia on balsam fir regeneration in central
Newfoundland. Canadian Journal of Forest Research. 19(4): 524-526.
[13238]
40. U.S. Department of Agriculture, Natural Resources Conservation Service.
2018. PLANTS Database, [Online]. U.S. Department of Agriculture, Natural
Resources Conservation Service (Producer). Available: https://plants.usda.gov/.
[34262]
41. Van Dersal, William R. 1938. Native woody plants of the United States,
their erosion-control and wildlife values. Washington, DC: U.S.
Department of Agriculture. 362 p. [4240]
42. Viereck, Leslie A.; Johnston, William F. 1990. Picea mariana (Mill.)
B.S.P. black spruce. In: Burns, Russell M.; Honkala, Barbara H.,
technical coordinators. Silvics of North America. Volume 1. Conifers.
Agric. Handb. 654. Washington, DC: U.S. Department of Agriculture,
Forest Service: 227-237. [13386]
43. Hall, Ivan V.; Jackson, Lloyd P.; Everett, C. Fred. 1973. The biology of
Canadian weeds. 1. Kalmia angustifolia L. Canadian Journal of Plant
Science. 53: 865-873. [14592]
44. Stickney, Peter F. 1989. Seral origin of species originating in northern
Rocky Mountain forests. Unpublished draft on file at: U.S. Department of
Agriculture, Forest Service, Intermountain Research Station, Fire
Sciences Laboratory, Missoula, MT; RWU 4403 files. 7 p. [20090]
FEIS Home Page