Bromus hordeaceus
| |
 |
| Soft brome. Image by Robert Videki, Doronicum Kft., Bugwood.org. |
AUTHORSHIP AND CITATION:
Howard, Janet L. 1998. Bromus hordeaceus. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service,
Rocky Mountain Research Station, Fire Sciences Laboratory (Producer).
Available: https://www.fs.usda.gov/database/feis/plants/graminoid/brohor/all.html[].
Revisions:
On 10 September 2018, the common name of this species was changed in FEIS
from: soft chess
to: soft brome. Images were also added.
ABBREVIATION:
BROHOR
SYNONYMS:
Bromus mollis L. [22,38,50,59,83]
B. molliformis Godron [92]
B. arvensis L. [83]
B. racemosus L. [83,92]
= B. hordeaceus L. [35,49,56,93,95]
B. h. subsp. molliformis (Godron) Maire [49]
= Bromus hordeaceus L. subsp. divaricatus (Bonnier & Layens) Kerguélen [89]
B. h. subsp. molliformis (Lloyd) Maire & Weiller [56]
= Bromus hordeaceus L. subsp. divaricatus (Bonnier & Layens) Kerguélen [89]
NRCS PLANT CODE:
BRHO2
BRHOD
BRHOF
BRHOH
BRHOP
BRHOT
COMMON NAMES:
soft brome
common soft-brome
soft chess
TAXONOMY:
Most North American systematists recognize Bromus hordeaceus L. as the
scientific name of soft brome [35,49,56,89,93,95]. European systematists
generally describe this entity as B. mollis L. [51,64]. Subspecies of
soft brome occurring in North America are [56,89]:
Bromus hordeaceus L. subsp. divaricatus (Bonnier & Layens) Kerguélen
Bromus hordeaceus L. subsp. ferronii (Mabille) P.M. Sm.
Bromus hordeaceus L. subsp. hordeaceus
Bromus hordeaceus L. subsp. pseudothominii (P.M. Sm.) H. Scholz
Bromus hordeaceus L. subsp. thominei (Hardham ex Nyman) Braun-Blanq.
Soft brome hybridizes with erect chess (B. erectus) [3] and
occasionally with Japanese brome (B. japonicus) [95].
LIFE FORM:
Graminoid
FEDERAL LEGAL STATUS:
No special status
OTHER STATUS:
NO-ENTRY
DISTRIBUTION AND OCCURRENCE
SPECIES: Bromus hordeaceus
GENERAL DISTRIBUTION:
Soft brome is native to Eurasia, where it is most common in the
Mediterranean region [61,95]. It has naturalized in all other
continents except Antarctica [54,95]. Soft brome is widely distributed
but scattered and uncommon in most of North America [35,38,54,59],
ranging from coastal southern Alaska south to Baja California and east
to Maine, North Carolina, and Texas [22,35,38,50,54,83]. Soft brome is
most common in low-elevation valleys and foothills of California and
southwestern Oregon where climate is mediterranean. It is more abundant
in mediterranean areas of California than in Mediterranean Europe [61].
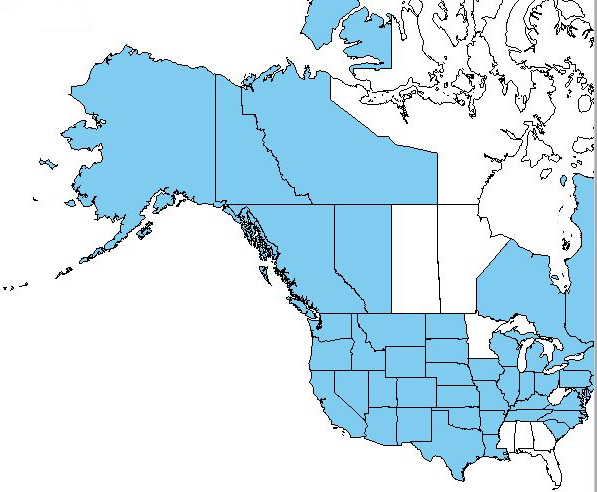 |
| Distribution of soft brome in Canada and the United States. Map courtesy of USDA, NRCS. 2018. The PLANTS Database.
National Plant Data Team, Greensboro, NC. [2018, September 10] [89]. |
ECOSYSTEMS:
FRES28 Western hardwoods
FRES29 Sagebrush
FRES30 Desert shrub
FRES34 Chaparral-mountain shrub
FRES38 Plains grasslands
FRES42 Annual grasslands
STATES:
AK AZ CA CO CT ID IL KS ME MA
MI MT NE NV NM ND NC OR RI SD
TX UT WA WI AB BC MB NB NF ON
PE PQ SK MEXICO
BLM PHYSIOGRAPHIC REGIONS:
1 Northern Pacific Border
2 Cascade Mountains
3 Southern Pacific Border
4 Sierra Mountains
5 Columbia Plateau
6 Upper Basin and Range
7 Lower Basin and Range
8 Northern Rocky Mountains
11 Southern Rocky Mountains
12 Colorado Plateau
13 Rocky Mountain Piedmont
14 Great Plains
15 Black Hills Uplift
16 Upper Missouri Basin and Broken Lands
KUCHLER PLANT ASSOCIATIONS:
K009 Pine-cypress forest
K026 Oregon oakwoods
K030 California oakwoods
K033 Chaparral
K034 Montane chaparral
K035 Coastal sagebrush
K038 Great Basin sagebrush
K040 Saltbush-greasewood
K048 California steppe
SAF COVER TYPES:
233 Oregon white oak
246 California black oak
248 Knobcone pine
249 Canyon live oak
250 Blue oak-foothills pine
255 California coast live oak
SRM (RANGELAND) COVER TYPES:
201 Blue oak woodland
202 Coast live oak woodland
203 Riparian woodland
204 North coastal shrub
205 Coastal sage shrub
206 Chamise chaparral
207 Scrub oak mixed chaparral
208 Ceanothus mixed chaparral
209 Montane shrubland
214 Coastal prairie
215 Valley grassland
414 Salt desert shrub
HABITAT TYPES AND PLANT COMMUNITIES:
Soft brome is typically dominant in annual grassland communities of
California and southwestern Oregon [9,61,63]. It is an important
component of some sagebrush (Artemisia spp.) steppe and Palouse prairie
communities of eastern Washington and Oregon and southern Idaho,
especially where cheatgrass (Bromus tectorum) is a community dominant
[24,82]. Soft brome is not usually important in other plant communities
in which it occurs [38,54,76,83].
California annual grassland - The native prairie that occurred on
coastal and inland valleys of California and southwestern Oregon has
been almost entirely replaced by annual grassland, agricultural land, or
urban areas [9,19,41,45]. Conversion from native prairie to annual
grassland occurred in less than two hundred years [45,61]. Soft brome
is thought to have naturalized in native California prairie in about
1860 [45]. Species composition of native grasslands was poorly
documented and will always be open to debate [96]. Most experts agree
that coastal prairie and mesic inland valleys were dominated by
perennial bunchgrasses [10,11,20,45,61]. Drier inland valleys may have
been dominated by native annual grasses [8]. Hoover [52] argued that
most native California prairie was dominated by annual forbs.
Species composition in California annual grassland is complex and
varied: Even slight differences in climate, topography, and soil type
can alter species composition [45,61]. However, soft brome dominates
California annual grassland communities more often than any other plant
species [45,46,47]. In Pinnacles National Monument, for example, soft
brome has 26 percent cover and 100 percent frequency: It is the most
commonly occurring seed plant in the Monument [41]. Even where it is
not dominant, soft brome is usually an important component of annual
grassland vegetation [45,61].
Publications describing plant communities in which soft brome is a
dominant part of the vegetation are listed below.
Plant communities of Santa Rosa Island, Channel Islands National Park [19]
Vegetation and floristics of Pinnacles National Monument [41]
Valley grassland [45]
Coastal prairie and northern coastal scrub [47]
Plant associations within the Interior Valleys of the Umpqua River
Basin, Oregon [85]
Plant species commonly associated with soft brome in California and
southwestern Oregon are listed below.
California: annual grassland - Broad-leaved filaree (Erodium botrys)
commonly codominates with soft brome throughout California annual
grassland. Red brome (B. rubens) and cutleaf filaree (E. cicutarium)
are also common associates, usually replacing soft brome and
broad-leaved filaree as dominants in portions of the Central Valley
where annual precipitation is less than 12 inches (305 mm) [9]. Other
common annuals include ripgut brome (B. rigidus), slender oat (Avena
barbata), wild oat (A. fatua), rattail fescue (Vulpia myuros), bur
clover (Medicago hispida), and yellow starthistle (Centaurea
solstitialis). Native perennial associates include purple needlegrass
(Stipa pulchra), Sandberg bluegrass (Poa secunda), melic grass (Melica
californica), California oatgrass (Danthonia californica), bottlebrush
squirreltail (Elymus elymoides), Spanish clover (Lotus americanus), and
ground lupine (Lupinus bicolor) [16,47].
California hardwoods: Soft brome is dominant to common in the
understory of oak (Quercus spp.) and other upland hardwood types.
Upland tree associates of soft brome not previously listed in SAF COVER
TYPES include valley oak (Q. lobata), tanoak (Lithocarpus densiflorus),
California black walnut (Juglans californica), and California buckeye
(Aesculus californica) [61,87,88]. Soft brome also occurs in Fremont
cottonwood/willow (Populus fremontii/Salix spp.) and other riparian
types [97].
Oregon: annual grassland - Soft brome/hedgehog dogtail (Cynosurus
schinatus) communities occur on grassy balds of the Umpqua River Basin.
Associated grasses include California oatgrass, pine bluegrass (P.
scabrella), Sandberg bluegrass, and bottlebrush squirreltail [85].
Oregon white oak - Associates of soft brome in Oregon white oak (Q.
garryana) communities of southwestern Oregon include California brome
(B. carinatus), sheep fescue (Festuca ovina), birchleaf
mountain-mahogany (Cercocarpus betuloides), California black oak (Q.
kelloggii), poison-oak (Toxicodendron diversilobum), blue wildrye (E.
glaucus), rough bluegrass (P. trivalis), and burr chervil (Anthriscus
caucalis) [78].
Basin big sagebrush (Artemisia tridentata ssp. tridentata) - At the
Sheep Rock Unit of John Day Fossil Beds National Monument, soft brome
associates in basin big sagebrush communities include Idaho fescue (F.
idahoensis), bluebunch wheatgrass (Pseudoroegneria spicata), cheatgrass,
western yarrow (Achillea millefolium), smallflower woodlandstar
(Lithaphragma parviflora), and western juniper (Juniperus occidentalis)
[82].
A medusahead (Taeniatherum caput-medusae)-rattail fescue-soft brome
community has been described in a bluebunch wheatgrass-Sandberg
bluegrass habitat type near Pendleton, Oregon [18].
MANAGEMENT CONSIDERATIONS
SPECIES: Bromus hordeaceus
IMPORTANCE TO LIVESTOCK AND WILDLIFE:
Soft brome is nutritious and palatable forage. Sampson and others [81]
rated it the best forage of all California's annual bromes. The awns
are short and soft, and livestock graze soft brome even after seeds
mature. Because soft brome matures later than most annual grasses and
the seeds do not readily shatter, cattle graze it well into summer,
gaining extra nutrition from the seeds [80,81].
Use of soft brome by native ungulates may be sparse in some areas. In
Point Reyes National Park, California, tule elk and mule deer avoided
soft brome and ripgut brome. Although grasses were the primary
component in the fall diets of tule elk, the elk used the annual bromes
very little. Grasses were less important in the diets of mule deer, but
annual bromes were the least preferred of the grass species that the
mule deer grazed [37].
PALATABILITY:
In Montana and Utah, palatability of soft brome has been rated fair for
wild and domestic ungulates, small mammals, small nongame birds, and
upland game birds. Palatability was rated poor for waterfowl in Utah
[26].
NUTRITIONAL VALUE:
Nutritional content of fresh, immature soft brome was as follows [73]:
Protein (N x 6.25, %) Potassium (%) 4.00
cattle 14.2 Ash (%) 12.2
domestic goats 14.5 Crude fiber (%) 24.2
horses 13.8 Calcium (%) 0.59
rabbits 13.5 Phosphorus (%) 0.39
domestic sheep 14.9
COVER VALUE:
In Utah, cover value of soft brome for small mammals, small nongame
birds, and upland game birds was rated fair. Cover value for waterfowl
was rated poor [26].
VALUE FOR REHABILITATION OF DISTURBED SITES:
NO-ENTRY
OTHER USES AND VALUES:
Soft brome is planted for hay. The seed is commercially available
[22,25].
OTHER MANAGEMENT CONSIDERATIONS:
Annual grasslands - Soft brome and other exotic annuals probably
replaced native California prairie because native perennial grasses were
severely overgrazed over several periods of extended drought [45,61,74].
Annual grasses are far more prolific seed producers than are perennials.
Once established, soft brome and other annual grasses probably
interfered greatly with perennial grass regrowth, seed production, and
seedling establishment [61]. In a greenhouse experiment, soft brome has
also been shown to interfere with seedling establishment of coyote bush
(Baccharis pilularis), a native chaparral shrub [23]. Soft brome and
other exotic annuals can probably not be eliminated from the California
flora [45,58,61]. Although some fire and grazing treatments have
reduced soft brome and other annuals, results have been mixed.
Control: grazing - Soft brome may be partially controlled by spring
grazing. Defoliation within a week after flowering has been found to be
effective in reducing seed formation in annual bromes [30]. Laude [67]
found that removing terminal buds of soft brome prevented leaf
elongation and seed production. Treatments of spring grazing and fall
fire have been successful in reducing soft brome (see FIRE MANAGEMENT
CONSIDERATIONS).
No grazing - Attempts to reduce soft brome cover by cessation of
grazing have sometimes succeeded. In the short term, cover of soft brome
and other annuals declined after cattle were removed from Golden Gate
National Recreation Area, California, in 1984. The next 3 years were
droughty, but native perennial cover increased relative to cover of
annuals despite low rainfall [88]. Cessation of grazing also reduced
soft brome in an upper riparian zone in eastern Oregon. On plots with
10 years of late summer cattle grazing, soft brome cover increased
greatly: Cover on grazed plots was 1.7 percent the first year of the
study and 47.5 percent in the tenth year. On exclosures, cover of soft
brome declined over the 10-year study period [39]. However, Heady [45]
found that in Mendocino County, California, soft brome and other annuals
continued to dominate the Hopland Field Station despite protection from
grazing for at least 40 years.
Fire - Studies using prescribed fire to control soft brome are discussed
in the FIRE EFFECTS section.
BOTANICAL AND ECOLOGICAL CHARACTERISTICS
SPECIES: Bromus hordeaceus
GENERAL BOTANICAL CHARACTERISTICS:
Soft brome is a cool-season exotic grass [38,49,50]. It is usually an
annual but is sometimes a biennial in the Great Basin and the Northeast
[35,95]. The erect to ascending plants are 4.4 to 26 inches (11-65 cm)
tall. Soft brome is generally pubescent, but culms and/or spikelets are
occasionally glabrous [50]. Awns are straight and from 0.16 to 0.4 inch
(4-10 mm) long [95].
RAUNKIAER LIFE FORM:
Therophyte
REGENERATION PROCESSES:
As an annual, soft brome regenerates entirely from seed. Soft brome is
mostly self-pollinating [55]. Seed set insufficient to maintain soft
brome populations has not been observed in the field [17,43]. Ewing and
Menke [27,28] found that drought reduced average mass and number of
seed, but some plants produced seed even under severe drought
conditions. Viable seeds germinate in their first autumn. Little seed
is carried over from year to year in the seedbank [27,28,98], although
dry-stored soft brome seed may remain viable for decades [53].
Germination is best on a seedbed of moderate mulch, but some seed
germinates without mulch [7,9]. In the laboratory, soft brome required
stratification to germinate [31,32], but not light [31]. Temperature
range for germination is wide, with best germination occurring between
50 and 86 degrees Fahrenheit (10-30 deg C) [4,31]. Seeds become dormant
with freezing temperatures or temperatures over 100 degrees Fahrenheit
(38 deg C) [31,55]. Most germinating seeds survive the sporadic cycles
of wetting and drying that are common in fall in mediterranean climates.
Flood [31] found that in the greenhouse, germination rates of soft brome
seed were actually better when seeds were exposed to several cycles of
wetting and drying.
Seedling establishment is limited by freezing temperature and exposure
to drying. At the Hopland Field Station, California, soft brome
coverage was best when germination was followed by warm autumn nights.
Ripgut brome became dominant in years when temperatures fell below
freezing in October and November [46]. Survival of soft brome seedlings
is enhanced by moisture-retaining clay substrates or mulches [46,47,57].
Kay [57] reported that seedling establishment of soft brome on
decomposed granite was 17 times greater when straw mulch was applied.
Soft brome seedlings grow rapidly. Rate of greenhouse-grown soft brome
seedlings was as follows [23]:
Age root length shoot length
(weeks) (cm) (cm)
_______ ___________ ____________
1 7.2 4.3
5 18.0 6.0
9 50.0 8.0
SITE CHARACTERISTICS:
Soft brome occurs mostly in waste places in Alaska, the Pacific
Northwest, the Great Plains, the Southwest, and the East [35,38,54,59].
Soils and aspect: Soft brome grows on a variety of soil types including
serpentine and caliche [19,72,80]. Best growth occurs on clay loam and
sandy soils [80]. In inland California, soft brome is most common on
deep, clayey soils [2] receiving 26 to 40 inches (650-1,000 mm) of
annual precipitation [12]. On the coast, it is most common on sandy
soils [47]. In Somewhere, California, McNaughton [72] found that soft
brome occurred on all aspects but was most common on southwest slopes.
Climate: Dry mediterranean climates are most favorable to soft brome.
Soft brome is probably more common in California than in its native
Mediterranean because the drier California climate favors establishment
of annual grasses over perennial herbs and shrubs. The relatively
moister climate of the Mediterranean favors perennials [61].
Outside mediterranean regions of California and southwestern Oregon,
soft brome is most common in the cold climates of the Pacific Northwest
[50] and in northern portions of the Great Basin [95]. It is uncommon
in warm desert regions [49,59]. Soft brome is probably not well adapted
to the climate of the Southeast: It does not occur further south than
North Carolina, where it is very rare [76].
Elevation: Soft brome occurs at the following elevations:
California below 6,300 feet (2,100 m) [49]
Colorado 5,000 to 9,200 feet (1,500-2,800 m) [42]
Utah 4,220 to 8,350 feet (1,280-2,530 m) [95]
SUCCESSIONAL STATUS:
Soft brome occurs on newly disturbed sites, in mid-succession, and on
sites left undisturbed for decades [24,45,61].
California annual grassland - In the absence of disturbance, soft brome
and other annual grasses tend to increase at the expense of forbs and
perennial grasses [40,45]. Heady [45] found soft brome was an important
component of California annual grassland that had not been burned or
grazed by livestock for at least 40 years. Mulch, which accumulates in
the absence of heavy grazing and/or fire, tends to favor germination of
soft brome and other annual bromes over forbs and perennial grasses
[45]. Heady and others [46] reported that soft brome decreased on
heavily grazed sites, probably because grazing removed mulch. Over 3
years, soft brome coverage increased greatly (from 0.9% to 37.3%) on a
newly disturbed site on the Hopland Field Station. However, soft brome
coverage remained below 2 percent on plots where mulch was mechanically
removed in each of the 3 years [45].
Chaparral - Soft brome and other annual grasses may be successional to
chaparral shrubs on some sites. Repeated burning, often intentional for
the purpose of "type-conversion" of chaparral to grassland, has
eliminated woody species on some sites. In the absence of heavy grazing
and/or fire, woody plants have recolonized some of these burned sites
[21,61,69]. Equilibrium dynamics of annual grassland and chaparral are
not well understood, however, and probably differ by site. On level
terrain with heavy clay soil, soft brome and other annual grasses are
apparently stable and do not succeed to woody shrubs [61]. Woody
species may displace annuals on nutrient-poor, rocky slopes [79].
Palouse prairie - In old-field succession on a bluebunch
wheatgrass-Sandberg bluegrass habitat type in eastern Washington, soft
brome was an important component of the vegetation on new fields, young
fields (1-12 years since cultivation), and old fields (39-52 years since
cultivation). Soft brome cover (percent) was as follows [24]:
Time since cultivation
___________________________________________________________
New field 1 year 12 years 39 years 52 years
_________ ______ ________ ________ ________
1.25 0.10 1.55 1.30 0.12
SEASONAL DEVELOPMENT:
Soft brome germinates and begins growth in fall [28,46]. Vegetative
growth slows or stops early in winter and resumes early the next growing
season [28]. Flowering occurs in early spring. Seeds mature later in
the season than do seeds of most annual grass species. In California,
soft brome seed matures in early summer. Seeds do not readily shatter
upon maturity and are shed about a month after ripening [46,80].
Phenological development of soft brome on the central coast of
California was as follows [46]:
1971 1972
__________ ___________
vegetative growth early Feb. early Feb.
boot stage mid-March early April
flowering begins mid-April mid-April
peak flowering late April mid-April
flowering ends late May mid-May
seeds ripen early June late May
plant dies late June late May
seeds disperse ---- early Aug.
Soft brome flowers from May to July in the Pacific Northwest and the
northern Great Basin [22,51].
FIRE ECOLOGY
SPECIES: Bromus hordeaceus
FIRE ECOLOGY OR ADAPTATIONS:
Fire autecology: Summer and fall fires have no direct effect on soft
brome. Soft brome has usually senesced and shattered seed when the fire
season starts. The seed is not killed until fire temperatures rise
above approximately 200 degrees Fahrenheit (93 deg C). Since grassland
fires are usually fast-burning and relatively "cool," soft brome seed is
usually not damaged by fire [71,80]. Fire can affect relative abundance
of soft brome in the postfire plant community, however [61,82]. Fire
removes mulch, which favors annual forbs over soft brome. Some soft
brome germinates the fall after fire, but best germination occurs in
mid-succession, when mulch layer is moderate [7,9].
Fire regimes: California native grassland - Data are lacking to
quantify intensity and frequency of fire in pristine California prairie.
It is generally accepted that lightning-caused fire was part of the
evolutionary history of California prairie. The California Division of
Forestry reported an average of 312 lightning-ignited fires per year in
its fire protection area, which is 43 percent woodland-annual grassland.
Frequency of lightning-caused fires was probably at least as great in
the presettlement era [45].
Native Americans may have used frequent fire to enhance production of
edible perennial bunchgrass seeds [13]. Fire enhances flowering and
seedling recruitment for some perennial bunchgrasses native to
California prairie including purple needlegrass [62] and bottlebrush
squirreltail [99]. Both species show mass flowering after fire and
require mineral soil for establishment [36,60].
Annual grassland - Since California annual grassland has existed for
less than two hundred years, it has no evolutionary history of fire.
Like the perennial grassland that preceded it, however, California
annual grassland is a fire-tolerant ecosystem [61]. Studies attempting
to promote native perennial bunchgrasses over exotic annuals by using
prescribed fire have had mixed results. These results are summarized
in FIRE MANAGEMENT CONSIDERATIONS.
FIRE REGIMES:
Find fire regime information for the plant communities in which this
species may occur by entering the species name in the FEIS home page under
"Find Fire Regimes".
POSTFIRE REGENERATION STRATEGY:
Ground residual colonizer (on-site, initial community)
FIRE EFFECTS
SPECIES: Bromus hordeaceus
IMMEDIATE FIRE EFFECT ON PLANT:
Fire has little direct effect on soft brome. Wildland and prescribed
fires usually occur after soft brome has dried and shattered seed
[44,45,46].
PLANT RESPONSE TO FIRE:
Fire may reduce soft brome in the short term [48]. Species composition
in the postfire plant community is difficult to predict, however.
Year-to-year plant composition in annual grassland is highly dependent
upon local weather patterns, and even slight differences in annual
precipitation can alter species assemblages [61]. Fall weather
patterns, especially interactions of precipitation and temperature after
rainfall, appear to be overriding factors in soft brome establishment
[45,46,57].
Fire affects plant species composition in annual grasslands largely by
removing mulch, which affects germination and seedling establishment
rates of soft brome relative to associated herbaceous species.
Bartolome [7,9] found that soft brome reached highest densities when
mulch biomass was at intermediate levels. Little quaking grass (Briza
minor) was favored when mulch biomass was low, as it would be in the
immediate postfire environment. Fescues (Vulpia and Festuca spp.) were
favored when mulch biomass was high. Heady [45] reported that without
heavy grazing the mulch layer usually recovers by postfire year 3, and
soft brome and other annual bromes regain dominance.
Decreases with fire: Hansen [40] found that fall prescribed fire in
Tulare County, California, significantly increased dominance of annual
forbs relative to soft brome. Greatest reduction soft brome and other
annual grasses (and greatest increase of annual forbs) was achieved by 3
years of successive fall burning. Response of native grasses was
similar to that of soft brome: Native grasses were reduced by fall
burning, with greatest reduction achieved after 3 years of consecutive
fall burning. Percent cover of soft brome the spring after fall burning
follows.
unburned single twice- thrice-
control burn burned burned
________ ______ ______ _______
1982 10 <1 -- --
1983 8 5 2 --
1984 23 44 16 2
1985 12 23 15 10
A July 1947 prescription fire reduced soft brome on ungrazed annual
grassland near Berkeley, California. Precipitation in the fall and
winter of 1947-1948 was slightly below average for the area (20.4 inches
with the average being 22.6 inches). Average height and yield of soft
brome on two burned and two unburned sites in May of 1948 was as follows
[48]:
burned unburned
______ ________
height (cm)
exclosure I 29.9 29.9
exclosure II 35.0 39.1
yield (g)
exclosure I 0.8 3.1
exclosure II 4.6 13.9
Mixed effects: Chaparral and oak woodland - Density of soft brome
increased greatly from prefire levels 5 years after prescribed fall
burning in a nonsprouting manzanita-Lemmon ceanothus (Arctostaphylos
spp.-Ceanothus lemmonii) community in Mendocino County. However,
density of soft brome had changed little 5 years after prescribed fall
fires in nearby nonsprouting manzanita (Arctostaphylos spp.)-Lemmon
ceanothus and interior live oak-blue oak (Quercus wislizenii-Q.
douglasii) woodland communities. Average density (plants/milacre) of
soft brome was [80]:
Postfire year
______________________________
Community Prefire 1 2 3 4 5
_____________________________________________________________
nonsprouting
manzanita-ceanothus 0.0 2.8 7.3 11.2 24.6 30.3
sprouting
manzanita-ceanothus 0.3 4.1 6.5 3.8 5.1 2.8
live oak-blue oak 1.5 6.6 6.7 5.8 3.0 1.3
No effect: Neither spring nor fall prescribed fire had significant
effect on soft brome in annual grassland of Sequoia National Park,
California. Precipitation averaged about 200 percent of normal during
postfire years 1 to 4. Soft brome formed an important component of the
vegetation (between 10 and 27%) on plots measured before fire and on
spring-burned, fall-burned, and unburned plots measured 4 years after
fire [75].
Sagebrush steppe - In central Idaho, fire had little effect on soft
brome coverage in either the long term or the short term. A long-term
study was conducted above the Snake River Canyon, after a July wildfire
occurred 1961 in a rubber rabbitbrush (Chrysothamnus
nauseosus)-cheatgrass community. At postfire year 12, soft brome had
declined on both burned and adjacent unburned plots. (Weather data were
not given.) Soft brome coverage was as follows [24]:
Unburned Burned
____________________ ___________________
Postfire year 2 4 12 2 4 12
____ ____ _____ _____ _____ _____
4.80 1.45 trace trace trace trace
A short-term study was conducted nearby when an August 1972 wildfire
occurred in a rubber rabbitbrush-cheatgrass stand within the Snake River
Canyon. The following spring, soft brome frequency was 21 percent on
unburned plots and 18 percent on burned plots [24].
DISCUSSION AND QUALIFICATION OF PLANT RESPONSE:
Fall and spring prescribed burning in east-central Oregon had no significant
effect on soft brome frequency in postfire year 1 or 2 [81]. See the
Research Project Summary of this study for more information on fire effects
on soft brome and 60 additional grasses, forbs, and woody plant species.
See the PDF of Hansen's [40] thesis, The effect of fire and fire frequency
on grassland species composition in California's Tulare Basin, for information
on the response of soft brome and other herbs to prescribed fire in an
annual grassland community.
FIRE MANAGEMENT CONSIDERATIONS:
California: annual grassland - Use of prescribed fire to increase the
balance of natives relative to non-natives such as soft brome has had
mixed results. In all cases, "remnant" California prairie contains
exotic annuals, and attempts to eliminate the exotics have been
unsuccessful [61]. However, fire sometimes tips the balance toward
natives. Perennial bunchgrasses are well adapted to frequent fire
[20,94]. Some authors have reported that fire favors native
bunchgrasses over exotic annuals [1,70]. However, Garcia and Lathrop
[33] reported no increase in purple needlegrass after burning, and
Lathrop and Martin [66] found that native deer grass (Muhlenbergia
rigens) decreased under some burning regimes. In view of the
differences in phenology and life histories between perennial
bunchgrasses and annual grasses such as soft brome, it would be
instructive to know how burning in different seasons affects the ratio
of native to non-natives. Since annual grasses produce seed about a
month earlier than perennial grasses, precise timing of burning may
alter the balance of reproductive success between annual and perennial
grasses [61].
When used with prescribed grazing, fire may favor purple needlegrass and
reduce soft brome and other annual grasses. Langstrotti [65] found that
on the Jepson Prairie (a relict perennial grassland reserve in Solano
County, California), short-term, intensive grazing by domestic sheep in
early spring (late March or early April) combined with late summer
(early September) prescribed fire favored tillering and seedling
establishment of purple needlegrass over exotic annual grasses including
soft brome. Purple needlegrass had been declining on the reserve for a
number of years. Frequency of soft brome was significantly reduced
(p=0.05) by early spring grazing and late summer fire. The treatments
reduced soft brome cover to less than 2 percent. Early spring grazing
reduced average seed mass, and the number of soft brome seeds was
reduced by 76 percent (p=0.25). Late summer fire reduced soft brome
cover by 50 percent (p<0.001). Summer grazing and late summer fire also
reduced soft brome, but not as much. Data from the spring
grazing/late summer fire treatments follow.
grazed- ungrazed-
burnt unburned
_______ __________
soft brome frequency (%) 39.7 3.0
soft brome seeds/sq dm 198 1,343
soft brome seed mass (mg) 0.57 0.97
Effects of postfire seeding of ryegrass on soft brome: Seeding Italian
ryegrass (Lolium multiflorum) to reduce postfire erosion had little
effect on postfire growth of soft brome and other exotic bromes in
southern California chaparral. Coverage of annual bromes was similar on
unseeded plots and on plots seeded with Italian ryegrass [15].
Oregon: big sagebrush - Prescribed fire had little effect on soft brome
in a basin big sagebrush/bluebunch wheatgrass community in John Day
Fossil Beds National Monument, Oregon. Weather patterns occurring after
fire greatly influenced plant community composition, however. One study
area was prescribed burned on September 25, 1987; an adjacent study area
was prescribed burned on May 24, 1988. Prescription burning was
followed by 3 years of drought, which appeared to greatly reduce soft
brome cover. By the third postfire year, soft brome was absent from all
treatments including the unburned control. Density of other annual
grass species was also greatly reduced on all treatments including the
unburned control. Density of annual forbs increased on all plots, and
density of native perennial grasses did not change. Density of woody
shrub species was greatly reduced on burned plots but did not change on
control plots. Average density of soft brome (plants/sq m) on unburned
control, fall-burned, and spring-burned plots is given below. Numbers
in parenthesis are the standard errors of the mean; different letters
denote a significant difference between years (p<0.1) [82].
1987 1988 1989
_________ _______ ______
control 160a (87) 0b (0) 0b (0)
fall burn 82a (28) 10b (8) 0b (0)
spring burn -- 37a (16) 0b (0)
References for species: Bromus hordeaceus
1. Ahmed, Elgaily Osman. 1983. Fire ecology of Stipa pulchra in California annual grassland. Davis, CA: University of California, Davis. 64 p. Ph.D. dissertation. [28343]
2. Amme, David; Pitschel, Barbara M. 1990. Restoration and management of California's grassland habitats. In: Hughes, H. Glenn; Bonnicksen, Thomas M., eds. Restoration `89: the new management challenge: Proceedings, 1st annual meeting of the Society for Ecological Restoration; 1989 January 16-20; Oakland, CA. Madison, WI: The University of Wisconsin Arboretum, Society for Ecological Restoration: 532-542. [14721]
3. Armstrong, K. C. 1981. The evolution of Bromus inermis and related species of Bromus sect. Pnigma. Botanische Jahrbucher Syst. 102(1-4): 427-443. [2933]
4. Ashby, William C.; Hellmers, Henry. 1955. Temperature requirements for germination in relation to wild-land seeding. Journal of Range Management. 8: 80-83. [25198]
5. Baker, H. G. 1986. Patterns of plant invasion in North America. In: Mooney, Harold A.; Drake, James A., eds. Ecology of biological invasions of North America and Hawaii. Ecological Studies 58. New York: Springer-Verlag: 44-57. [17511]
6. Barry, W. James. 1972. The Central Valley prairie. Vol 1. Sacramento, CA: State of California, Department of Parks and Recreation. 82 p. [28344]
7. Bartolome, James W. 1979. Germination and seedling establishment in California annual grasslands. Journal of Ecology. 67: 272-281. [28345]
8. Bartolome, James W. 1981. Stipa pulchra, a survivor from the pristine prairie. Fremontia. 9(1): 3-6. [28346]
9. Bartolome, James W. 1987. California annual grassland and oak savannah. Rangelands. 9(3): 122-125. [2861]
10. Bartolome, James W. 1989. Local temporal and spatial structure. In: Huenneke, L. F.; Mooney, H., eds. Grassland structure and function: California annual grassland. Dordrecht, The Netherlands: Kluwer Academic Publishers: 73-80. [28348]
11. Bartolome, James W.; Klukkert, Steven E,; Barry, W. James. 1986. Opal phytoliths as evidence for displacement of native California grassland. Madrono. 33(3): 217-222. [28349]
12. Bartolome, James W.; Stroud, Michael C.; Heady, Harold F. 1980. Influence of natural mulch on forage production on differing California annual range sites. Journal of Range Management. 33(1): 4-8. [28347]
13. Bean, Lowell John; Lawton, Harry W. 1973. Some explanations for the rise of cultural complexity in native California with comments on proto-agricultural and agricultural. In: Lewis, Henry T., author; Bean, Lowell John, ed. Patterns of Indian burning in California: Ecology and ethnohistory. Ramona, CA: Ballena Press: v-x1vii. [28350]
14. Bernard, Stephen R.; Brown, Kenneth F. 1977. Distribution of mammals, reptiles, and amphibians by BLM physiographic regions and A.W. Kuchler's associations for the eleven western states. Tech. Note 301. Denver, CO: U.S. Department of the Interior, Bureau of Land Management. 169 p. [434]
15. Beyers, Jan L.; Conard, Susan G.; Wakeman, Carla D. 1994. Impacts of an introduced grass, seeded for erosion control, on postfire community composition & species diversity in s. California chaparral. In: Proceedings, 12th conference on fire and forest meteorology; 1993 October 26-28; Jekyll Island, GA. Bethesda, MD: Society of American Foresters: 594-601. [26330]
16. Biswell, H. H. 1956. Ecology of California grasslands. Journal of Forestry. 9: 19-24. [11182]
17. Biswell, H. H.; Graham, Charles A. 1956. Plant counts and seed production on California annual-type ranges. Journal of Range Management. 9: 116-118. [28352]
18. Bodurtha, Timothy S.; Peek, James P.; Lauer, Jerry L. 1989. Mule deer habitat use related to succession in a bunchgrass community. Journal of Wildlife Management. 53(2): 314-319. [6677]
19. Clark, Ronilee A.; Halvorson, William L.; Sawdo, Andell A.; Danielsen, Karen C. 1990. Plant communities of Santa Rosa Island, Channel Islands National Park. Tech. Rep. No. 42. Davis, CA: University of California at Davis, Institute of Ecology, Cooperative National Park Resources Studies Unit. 93 p. [18245]
20. Clements, Frederic E. 1934. The relict method in dynamic ecology. Journal of Ecology. 22: 39-68. [11632]
21. Cooper, W. S. 1922. The broad-sclerophyll vegetation of California. Publ. No. 319. Washington, DC: The Carnegie Institution of Washington. 145 p. [6716]
22. Cronquist, Arthur; Holmgren, Arthur H.; Holmgren, Noel H.; Reveal, James L. 1972. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Vol. 1. New York: Hafner Publishing Company, Inc. 270 p. [717]
23. DaSilva, Paul G.; Bartolome, James W. 1984. Interaction between a shrub, Baccharis pilularis subsp. consanguinea (Asteraceae), and an ann. grass, Bromus mollis (Poaceae), in coastal CA. Madrono. 31(2): 93-101. [3198]
24. Daubenmire, Rexford F. 1975. Plant succession on abandoned fields, and fire influences, in a steppe area in southeastern Washington. Northwest Science. 49(1): 36-48. [745]
25. Bartolome, James W. 1987. California annual grassland and oak savannah. Rangelands. 9(3): 122-125. [2861]
26. Dittberner, Phillip L.; Olson, Michael R. 1983. The plant information network (PIN) data base: Colorado, Montana, North Dakota, Utah, and Wyoming. FWS/OBS-83/86. Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service. 786 p. [806]
27. Ewing, Anne L.; Menke, John W. 1983. Reproductive potential of Bromus mollis and Avena barbata under drought conditions. Madrono. 30(3): 159-167. [3522]
28. Ewing, Anne L.; Menke, John W. 1983. Response of soft chess (Bromus mollis) and slender oat (Avena barbata) to simulated drought cycles. Journal of Range Management. 36(4): 415-418. [3797]
29. Eyre, F. H., ed. 1980. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters. 148 p. [905]
30. Finnerty, D. W.; Klingman, Dayton L. 1962. Life cycles and control studies of some weed bromegrasses. Weeds. 10: 40-47. [921]
31. Flood, R. G. 1986. Germination of soft brome (Bromus hordeaceus). Plant Protection Quarterly. 1(4): 144-147. [22589]
32. Fulbright, Timothy E.; Redente, Edward F.; Hargis, Norman E. 1982. Growing Colorado plants from seed: a state of the art: Volume II: Grasses and grasslike plants. FWS/OBS-82/29. Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service. 113 p. [3709]
33. Garcia, Doris; Lathrop, Earl D. 1984. Ecological studies on the vegetation of an upland grassland (Stipa pulchra) range site in Cuyamaca Rancho State Park, San Diego County, California. Crossosoma. Claremont, CA: Southern California Botanists, Rancho Santa Ana Botanic Garden. 10(7): 5-12. [28353]
34. Garrison, George A.; Bjugstad, Ardell J.; Duncan, Don A.; [and others]. 1977. Vegetation and environmental features of forest and range ecosystems. Agric. Handb. 475. Washington, DC: U.S. Department of Agriculture, Forest Service. 68 p. [998]
35. Gleason, Henry A.; Cronquist, Arthur. 1991. Manual of vascular plants of northeastern United States and adjacent Canada. 2nd ed. New York: New York Botanical Garden. 910 p. [20329]
36. Glenn-Lewin, David C.; Johnson, Louise A.; Jurik, Thomas W.; [and others]. 1990. Fire in central North American grasslands: vegetative reproduction, seed germination, and seedling establishment. In: Collins, Scott L.; Wallace, Linda L., eds. Fire in North American tallgrass prairies. Norman, OK: University of Oklahoma Press: 28-45. [14194]
37. Gogan, Peter J. P.; Barrett, Reginald H. 1995. Elk and deer diets in a coastal prairie-scrub mosaic, California. Journal of Range Management. 48(4): 327-335. [25705]
38. Great Plains Flora Association. 1986. Flora of the Great Plains. Lawrence, KS: University Press of Kansas. 1392 p. [1603]
39. Green, Douglas M.; Kauffman, J. Boone. 1995. Succession and livestock grazing in a northeastern Oregon riparian ecosystem. Journal of Range Management. 48(4): 307-313. [25925]
40. Hansen, Robert Bruce. 1986. The effect of fire and fire frequency on grassland species composition in California's Tulare Basin. Fresno, CA: California State University, Fresno. 133 p. Thesis. [27963]
41. Halvorson, William L.; Clark, Ronilee A. 1989. Vegetation and floristics of Pinnacles National Monument. Tech. Rep. No. 34. Davis, CA: University of California at Davis, Institute of Ecology, Cooperative National Park Resources Study Unit. 113 p. [11883]
42. Harrington, H. D. 1964. Manual of the plants of Colorado. 2d ed. Chicago: The Swallow Press Inc. 666 p. [6851]
43. Heady, Harold F. 1961. Continuous vs. specialized grazing systems: a review and application to the California annual type. Journal of Range Management. 14: 182-193. [28354]
44. Heady, Harold F. 1973. Burning and the grasslands in California. In: Komarek, Edwin V., Sr., technical coordinator. Proceedings, annual Tall timbers fire ecology conference; 1972 June 8-9; Lubbock, TX. Number 12. Tallahassee, FL: Tall Timbers Research Station: 97-107. [8463]
45. Heady, Harold F. 1977. Valley grassland. In: Barbour, Michael G.; Major, Jack, eds. Terrestrial vegetation of California. New York: John Wiley and Sons: 491-514. [7215]
46. Heady, H. F.; Bartolome, J. W.; Pitt, M. D.; [and others]. 1992. California prairie. In: Coupland, R. T., ed. Natural grasslands: Introduction and western hemisphere. Ecosystems of the World 8A. Amsterdam, Netherlands: Elsevier Science Publishers B. V: 313-335. [23831]
47. Heady, Harold F.; Foin, Theodore C.; Hektner, Mary M.; [and others]. 1977. Coastal prairie and northern coastal scrub. In: Barbour, Michael G.; Major, Jack, eds. Terrestrial vegetation of California. New York: John Wiley and Sons: 733-760. [7211]
48. Hervey, Donald F. 1949. Reaction of a California annual-plant community to fire. Journal of Range Management. 2: 116-121. [1140]
49. Hickman, James C., ed. 1993. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 1400 p. [21992]
50. Hitchcock, C. Leo; Cronquist, Arthur. 1973. Flora of the Pacific Northwest. Seattle, WA: University of Washington Press. 730 p. [1168]
51. Hitchcock, C. Leo; Cronquist, Arthur; Ownbey, Marion. 1969. Vascular plants of the Pacific Northwest. Part 1: Vascular cryptograms, gymnosperms, and monocotyledons. Seattle, WA: University of Washington Press. 914 p. [1169]
52. Hoover, Robert Francis. 1935. Character and distribution of the primitive vegetation of the San Joaquin Valley. Berkeley, CA: University of California, Berkeley. 78 p. M.A. thesis. [28553]
53. Hull, A. C., Jr. 1973. Germination of range plant seeds after long periods of uncontrolled storage. Journal of Range Management. 26(3): 198-200. [18728]
54. Hulten, Eric. 1968. Flora of Alaska and neighboring territories. Stanford, CA: Stanford University Press. 1008 p. [13403]
55. Jain, S. K. 1982. Variation and adaptive role of seed dormancy in some annual grassland species. Botanical Gazette. 143 (1): 101-106. [28554]
56. Kartesz, John T. 1994. A synonymized checklist of the vascular flora of the United States, Canada, and Greenland. Volume II--thesaurus. 2nd ed. Portland, OR: Timber Press. 816 p. [23878]
57. Kay, Burgess L. 1987. Modifications of seedbeds with natural and artificial mulches. In: Frasier, Gary W.; Evans, Raymond A., eds. Proceedings of symposium: "Seed and seedbed ecology of rangeland plants"; 1987 April 21-23; Tucson, AZ. Washington, DC: U.S. Department of Agriculture, Agricultural Research Service: 221-224. [3851]
58. Kay, Burgess L.; Love, R. Morton; Slayback, Robert D. 1981. Discussion: revegetation with native grasses. I. A disappointing history. Fremontia. 9(3): 11-14. [28356]
59. Keeley, Jon E. 1981. Reproductive cycles and fire regimes. In: Mooney, H. A.; Bonnicksen, T. M.; Christensen, N. L.; [and others], technical coordinators. Fire regimes and ecosystem properties: Proceedings of the conference; 1978 December 11-15; Honolulu, HI. Gen. Tech. Rep. WO-26. Washington, DC: U.S. Department of Agriculture, Forest Service: 231-277. [4395]
60. Keeley, Jon E. 1990. The California valley grassland. In: Schoenherr, Allan A., ed. Endangered plant communities of southern California: Proceedings of the 15th annual symposium; 1989 October 28; Fullerton, CA. Special Publication No. 3. Claremont, CA: Southern California Botanists: 2-23. [21317]
61. Keeley, Jon E.; Keeley, Sterling C. 1984. Postfire recovery of California coastal sage scrub. The American Midland Naturalist. 111(1): 105-117. [5587]
62. Kuchler, A. W. 1964. United States [Potential natural vegetation of the conterminous United States]. Special Publication No. 36. New York: American Geographical Society. 1:3,168,000; colored. [3455]
63. Lambinion, J. 1981. Proposition de rejet des noms Aegilops ovata L., Bromus hordeaceus L., Carex muricata L., Crataegus X media Bechst., Crataegus oxyacantha L., Dipsacus fullonum L., Eurphorbia verrucosa L., Medicago polymorpha L., Phleum exartum Hochst. ex Grisb., Potamogeton pusillus L., Salix X smithiana Willd., Spergularia media (L.) C. Presl (=Arenaria media L.) et Vulpia membranacea (L.) Dum. (=Stipa membranacea (L.) Taxon. 30(1): 362. [28355]
64. Langstrotii, Robert Peter. 1991. Fire and grazing ecology of Stipa pulchra grassland: a field study at Jepson Prairie, California. Davis, CA: University of California. 75 p. Thesis. [27349]
65. Lathrop, Earl; Martin, Bradford. 1982. Fire ecology of deergrass (Muhlenbergia rigens) in Cuyamaca Rancho State Park, California. Crossosoma. Claremont, CA: Southern California Botantists, Rancho Santa Ana Botanic Garden. 8(5): 1-10; December. [28357]
66. Laude, Horton M. 1957. Growth of the annual grass plant in response to herbage removal. Journal of Range Management. 10(1): 37-39. [28358]
67. Martens, Ellen; Palmquist, Debra; Young, James A. 1994. Temperature profiles for germination of cheatgrass versus native perennial bunchgrasses. In: Monsen, Stephen B.; Kitchen, Stanley G., compilers. Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 238-243. [24289]
68. McBride, Joe; Heady, Harold F. 1968. Invasion of grassland by Baccaris pilularis DC. Journal of Range Management. 21: 106-108. [28359]
69. McClaran, Mitchel P. 1981. Propagating native perennial grasses. Fremontia. 9(1): 21-23. [28360]
70. McKell, Cyrus M.; Wilson, Alma M.; Kay, B. L. 1962. Effective burning of rangelands infested with medusahead. Weeds. 10(2): 125-131. [1617]
71. McNaughton, S. J. 1968. Structure and function in California grasslands. Ecology. 49: 962-972. [28361]
72. National Academy of Sciences. 1971. Atlas of nutritional data on United States and Canadian feeds. Washington, DC: National Academy of Sciences. 772 p. [1731]
73. Parker, Kenneth William. 1929. Growth of Stipa pulchra and Bromus hordeaceus as influenced by herbage removal. Berkeley, CA: University of California, Berkeley. 31 p. M.S. thesis. [28362]
74. Parsons, David J.; Stohlgren, Thomas J. 1989. Effects of varying fire regimes on annual grasslands in the southern Sierra Nevada of California. Madrono. 36(3): 154-168. [9244]
75. Radford, Albert E.; Ahles, Harry E.; Bell, C. Ritchie. 1968. Manual of the vascular flora of the Carolinas. Chapel Hill, NC: The University of North Carolina Press. 1183 p. [7606]
76. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press. 632 p. [2843]
77. Riegel, Gregg M.; Smith, Bradley G.; Franklin, Jerry F. 1992. Foothill oak woodlands of the interior valleys of southwestern Oregon. Northwest Science. 66(2): 66-76. [18470]
78. Robinson, Richard Hayes. 1971. An analysis of ecological factors limiting the distribution of a group of Stipa pulchra associations. Korean Journal of Botany. 14(3): 61-80. [28363]
79. Sampson, Arthur W. 1944. Plant succession on burned chaparral lands in northern California. Bull. 65. Berkeley, CA: University of California, College of Agriculture, Agricultural Experiment Station. 144 p. [2050]
80. Sampson, Arthur W.; Chase, Agnes; Hedrick, Donald W. 1951. California grasslands and range forage grasses. Bull. 724. Berkeley, CA: University of California College of Agriculture, California Agricultural Experiment Station. 125 p. [2052]
81. Sapsis, David B. 1990. Ecological effects of spring and fall prescribed burning on basin big sagebrush/Idaho fescue--bluebunch wheatgrass communities. Corvallis, OR: Oregon State University. 105 p. Thesis. [16579]
82. Seymour, Frank Conkling. 1982. The flora of New England. 2d ed. Phytologia Memoirs 5. Plainfield, NJ: Harold N. Moldenke and Alma L. Moldenke. 611 p. [7604]
83. Shiflet, Thomas N., ed. 1994. Rangeland cover types of the United States. Denver, CO: Society for Range Management. 152 p. [23362]
84. Smith, Winston Paul. 1985. Plant associations within the Interior Valleys of the Umpqua River Basin, Oregon. Journal of Range Management. 38(6): 526-530. [2179]
85. Stickney, Peter F. 1989. Seral origin of species originating in northern Rocky Mountain forests. Unpublished draft on file at: U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Fire Sciences Laboratory, Missoula, MT; RWU 4403 files. 10 p. [20090]
86. Tappeiner, John C., II; McDonald, Philip M.; Roy, Douglass F. 1990. Lithocarpus densiflorus (Hook. & Arn.) Rehd. tanoak. In: Burns, Russell M.; Honkala, Barbara H., technical coordinators. Silvics of North America. Volume 2. Hardwoods. Agric. Handb. 654. Washington, DC: U.S. Department of Agriculture, Forest Service: 417-425. [13969]
87. Thomas, Terri. 1990. Post-grazing grassland succession in Golden Gate National Recreation Area. In: Van Riper, Charles, III; Stohlgren, Thomas J.; Veirs, Stephen D., Jr.; Hillyer, Silvia Castillo, eds. Examples of resource inventory and monitoring in National Parks of California: Proceedings, 3rd biennial conference on research in California's National Parks; 1988 September 13-15; Davis, CA: Trans. and Proceedings Series No.8. Washington, DC: U.S. Department of the Interior, National Park Service: 195-202. [15200]
88. Thomas, Timothy W. 1987. Population structure of the valley oak in the Santa Monica Mountains National Recreation Area. In: Plumb, Timothy R.; Pillsbury, Norman H., technical coordinators. Proceedings of the symposium on multiple-use management of California's hardwood resources; 1986 November 12-14; San Luis Obispo, CA. Gen. Tech. Rep. PSW-100. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: 335-340. [5384]
89. U.S. Department of Agriculture, Natural Resources Conservation Service. 2018. PLANTS Database, [Online]. U.S. Department of Agriculture, Natural Resources Conservation Service (Producer). Available: https://plants.usda.gov/. [34262]
90. U.S. Department of the Interior, National Biological Survey. [n.d.]. NP Flora [Data base]. Davis, CA: U.S. Department of the Interior, National Biological Survey. [23119]
91. Voss, Edward G. 1972. Michigan flora. Part I. Gymnosperms and monocots. Bloomfield Hills, MI: Cranbrook Institute of Science; Ann Arbor, MI: University of Michigan Herbarium. 488 p. [11471]
92. Lockhart, Brian R.; Hodges, John D.; Guldin, James M. 1993. Development of advanced cherrybark oak reproduction following midstory & understory competition control & seedling clipping: 4-year results. In: Brissette, John C., ed. Proceedings, 7th biennial southern silvicultural research conference; 1992 November 17-19; Mobile, AL. Gen. Tech. Rep. SO-93. New Orleans, LA: U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station: 109-116. [23255]
93. Wells, Philip V. 1962. Vegetation in relation to geological substratum and fire in the San Luis Obispo Quadrangle, California. Ecological Monographs. 32(1): 79-103. [14183]
94. Welsh, Stanley L.; Atwood, N. Duane; Goodrich, Sherel; Higgins, Larry C., eds. 1987. A Utah flora. The Great Basin Naturalist Memoir No. 9. Provo, UT: Brigham Young University. 894 p. [2944]
95. Wester, Lyndon. 1981. Composition of native grasslands in the San Joaquin Valley, California. Madrono. 28(4): 231-241. [28364]
96. Willoughby, John W.; Davilla, William. 1984. Plant species composition and life form spectra of tidal streambanks and adjacent riparian woodlands along the lower Sacramento River. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 642-651. [5866]
97. Young, J. A.; Evans, R. A.; Raguse, C. A.; Larson, J. R. 1981. Germinable seeds and periodicity of germination in annual grasslands. Hilgardia. 49(2): 1-37. [4498]
98. Young, Richard P.; Miller, Richard. F. 1985. Response of Sitanion hystrix (Nutt.) J. G. to prescribed burning. The American Midland Naturalist. 113(1): 182-187. [28365]
FEIS Home Page